人工脱涩处理对广西野生柿种质果实贮藏性的影响
2020-10-20黄思婕陆笛潘介春孙宁静
黄思婕 陆笛 潘介春 孙宁静
摘 要:為研究广西野生柿种质资源果实采后软化机制,该研究以广西特有的野生柿种质‘油柿(YS)、‘农家栽培柿(ZP)和主栽品种‘恭城月柿为材料,分析了人工脱涩乙烯利处理的果实采后贮藏中可滴定酸含量、可溶性糖含量、硬度、乙烯生物合成量、色差、可溶性单宁含量、果胶及细胞壁降解酶活性的变化。结果表明:(1)与‘恭城月柿相比,‘油柿可滴定酸含量高,可溶性糖含量低,转色缓慢,果实易软化。其中,YS-4最不耐贮藏,乙烯利处理后,贮藏第4天硬度降至1.620 N,第6天可溶性单宁含量下降至2.398 mg·g-1,整个贮藏期原果胶含量下降最慢,末期可溶性果胶值最低,为0.832%;YS-2最耐贮藏,乙烯利处理第8天硬度为YS-4的3.6倍,贮藏末期硬度在所有‘油柿中最高。(2)‘农家栽培柿中乙烯利处理贮藏末期ZP-2可滴定酸含量最高,可溶性糖含量低,转色最缓慢,原果胶降幅最小,可溶性果胶含量最高;相反ZP-3果实转色快且易软化,可滴定酸含量最低,可溶性糖含量低,原果胶降幅最大,可溶性果胶含量低。(3)‘恭城月柿PG酶活性和Cx酶活性远远高于‘油柿,各‘油柿种质的果实软化程度与细胞壁降解酶活性的相关性不同,β-D-Gal酶和Cx酶在‘农家栽培柿贮藏前期起作用,PG酶在贮藏前期和后期对果实软化起作用,说明不同柿种质品种在软化过程中起主导作用的酶不同。综上认为,与‘恭城月柿相比,‘油柿耐贮性差,其中YS-4极不耐贮藏,而‘农家栽培柿ZP-2极耐贮藏,这为研究柿果实软化机制提供了基础的种质材料。
关键词:柿种质, 脱涩处理, 贮藏, 软化, 酶活性
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2020)08-1188-15
Abstract:In order to investigate the softening mechanism of putharvest fruits of Diospyros germplasm, Diospyros oleifera, cultivated persimmon and the main cultivar ‘Gongcheng persimmon in Guangxi were used as experimental materials to analyze the changes of titratable acid content, soluble sugar content, firmness, ethylene biosynthesis, total chromatism, soluble tannin content, pectin content and cell wall degrading enzyme activities in postharvest storage of artificially dehydrated ethephon. The results were as follows:(1) Compared with ‘Gongcheng persimmon, the Diospyros oleifera had high titratable acid content, low soluble sugar content, slow color change and easy softening of fruits. Among them, YS-4 was the most resistant to storage. After ethephon treatment, the firmness decreased to 1.620 N on the 4th day, and the soluble tannin content decreased to 2.398 mg·g-1 on the 6th day. The pectin content decreased the most slowly during the whole storage period. The soluble pectin content value was the lowest at the final phase (0.832%); while YS-2 was the most resistant to storage, the firmness of ethephon treatment was 3.6 times than that of YS-4 on the 8th day, and the firmness at the end of storage was the highest among all persimmon germplasms. (2) The titratable acid content of ZP-2 was the highest, the soluble sugar content was low, the change of color was the slowest, the decline of original pectin was the least, and the content of soluble pectin was the highest after the end of storage period of ethephon treatment. On the contrary, ZP-3 fruit color changed quickly and softened easily, the titratable acid content was the lowest, the soluble sugar content was low, the original pectin content was the biggest, and the soluble pectin content was low. (3) Polygalacturonase (PG) and cellulase (Cx) of ‘Gongcheng persimmon were much higher than D. oleifera during the ripening period after fruit softening. The relationship between the softening degree and the cell wall degradation enzyme activity was different among all the persimmon germplasms. β-D-galactosidase (β-D-Gal) and cellulase (Cx) played an important role in the early stage of storage, while poly galacturonase (PG) played an important role in fruit softening at the early and late stage of storage. The results suggest that different persimmon germplasms have different enzymes that play major roles in fruit softening progress. In general, compared with ‘Gongcheng persimmon, the storage tolerance of D. oleifera, in which YS-4 was extremely intolerant to storage, and ZP-2 was extremely tolerant to storage. These results may provide a basis for the study of fruit softening mechanism of persimmon germplasm materials.
