乌龙茶做青过程中导管形态和成分的变化研究
2020-10-21于松慧刘璇宋育林苏虹羊李静雯李小超陈玉琼倪德江余志
于松慧 ,刘璇 ,宋育林 ,苏虹羊 ,李静雯 ,李小超 ,陈玉琼 ,倪德江,余志 *
(1.华中农业大学园艺林学学院,园艺植物生物学教育部重点试验室,湖北武汉430070; 2.秭归县农业技术推广中心,湖北秭归 443600;3.华中农业大学农业部华中都市农业重点实验室,湖北武汉430070; 4.威泰克科学技术有限公司,北京100125)
植物导管是运输水分和内含物质的重要通道,因其次生壁加厚方式的不同,分为螺纹导管、环纹导管、梯纹导管、网纹导管以及孔纹导管[1]。不同植物中导管增厚方式有差异, 导管增厚结构会影响水分的传输。陈琦等[2]通过建立导管内壁增厚结构的伯努利数学模型发现导管内径、 增厚纹间距、纹高等对导管内水分传输有显著影响。导管微观结构决定功能[3],导管数量、直径等结构特征决定导管输导水分的能力。宋朝鹏等[4]对烘烤过程中烟叶的水分迁移特性进行研究发现叶片主脉直径的收缩与水分的散失密切相关。 导管直径的轻微改变就会引起导水效率显著变化[5-6]。 茶叶中的导管研究较少, 乌龙茶加工中导管本身的结构成分变化与茶叶品质之间的直接关系认识尚不充分。乌龙茶加工的关键环节做青包含摇青与晾青两道工序,二者交替进行[7]。做青经历“走水还阳”过程,促进茶叶中萜烯糖苷的水解, 增进香气的形成[8]。嫩茎中水分及内含物质通过导管运送至叶片中,被更充分地利用[9]。导管成分研究目前多采用将组织分离提取, 用傅立叶变换红外光谱或高效液相法进行检测[10]。 这些方法通常难以保证样品原生状态。 而激光共聚焦拉曼成像技术具有较高的空间分辨率,无需进行复杂的样品前处理,可以直接对样品进行无损、快速检测[11],将拉曼光谱数据转化为可视化图像, 还能显示出导管化学组成及成分分布变化信息。
研究以铁观音为材料, 用扫描电子显微镜和激光共聚焦拉曼成像技术直观展现茶叶中导管存在的主要类型,分析茎、叶柄、叶片主脉的导管数量差异, 以及上述三个部位的导管形态和结构成分在做青过程中的变化,丰富茶叶加工基础理论。
1 材料与方法
1.1 试验材料
2019 年8 月12 日, 选取华中农业大学茶资源圃铁观音茶树形成驻芽的一芽三叶为原料,按铁观音工艺进行加工,具体流程如下:鲜叶、萎凋、做青、炒青、揉捻、干燥。 做青3 次,第一次摇青1 min,晾青 1 h;第二次摇青 3 min,晾青 3 h;第三次摇青 6 min,晾青 6 h。 摇青机转速 25 r/min。 每一环节结束后分A、B 两组进行取样,每组5 个重复:A 组对鲜叶、萎凋叶、三次摇青叶和晾青叶的三个部位进行取样(图1),样品长度为3~5 mm,供扫描电镜测试;B 组对鲜叶、第三次晾青叶和炒青叶的部位3 进行取样, 样品长度大约为1 cm,供激光共聚焦拉曼成像测试。 取样部位如图1 所示,部位1 为第二叶与第三叶之间茎的中间部位;部位2 为第二叶叶柄与茎的连接部位; 部位3 为第二叶叶片中间主脉部位。

图1 铁观音鲜叶采摘标准及取样部位Fig. 1 Selection criteria and sampling sites of Tieguanyin fresh leaves
1.2 试验仪器
试验设备主要为:6CWL-85 型摇青机 (福建佳友茶叶机械有限公司)、6CST-50 型滚筒杀青机(浙江绿峰机械有限公司)、 生物扫描电子显微镜(JSM-6390LV) 和激光共聚焦拉曼成像系统(WITec,Alpha 300R)。
1.3 试验方法
1.3.1 扫描电镜观察导管形态
将A 组样品迅速转入2.5%成二醛溶液中固定,用梯度乙醇脱水,30%、50%、60%、70%和80%各8 min;90%和100%各6 min; 然后用乙酸异戊酯置换20 min,临界点干燥。分别将三个部位进行横切, 取3 个重复; 并将部位1 与部位3 进行纵切,取3 个重复。然后将样品横截面与纵切面用导电胶粘贴于样品台上, 用离子溅射仪将样品表面镀膜后,放入扫描电镜腔室进行观察。
1.3.2 激光共聚焦拉曼成像分析导管结构成分