Key words:Diospyros germplasm, deastringency, storage, softening, enzymatic activity
柿属于柿科(Ebenaceae)柿属(Diospyros L.)落叶乔木,主要种植在热带和温带地区,中国作为柿的原产国,有柿树2属50多种,在世界上柿种植面积最大(高志强,2008)。广西作為中国柿主要产区,柿种质资源丰富,含有‘柿‘君迁子‘油柿‘乌柿‘毛柿和‘野柿等柿品种。在广西乡村农户房屋周围生长着多个野生‘油柿品种,叶片不具角质,叶面无光泽,具柔毛,果实接近成熟时表面附着有粘性物质,成熟时粘性物质不会消失,果面多有褐斑,通常种子大且较多,种植适应性广,多作为嫁接砧木(邓立宝等,2012)。农家栽培品种多生长于房前屋后,叶片革质,表面深绿光滑,背面淡绿有柔毛(邓立宝等,2012),果实中等大小,外观品质好,无种子或种子较少,由于无人管理,综合性状较差(邓立宝,2013)。Deng et al.(2015)对广西12个地区189份柿种质进行亲缘关系分析,发现乐业、西林、田林、右江地区以及泉州、环江、中山、鹿寨、武宣、秦塘和横县的柿种质亲缘关系相近;并运用Scot基因标记法将广西特有的‘油柿分为两个亚组,在欧氏距离D=12.09处,将35份第Ⅲ亚组‘油柿种质分为四个小亚组,第Ⅳ亚组‘农家栽培柿在欧氏距离D=9.59处,可将17份‘农家栽培柿种质分为三个小亚组(邓立宝,2013)。
柿属于呼吸跃变型果实,在贮藏过程中由内至外发生一系列转变(Smith et al., 2002),软化就是一个主要又直观的变化(Ahmed & Labavitch,1980)。果实表皮的软化是在各种大小物质共同作用下造成的,其中细胞壁结构与内在组分的改变对其影响最为直接(Huber,1983)。细胞壁由纤维素和果胶等物质构成,果胶作为细胞壁主要成分之一,随着贮藏时间的推移,果实逐渐成熟,果胶由原果胶向可溶性果胶转化,导致果实软化,硬度下降(Brummell, 2006)。在贮藏第5天自然成熟的‘恭城月柿果实软化度为第0天的3.5倍(范灵姣,2016);采后的‘安溪油柿在贮藏第33天时硬度降为6.0 N,贮藏期间会有呼吸高峰的出现,后期果实腐烂率高,果实品质大幅度降低(王慧等,2018)。软化是一个繁复的过程,除了大分子之间的转化,还有各种酶的协同作用,多聚半乳糖醛酸酶(PG)、纤维素酶(Cx)、果胶甲酯酶(PME)、β-半乳糖苷酶(β-D-Gal)和果胶酸移除酶(PL)等对果实软化具有调控作用,这些在柿(Kang et al., 1998;罗自生,2005)、鳄梨(Buse & Laties,1993)、苹果(Abwles & Biles,1991;申曙光,1991)、香蕉(Prabha & Bhagyalakshmi,1998)等均得到验证。