将B 组样品迅速转入FAA 标准固定液(50%乙醇∶冰醋酸∶福尔马林=18∶1∶1)中固定,缓慢将叶片撕开,分离出导管。采用激光共聚焦拉曼成像系统进行光谱采集和成像。 光谱采集时采用100 倍物镜(ZEISS,NA=0.9)、50 倍物镜(ZEISS,NA=0.8)以及785 nm 激发波长。 测试时光栅为300 mm-1,扫描步距200 nm,成像时单点光谱采集时间3 s,扫描范围 200~1800 cm-1。
1.4 数据分析
应用 Nano Measurer 1.2 和 ImageJ 1.8.0 软件对导管相对数量和导管内径进行测量。 导管相对数量是在 X2500 (2000 μm2)下统计,随机统计 10个视野,取平均值;导管内径是随机测量10 个导管分子的长和宽,取平均值。 利用Excel 2010 软件对测量数据进行统计分析, 用SPSS 17.0 软件单因素方差分析中的Duncan 检验进行显著性分析 (p<0.05)。 采用 Origin 2018 软件作图。 利用 WITec Project 5 软件及自带数据库对激光共聚焦拉曼成像系统采集的拉曼光谱数据进行后期处理与分析。
2 结果与分析
2.1 茶叶中导管形态类型
茶叶导管在做青过程中结构发生变化。 图2表明, 鲜叶中茶叶导管存在形式主要有螺纹导管(图 2a)、网纹导管(图 2b)和孔纹导管(图 2c)3种。萎凋和前两次做青阶段孔纹导管较多,但在第三次摇青阶段后螺纹导管增多, 出现网纹与孔纹结合的网孔纹导管(图2d)且有导管壁消失的现象(图 2e 和图 2f)。
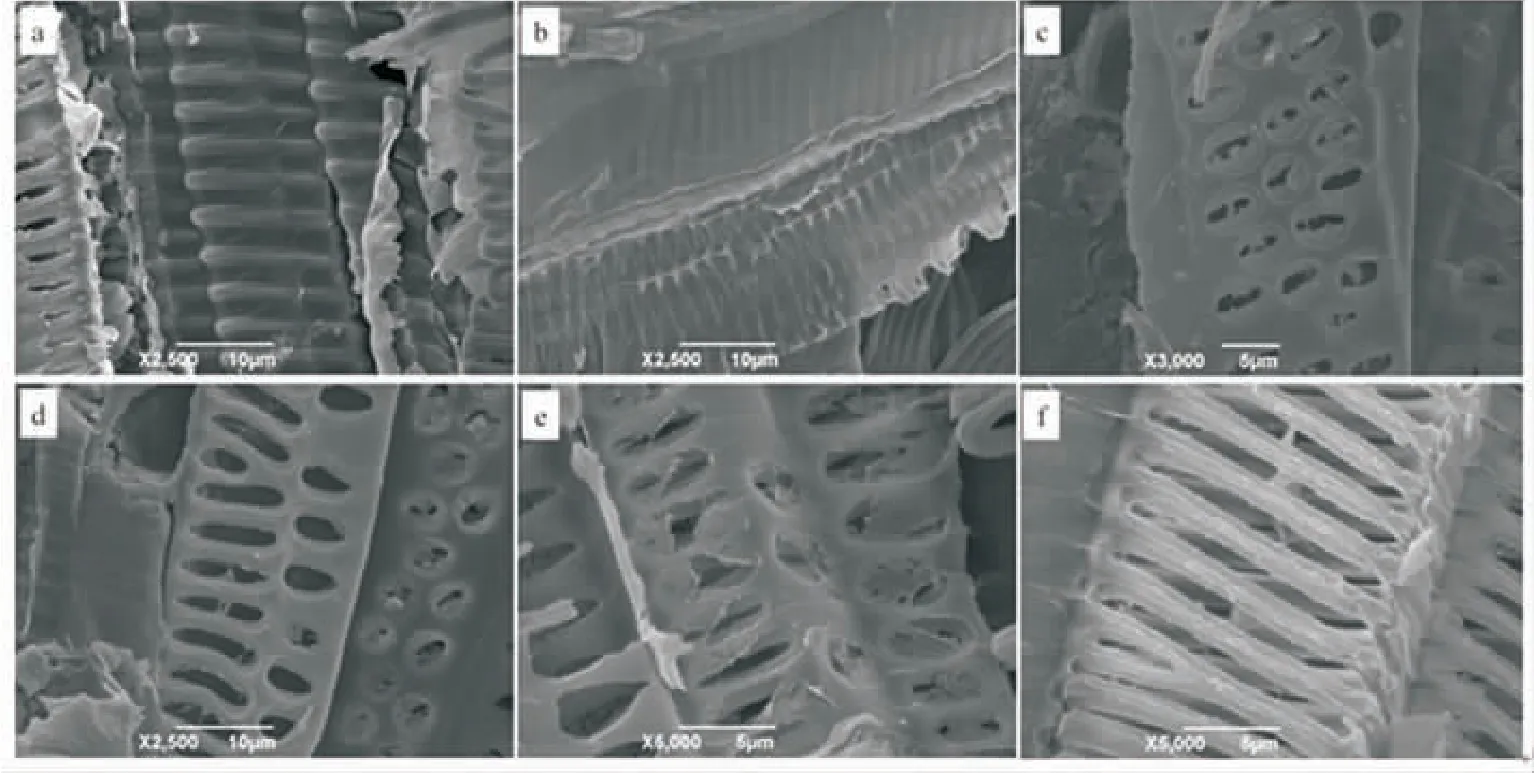
图2 茶叶中导管存在类型Fig. 2 Types of vessels in tea
2.2 不同部位导管相对数量
茶树地上部分的水分运输途径为茎木质部导管→叶柄木质部导管→叶脉木质部导管→叶细胞间隙→叶肉细胞→气孔→大气, 质外体途径是茶树水分主要运输途径[12]。 导管数量与导管运输能力密切相关,叶片、叶柄以及茎这三个部位是水分运输的必经之路, 其中叶柄的Ⅰ部分与叶片主脉相连, Ⅲ部分与茎相连, Ⅱ部分将Ⅰ与Ⅲ联系起来。对以上五个部分的导管相对数量进行比较,结果如表1 所示,导管相对数量大小顺序为:叶片主脉>Ⅰ>Ⅱ>Ⅲ>茎。
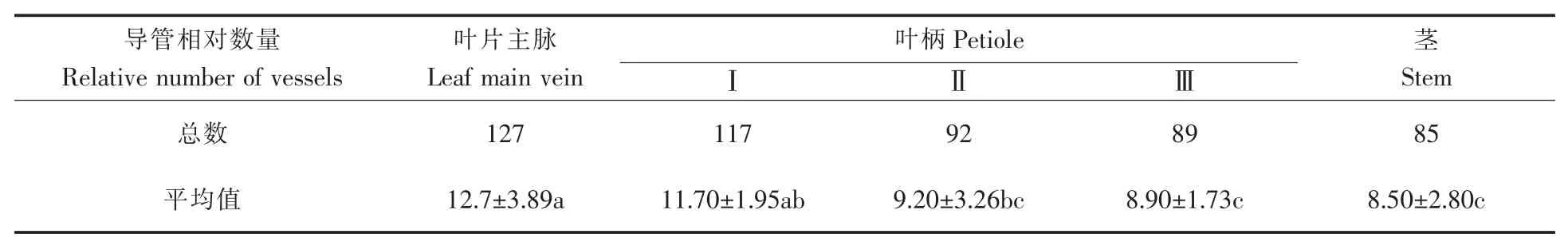
表1 不同部位导管相对数量Table 1 Relative number of vessels in different sites
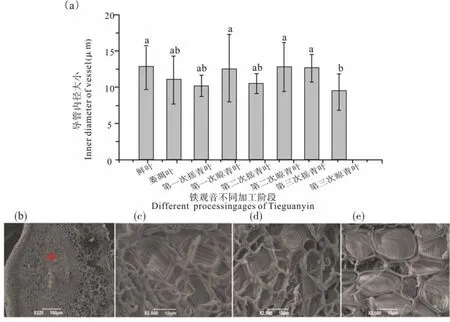
图3 铁观音在做青过程中叶片主脉导管内径及形态的变化Fig. 3 Changes of inner diameter and morphology of the vessels in leaves main vein of Tieguanyin during fine manipulation process
2.3 做青过程中叶片导管的变化特点
做青阶段叶片主脉导管内径有显著的变化(图3)。 萎凋阶段,叶片通过蒸腾作用失水,茎中水分向叶片主脉中迁移, 主脉中的水分向支脉木质部导管迁移,叶片整体失水速率大,导致导管内径呈缩小趋势。 摇青阶段,由于外力作用,茶叶发生碰撞, 导管皱缩, 叶片主脉导管内径进一步变小。晾青阶段,茶叶因为摇青受到机械损伤后叶片呼吸作用加剧,细胞膨胀产生张力,水分由茎中木质部导管向叶片主脉木质部导管迁移, 导致晾青后导管内径变大。在前两次做青中,导管内径都遵循摇青阶段减小,晾青阶段增大的趋势,但在第三次晾青叶中,叶片含水量已显著降低,导管内径不再增大,反而减小。
2.4 做青过程中茎部导管的变化特点
做青过程中茎内导管内径发生了显著变化(图4a)。 萎凋阶段,叶片蒸腾作用,茎内水分向叶片运输, 茎呈失水状态, 茎部导管内径呈缩小趋势。 第一次和第二次摇青阶段,茶叶经过碰撞,导管皱缩,茎部导管内径进一步减小(图4d)。 第一次和第二次晾青阶段, 在蒸腾拉力和毛细管力等作用下,茎内水分、组织间的自由水以及一些生化反应生成的内含物质通过导管壁上的开孔进入导管[13-14],茎部导管内径有增大趋势。 