目前对广西野生柿种质资源果实软化相关研究尚未见报道。本研究将广西特有的‘油柿和‘农家栽培柿种质按照亲缘关系进行分组,研究不同‘油柿采后自然成熟和乙烯利脱涩处理二者果实软化相关指标之间的关系,并通过比较其与广西主栽品种‘恭城月柿耐贮性差异,探讨野生柿种质果实软化的原因,为研究柿果实软化机制提供种质基础,也为今后野生柿种质资源的开发利用提供理论依据。
1 材料与方法
1.1 材料
以‘油柿‘农家栽培柿和‘恭城月柿为材料,均采摘于广西大学农学院标本园柿种质资源圃。‘农家栽培柿与对应的‘恭城月柿采摘时间为2017年10月21日,贮藏至2017年11月6日;‘油柿与对应的‘恭城月柿采摘时间为2017年11月12日,贮藏至2017年11月26日。挑选大小均一、成熟度高度相似、无虫害和病害、无明显表面机械损伤的果实样品于恒温库中贮藏,贮藏温度为25 ℃(误差严格控制在1 ℃以下),相对湿度保持在60%~70%范围内浮动。
参照邓立宝(2013)的方法将‘油柿根据亲缘关系分为四个亚种,以‘YS-命名;农家栽培品种分为三个亚种,以‘ZP-命名。野生柿果实性状见表1。
1.2 试验处理
采用乙烯利脱涩处理果实,处理步骤如下:将柿果实完全浸泡于乙烯利溶液中,溶液浓度为500 mg·L-1,浸泡时长为5 min;5 min后,迅速捞出柿果实并置于阴凉处通风晾干,接着用密封袋密封24 h,之后放入恒温库贮藏;对照为相同处理方式,浸泡改用去离子水。间隔一天测定一次指标,测定后将果芯外围的果肉切碎,用液氮冷冻,置于-80 ℃的恒温库中。
1.3 测定指标与测定方法
1.3.1 硬度的测定 测定硬度主要参照Wang et al.(2018)的方法。处理组与对照组分别随机选取9个柿果实,分为3次重复,每次重复3个果实。测量时先将果皮削掉,穿刺时选用质构仪来测定硬度。
1.3.2 总色差的测定 总色差的测定,处理组与对照组随机选取9个柿果实,分为3次重复,每次重复3个果实。随机选取果实表面两对立面用色差计进行测定。
1.3.3 乙烯生物合成量的测定 参照刘思敏等(2018)的方法,处理组与对照组随机选取15个柿果实,分为3次重复,每次重复5个果实。将5个果实置于体积为4 L 的密封罐中封闭6 h后,抽取5管气体,之后于气相色谱仪测定。
1.3.4 可滴定酸的测定 可滴定酸采用氢氧化钠滴定法(曹建康等,2007)。
1.3.5 可溶性糖的测定 可溶性糖采用蒽酮试剂法(曹建康等,2007)。
1.3.6 可溶性单宁的测定 采用Folin-Denis法测定可溶性单宁(Taira,1996)。
1.3.7 原果胶与可溶性果胶的测定 采用咔唑比色法测定原果胶与可溶性果胶(曹建康等,2007)。
1.3.8酶液制备及酶活性测定 酶液的制备方法主要参照刘思敏等(2018)的方法,此外还进行了部分改进。具体制备步骤如下:
(1)取2.0 g果肉用液氮进行研磨,充分研磨后置于10 mL离心管中;(2)往管中加入4.0 mL浓度为0.2% 硫酸钠溶液,多次摇动离心管使研磨后的果肉和溶液混合均匀;(3)4 ℃下离心30 min,移除上清液,加入4.0 mL 浓度为100 mmol·L-1的醋酸钠缓冲液,再多次搖动使管中溶液混合均匀;(4)4 ℃下离心30 min,取出上清液弃去沉淀物,酶液制备完成。
酶活性的测定方法,果胶酶活性、纤维素酶活性和β-D-Gal酶活性测定分别参照刘思敏等(2018)的方法、3,5-二硝基水杨酸法(刘思敏等,2018)和Itamura(1986)的方法。
2 结果与分析
2.1 脱涩处理对野生柿种质贮藏期果实营养成分含量变化的影响
2.1.1 柿果实可溶性单宁含量变化 贮藏期间柿可溶性单宁呈下降趋势。由图1:A可知,第0天时‘油柿可溶性单宁含量均高于‘恭城月柿,贮藏过程中YS-2下降最慢,YS-5下降速度最快,在第10天时YS-5含量达到最低值,为0.440 mg·g-1。贮藏期结束后YS-2仍保持较高单宁含量,为21.722 mg·g-1,下降最少,其次为‘恭城月柿,可溶性单宁含量降幅为11.946 mg·g-1。由图1:B可知,乙烯利处理组‘油柿在0~10 d可溶性单宁含量大幅度下降,之后趋于平缓,第10天时可溶性单宁含量均低于5 mg·g-1,贮藏期结束时‘恭城月柿可溶性单宁含量最低,为0.002 mg·g-1,‘油柿含量为1 mg·g-1左右。
由图1:C可知,贮藏初期‘恭城月柿可溶性单宁含量最低,ZP-2最高。ZP-2在12~14 d可溶性单宁含量迅速下降;ZP-3在0~4 d可溶性单宁含量迅速下降;ZP-1贮藏后期比前期降幅要大;‘恭城月柿可溶性单宁含量在整个贮藏期下降缓慢。其中ZP-3可溶性单宁含量下降最多,为13.13 mg·g-1。
由图1:D可知,乙烯利处理后柿果实在贮藏12~16 d时可溶性单宁含量急剧下降。贮藏末期‘恭城月柿可溶性单宁含量最低,为6.75 mg·g-1,其次为ZP-3,为7.45 mg·g-1,ZP-2含量最高。其中ZP-3可溶性单宁含量下降最多,为17.99 mg·g-1。这说明野生柿种质单宁含量高于‘恭城月柿,乙烯利处理能加快可溶性单宁含量的下降。
2.1.2 柿果实可溶性糖含量变化 由图2:A可知,贮藏期间对照组中YS-5、YS-6可溶性糖含量呈上升趋势,YS-2、YS-4呈先上升后下降趋势,‘恭城月柿整体呈下降趋势。贮藏初期‘恭城月柿可溶性糖含量约为‘油柿的2倍。在12~14 d时,果实可溶性糖含量急剧下降至0.6%~0.8%。由图2:B可知,乙烯利处理组中果实在0~10 d可溶性糖含量呈下降趋势,之后开始上升。在8~10 d可溶性糖含量迅速下降低至1%以下,之后开始大幅度上升。如图2:C所示,在贮藏期间,‘恭城月柿可溶性糖含量呈上升趋势,而农家栽培品种可溶性糖含量总体呈下降趋势。贮藏期结束时恭城月柿可溶性糖含量为6.7%,农家栽培种可溶性糖含量为4.0%左右。如图2:D所示,乙烯利处理抑制月柿可溶性糖含量的上升,对‘农家栽培柿果实可溶性糖含量的影响不大。这说明‘油柿可溶性糖含量低于‘恭城月柿,‘油柿乙烯利处理后贮藏前期加速可溶性糖含量下降,末期促使可溶性糖含量上升;‘农家栽培柿经乙烯利处理可减缓柿果实可滴定酸含量的下降。
2.1.3 柿果实可滴定酸含量变化 由图3:A可知,果实贮藏期间可滴定酸含量整体呈下降趋势,初期‘恭城月柿可滴定酸含量最低,为1.10%,其次是YS-5,为1.16%,YS-6含量最高,为1.85%。贮藏期结束后YS-4可滴定酸含量下降最多,降幅为1.19%,YS-2可滴定酸含量下降,最小为0.34%,YS-5和‘恭城月柿分别下降0.66%和0.55%。由图3:B可知,乙烯利处理组YS-2可滴定酸含量下降最小,降幅为0.22%,YS-6降幅最大,为1.11%,月柿降幅为0.59%。由图3:C可知,除ZP-1外,柿果实贮藏期间可滴定酸含量整体呈下降趋势,贮藏初期ZP-2果实可滴定酸含量最高,为0.86%;ZP-3可滴定酸含量最低,为0.58%。贮藏第12 天时,ZP-1可滴定酸含量急剧上升。由图3:D可知,乙烯利处理能微弱减缓月柿可滴定酸含量的下降,在贮藏后期果实可滴定酸含量上升。这说明‘油柿可滴定酸含量高于‘恭城月柿,脱涩处理对‘油柿果实可滴定酸含量下降无太大影响,但是能减缓‘农家栽培柿果实可滴定酸含量的下降。
2.2 脱涩处理对野生柿种质果实软化和乙烯生成的影响
2.2.1柿果实硬度变化 在贮藏期间柿硬度呈下降趋势。由图4:A可知,不同‘油柿贮藏期存在差异。0~6 d时YS-4硬度迅速下降了27.08 N,之后下降趋于平缓;其次,YS-5在0~8 d迅速下降了25.40 N,之后趋于平缓;在8 d之后,‘恭城月柿的硬度均高于‘油柿,整个贮藏期间硬度下降幅度最小,为16.57 N。由图4:B可知,乙烯利处理组贮藏0~4 d硬度急剧下降,其中YS-4下降最为显著,下降了26.67 N;其次,YS-5下降了24.01 N,之后二者硬度下降趋于平缓;在第6天之后‘恭城月柿的硬度均高于‘油柿,直到贮藏末期‘恭城月柿硬度降至最低,为0.39 N。
由图4:C可知,‘恭城月柿起始硬度大于‘农家栽培柿。‘恭城月柿贮藏0~4 d和12~16 d降幅明显,贮藏中期降幅趋于平缓;ZP-1和ZP-2在第4天后硬度开始下降,在12~16 d降幅最大;ZP-3在贮藏0~12 d硬度下降缓慢,在第12天后硬度急剧下降至最低,为8.57 N。贮藏期间‘恭城月柿硬度降幅最大,为15.91 N;其次,ZP-3降幅为15.36 N;ZP-2降幅最小,为6.98N。由图4:D可知,乙烯利处理组在0~4 d硬度降幅大于对照组,ZP-3在8 d后开始急剧下降,ZP-1在整个贮藏期间降幅均匀,‘恭城月柿在8~12 d下降平缓,其他贮藏时期硬度急剧下降。贮藏末期‘恭城月柿硬度最小,为6.09 N,其次是ZP-3,为7.66 N,ZP-2硬度最大。这说明乙烯利处理能加快果实的软化,对‘油柿作用明显,而对‘农家栽培柿不明显。‘油柿中,YS-4软化最快,最不易软化;‘农家栽培柿中ZP-3最易软化,ZP-2最不易软化。