第三次摇青叶与第二次晾青叶相比, 茎部导管内径没有显著变化。但在第三次晾青结束,茎部导管周围的薄壁细胞皱缩,导管受到的压力减小,且导管自身形态和结构成分发生变化,出现网纹与孔纹结合的形式,导管壁消失, 导管排列松散, 茎部导管内径增大(图 4e)。
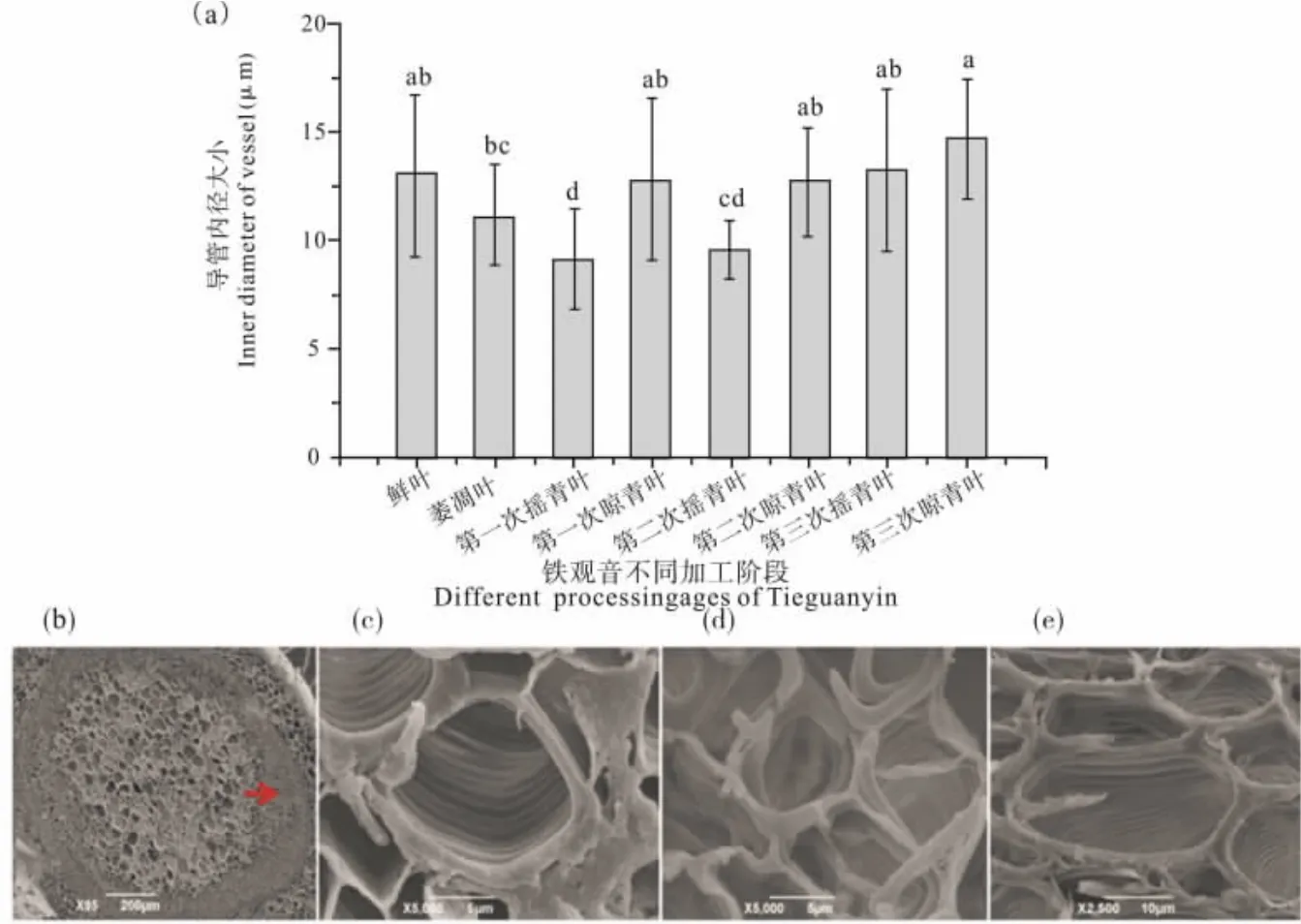
图4 铁观音在做青过程中茎部导管内径及形态的变化Fig. 4 Changes of inner diameter and morphology of vessels in stem of Tieguanyin during fine manipu lation process
2.5 做青过程中叶柄导管的变化特点
叶柄是连接茎与叶片的桥梁, 导管内径变化更复杂。 选择叶柄的三个关键部分Ⅰ、Ⅱ、Ⅲ观察发现不同部分导管内径大小有差异(图5b)。 其中Ⅰ与Ⅲ导管内径大于Ⅱ, Ⅱ多为小导管或正在发育的导管,内径小,数量少。在做青过程中,Ⅰ和Ⅱ导管内径变化趋势基本一致(图5a),即在萎凋和摇青阶段,导管内径呈减小趋势,在晾青阶段呈增大趋势,与茎中导管内径变化趋势相似。Ⅲ部分在做青过程中导管内径变化幅度较大, 与Ⅰ和Ⅱ在第一次摇青、 第三次摇青和第三次晾青阶段变化趋势不同。 第一次摇青叶的Ⅲ部分导管内径有增大的趋势,与叶片和茎的变化趋势都不同,原因有待研究; 第三次摇青叶和晾青叶的Ⅲ部分导管内径变化与叶片变化趋势相同,导管内径变小。
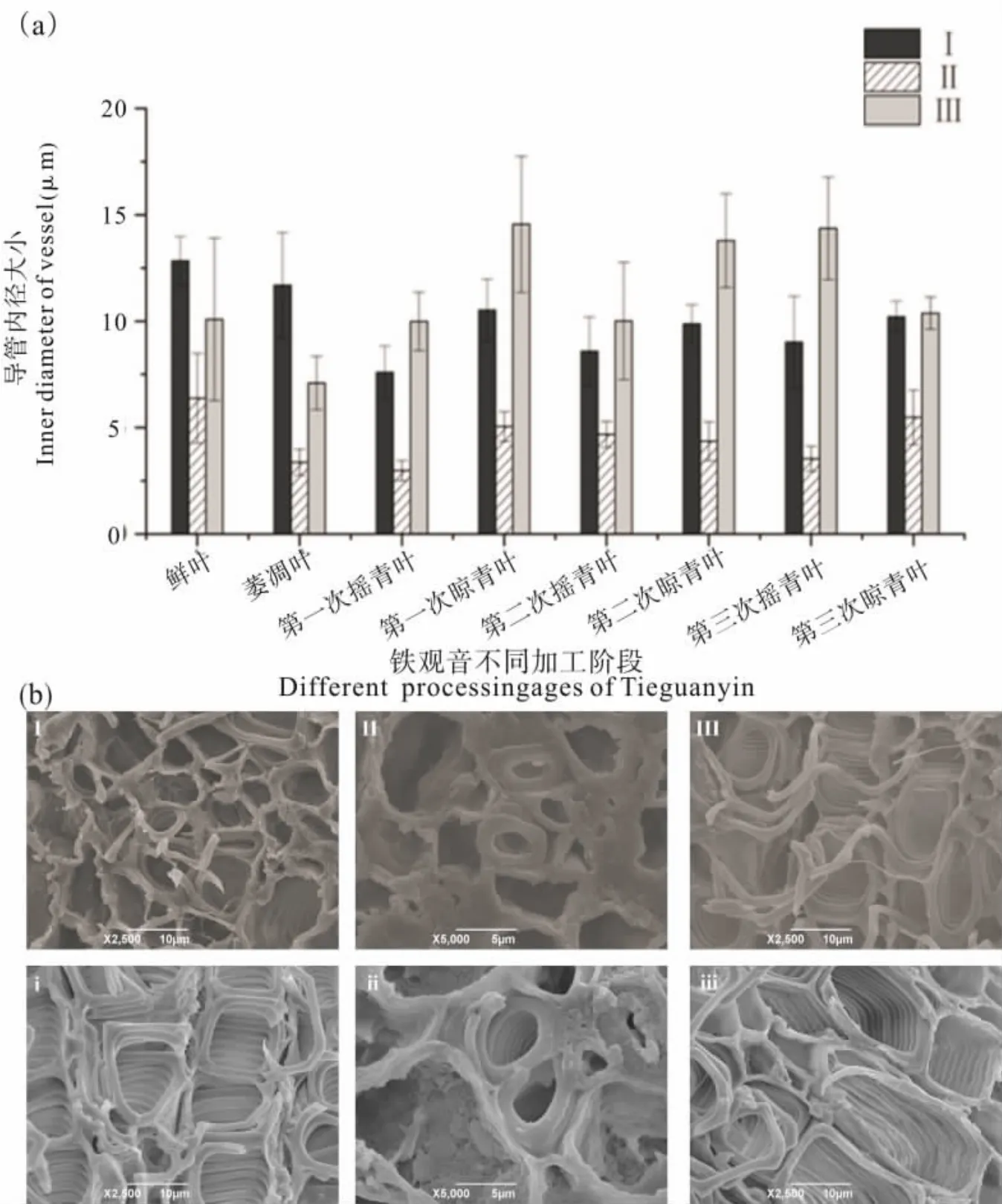
图5 铁观音品种在做青过程中叶柄导管内径及形态的变化Fig. 5 Changes of inner diameter and morphology of vessels in petiole of Tieguanyin during fine manipulation process
2.6 导管结构成分变化