2.2.2 柿果实乙烯生物合成量变化 由图5:A可知,乙烯含量的变化呈上升-下降-上升-下降的趋势,峰值出现的时间存在差异。YS-2、YS-4和‘恭城月柿在第4天出现第一个高峰,分别为3.88、3.58和1.39 μL·g-1·h-1,YS-5、 YS-6峰值分别出现在第8天和第10天,分别为9.94和7.51 μL·g-1·h-1。由图5:B可知,乙烯利处理组YS-2、‘恭城月柿在第4天出现峰值,分别为9.76和3.44 μL·g-1·h-1,YS-4、YS-5在第8天出现峰值,分别为12.60、10.36 μL·g-1·h-1,YS-6在第10天出现峰值,为10.23 μL·g-1·h-1。由图5:C可知,乙烯含量的变化幅度不大,农家栽培品种在第12天出现峰值,而‘恭城月柿第4天和第12天出现两个峰值。由图5:D可知,经乙烯利处理后的不同柿乙烯生物合成量出现峰值的时间不同。ZP-1在第4天最先出现峰值,峰值为2.94 μL·g-1·h-1;接着第8天‘恭城月柿、ZP-2出现峰值,分别为2.34和2.00 μL·g-1·h-1;ZP-3出现峰值的时间最晚也最大,在第12天,峰值为2.56 μL·g-1·h-1。这说明‘油柿乙烯含量峰值远远大于‘农家栽培柿和‘恭城月柿,乙烯利处理对增大‘油柿乙烯生成量峰值效果极为明显,同样也加快ZP-1、ZP-2和‘恭城月柿峰值的出现。
2.2.3 柿果实总色差的变化 柿果实贮藏期间总色差呈上升趋势。由图6:A可知,总色差在0~4 d上升最为显著,与硬度在0~4 d急剧下降正好相反。贮藏期结束时‘恭城月柿色差值远远高于‘油柿,为25.28,其次是YS-4、YS-5,分别为14.77、14.44。由图6:B可知,乙烯利处理组总色差高于对照组,两组在8~10 d上升趋势均趋于平缓,第6天起‘恭城月柿总色差高于‘油柿。0~8 d‘恭城月柿总色差含量上升幅度大,之后趋于平缓,在12~14 d再次剧烈上升,而‘油柿在12~14 d上升幅度远远小于‘恭城月柿。由图6:C可知,总色差在0~4 d急剧上升,接着趋于平缓,之后又开始显著上升。其中ZP-3色差上升最快,直到贮藏末期ZP-3色差值最高,为18.6,其次到ZP-2,色差值为15.8,‘恭城月柿色差值最低,为13.0。由图6:D可知,乙烯利处理组总色差高于对照组,两组在8~12 d上升趋势均趋于平缓,贮藏前后期色差值呈显著上升趋势。至贮藏末期ZP-3色差值最高,为25.0,其次到ZP-2,色差值为20.0,ZP-1色差值最低,为15.2。这说明柿果实随着贮藏期延长,果实会逐渐变黄,乙烯利处理能加速果实的成熟与转色。