将鲜叶、 第三次晾青叶和炒青叶叶片主脉导管进行分离(图6a),通过拉曼光谱分析鲜叶导管结构成分和做青、炒青过程对导管成分的影响(图6c)。 拉曼光谱属于分子特征光谱,谱带频率、强度和形状与测试分子结构密切相关,且可以反映出相应物质含量变化。各样品经激光共聚焦拉曼显微镜扫描并与数据库(ST-Japan)比对分析后发现:鲜叶导管中有三个是纤维素的特征峰[15-18],分别是378 cm-1的吡喃环C-C-C 的对称弯曲振动、1094 cm-1的糖苷键C-O-C 不对称伸缩振动和1121 cm-1的糖苷键 C-O-C 对称伸缩振动[19]。1279 cm-1是木质素中芳香环扭曲振动和C=O 伸缩振动[20]。 1329 cm-1由 HCC 和 HCO 弯曲振动引起[20],纤维素、半纤维素以及木质素对该峰都有贡献。 1370 cm-1峰由 HCC、HCO 和 HOC 弯曲振动引起[20],与纤维素和半纤维素有关。1458 cm-1与木聚糖中-OH 平面内弯曲振动和亚甲基变形振动有关[21]。 1602 cm-1和1659 cm-1是木质素的特征峰,1602 cm-1是苯环的对称伸缩振动,1659 cm-1是松伯醇/芥子醇结构的C=C 的伸缩振动[22]。 由以上分析结果推论,鲜叶中导管主要结构成分为纤维素、 半纤维素和木质素。 第三次晾青叶和炒青中导管结构成分种类变化不显著, 但是成分含量有变化。 做青后1279~1370 cm-1波段的峰,峰强明显减弱,该成分含量降低;炒青后,纤维素特征峰1092 cm-1强度显著降低。
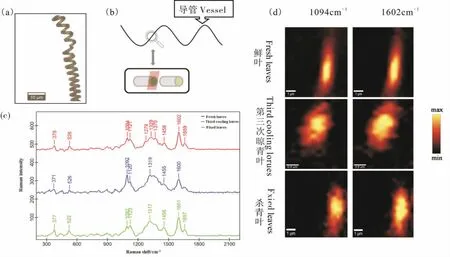
图6 叶片主脉导管在加工过程中拉曼光谱和共聚焦拉曼成像Fig. 6 Raman spectroscopy and confocal Raman imaging of the leaves main vein vessel during processing