2.2.4 柿果实原果胶含量变化 在贮藏期间原果胶含量总体呈下降趋势。由图7:A可知,‘油柿起始值大于‘恭城月柿,且下降幅度高于‘恭城月柿,贮藏期结束后原果胶含量下降幅度YS-2最大为2.47%,其次是YS-4,为1.84%,‘恭城月柿最小,为0.14%。由图7:B可知,乙烯利处理组YS-2降幅为2.81%,YS-4降幅为2.54%,‘恭城月柿仅下降1.03%。说明‘油柿原果胶含量高于‘恭城月柿,在果实后熟时期原果胶含量降幅增大。由图7:C可知,农家栽培品种原果胶含量起始值大于‘恭城月柿。贮藏8天后原果胶含量呈显著下降趋势。贮藏末期ZP-3原果胶含量降幅最大,降幅為1.12%,ZP-2降幅最小,降幅为0.54%。由图7:D可知,经过乙烯利处理后柿果实在贮藏0~4 d内原果胶含量急剧下降。贮藏末期ZP-3原果胶含量降幅最大,下降1.37%,其次是‘恭城月柿,下降1.05%,ZP-2降幅最小,为0.79%。说明乙烯利处理可促进柿果实在贮藏初期原果胶含量的下降。
2.2.5 柿果实可溶性果胶含量的变化 果实贮藏期间可溶性果胶含量呈上升趋势。由8:A可知,可溶性果胶上升幅度,YS-2最大,为0.57%,其次是YS-4,为0.41%,‘恭城月柿最小,为0.09%。由图8:B可知,乙烯利处理组含量高于对照组。贮藏末期乙烯利处理组中YS-2可溶性果胶含量最高,为1.00%,上升幅度为0.68%,‘恭城月柿含量最低,为0.70%,升幅为0.34%。由图8:C可知,贮藏末期ZP-2可溶性果胶含量最高,为0.66%,其次是ZP-1,为0.57%,‘恭城月柿最小为0.44%。由图8:D可知,经乙烯利处理后0~4 d柿果实可溶性果胶含量迅速上升,处理大大提高了‘恭城月柿可溶性果胶含量的升幅,但对ZP-1和ZP-2影响不大。贮藏末期ZP-2可溶性果胶含量最高,为0.62%;ZP-3最低,为0.53%。这说明‘油柿乙烯利处理初期能促进可溶性果胶含量的上升,脱涩处理能加快原果胶向可溶性果胶的转化。
2.3 脱涩处理对野生柿种质果实细胞壁降解酶活性的影响
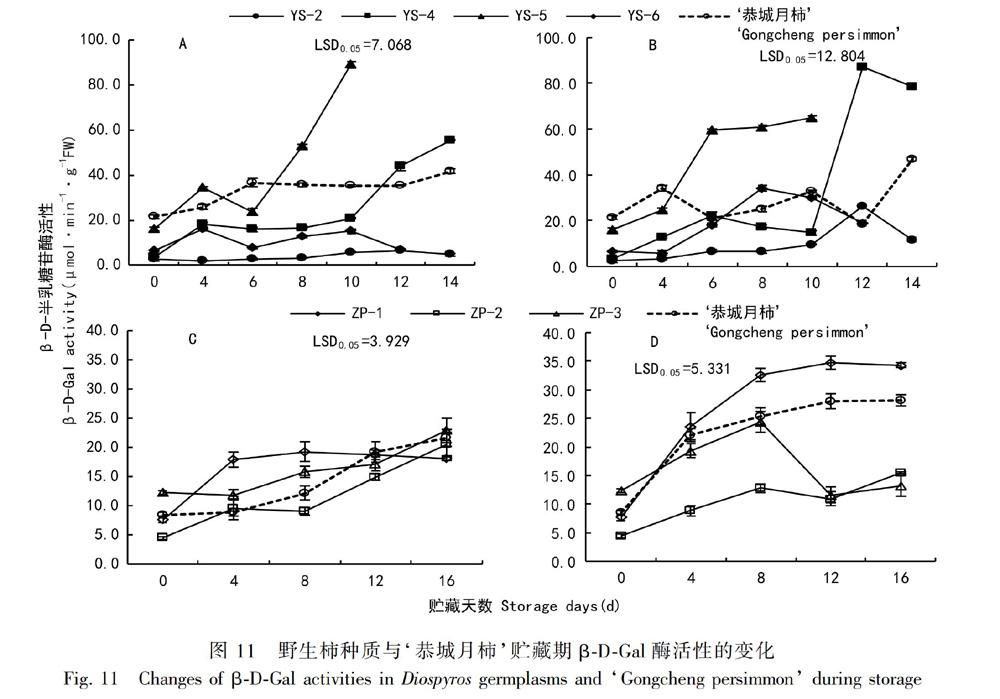
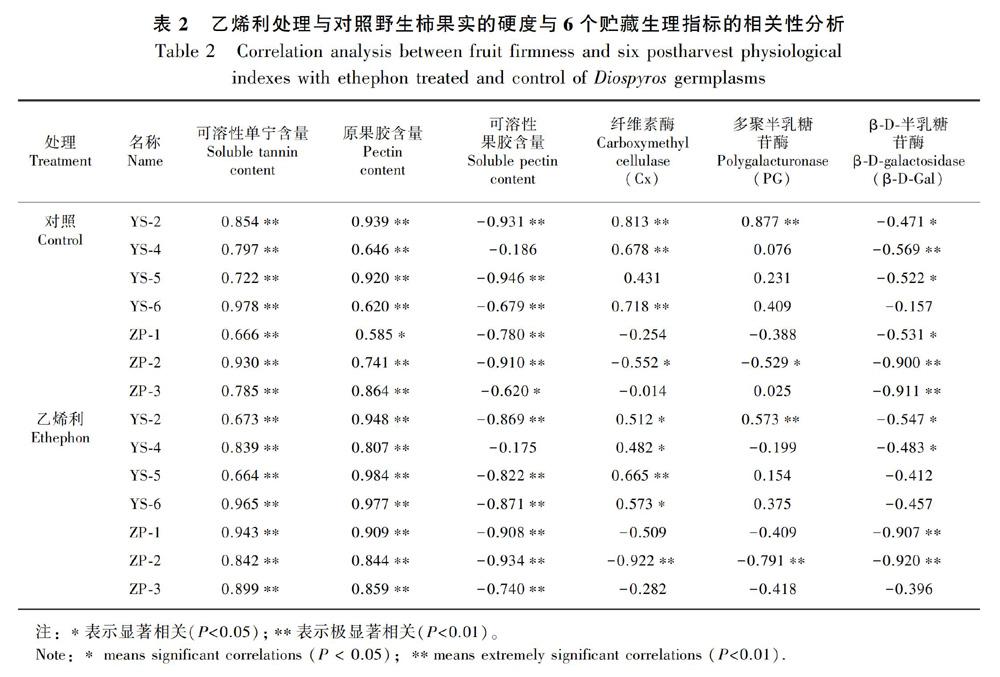
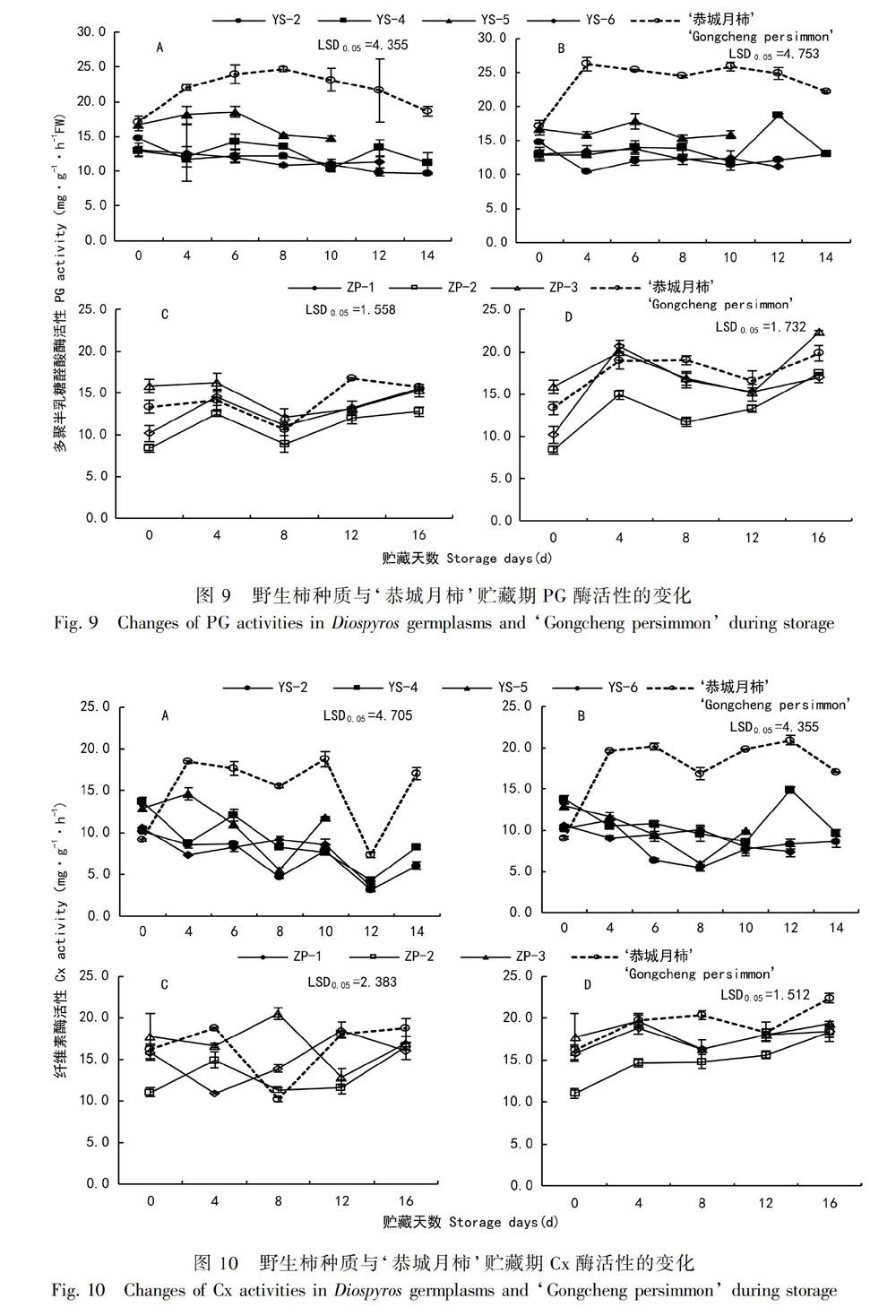
2.3.1 柿果实PG酶活性的变化 PG、Cx、β-D-Gal三个酶在果实贮藏软化过程中起着至关重要的作用。由图9:A可知,‘恭城月柿PG酶活性始终高于‘油柿,‘恭城月柿呈先上升后下降趋势,‘油柿整体呈下降趋势。‘恭城月柿在第8天酶活性达到最大值(24.67 mg·h-1·g-1)。‘油柿中YS-5酶活性最高,YS-4在第6天和第12天酶活性较高。由图9:B可知,处理组除YS-2外,均呈先上升后下降趋势,乙烯利处理组PG酶活性的降幅小于对照组。