为进一步分析导管主要结构成分纤维素(1094 cm-1)和木质素(1602 cm-1)在鲜叶,做青和炒青后其分布状态的变化, 用激光共聚焦拉曼成像技术对鲜叶、 第三次晾青叶和炒青叶中叶片主脉导管剖面扫描成像(图6b)。 拉曼映射强度与样品中所含有机成分的浓度成正相关。 由图6d 可见, 导管纵切剖面靠近中心部分纤维素和木质素含量高,越靠近导管表面,拉曼映射强度越低,纤维素和木质素分布较少。茶叶经过做青后,导管表面光斑分散,纤维素和木质素分布分散,显示导管结构可能出现松散状态, 这与扫描电镜观察到的网孔纹导管数量增加趋势符合。 高温炒青进一步影响导管中纤维素的分布, 导管表面拉曼映射强度与鲜叶相比显著降低。
3 讨论
茶叶中导管类型主要是螺纹导管、 网纹导管和孔纹导管。茶叶导管主要结构成分为纤维素、半纤维素和木质素,这与其他植物导管成分相似。随着茶叶加工的进行导管形态和结构成分会发生改变。 形态上, 导管在萎凋和摇青阶段导管内径减小,晾青阶段内径增大,但在第三次晾青结束,叶片含水率显著降低,主脉导管内经没有继续增大。茶叶从第三次摇青阶段后有网孔纹导管出现,而且有导管壁消失的现象。 产生这一现象的原因可能是摇青过程中叶片的碰撞[23]、晾青过程中叶片的翻动等多种外力作用对导管产生的拉力和做青过程中的酶促氧化对导管成分产生的影响。 为证实这一猜测, 研究采用激光共聚焦拉曼成像技术直观的观察做青和炒青中导管成分的变化。 扫描结果显示,在1270~1370 cm-1波段,峰强在做青后降低,有研究认为纤维素、半纤维素和木质素对这一波段波峰强度都有贡献[24]。 炒青后导管纤维素含量显著降低, 这可能与做青过程的酶促反应和高温杀青的热化学作用能使茶叶中部分大分子物质降解形成小分子物质[25],部分纤维素降解成可溶性糖有关。
研究结果清楚展示了导管在做青过程中形态与结构成分的变化规律, 也进一步印证乌龙茶选择茎壮叶肥的鲜叶原料, 木质部导管形态会更利于走水。在萎凋和晾青阶段,轻缓操作避免叶片损伤可以保持导管的结构, 利于实现水分和内含物质的迁移。同时随着加工进行,茶叶导管本身结构成分发生了变化,其中部分纤维素含量降低,水解成小分子碳水化合物参与茶叶品质的形成。