月柿在第4天酶活性达到最大值(26.22 mg·h-1·g-1),之后一直保持较高酶活性。YS-4在贮藏0~10 d期间酶活性波动小,末期酶活性出现一个高峰值。由图9:C可知,柿果实PG酶呈先上升后下降再上升趋势,贮藏第8天为酶活性最低点。ZP-1和ZP-2酶活性在第4天出现第一个峰值(分别为14.50、12.50 mg·h-1·g-1),之后酶活性下降又上升,直到第16天酶活性上升为最大值(15.60、12.81 mg·h-1·g-1),ZP-3的峰值出现在第4天(16.29 mg·h-1·g-1),‘恭城月柿的峰值出现在第12天(16.74 mg·h-1·g-1)。由图9:D可知,乙烯利处理酶活性趋势与对照组相同,处理推迟了ZP-1、ZP-3和‘恭城月柿酶活性最低点的出现,使酶活性峰值增高。‘恭城月柿PG酶活性始终高于‘油柿,软化后期仍保持较高的酶活性。YS-2硬度与PG酶活性呈显著负相关关系,ZP-2硬度与酶活性呈显著负相关关系(表2),说明PG酶对YS-2和ZP-2具有调控作用。
2.3.2 柿果实Cx酶活性的变化 由图10:A可知,‘恭城月柿Cx酶活性总体呈上升趋势,‘油柿总体呈下降趋势。在0~4 d,‘恭城月柿Cx酶活性迅速升高了9.33 mg·h-1·g-1;在10~12 d,Cx活性均下降至最低值,然后上升。贮藏末期YS-6酶活性下降最多,降幅为6.70 mg·h-1·g-1,其次是YS-4,降幅为5.50 mg·h-1·g-1。由图10:B可知,乙烯利处理能有效减小‘油柿的降幅,对‘恭城月柿基本无影响。‘恭城月柿酶活性在第4天迅速上升,之后一直保持较高的酶活性,YS-4在第12天出现酶活性高峰,峰值为14.92 mg·h-1·g-1。由图10:C可知,不同柿果实Cx酶活性变化趋势不同。ZP-1的峰值出现在第12天,ZP-2和‘恭城月柿酶活性峰值分别出现在第4天和第16天,ZP-3的酶活性峰值出现在第8天。由图10:D可知,乙烯利处理促进柿果实Cx酶活性的升高,对‘恭城月柿的促进作用最為显著,其次是ZP-3,对ZP-2酶活性升高的促进最不明显。
对照组YS-2、YS-4和YS-6的硬度与Cx酶活性呈显著性正相关,乙烯利处理组‘恭城月柿硬度与Cx酶活性呈显著性负相关,‘油柿与Cx酶活性呈极显著性正相关(表2),说明乙烯利处理能增强Cx酶对柿的调控作用。不同‘农家栽培柿Cx酶活性峰值出现时间不同,乙烯利处理能提高Cx酶活性峰值。乙烯利处理能有效抑制‘油柿Cx活性的下降。
2.3.3 柿果实β-D-Gal酶活性的变化 如图11:A所示,贮藏期间果实β-D-Gal酶活性整体呈上升趋势,YS-5和YS-4分别在第6天和第10天开始迅速上升,贮藏末期酶活性为89.03和55.50 mg·h-1·g-1。如图11:B所示,乙烯利处理组活性增幅大于对照组。不同‘油柿在经过乙烯利处理后酶活性迅速上升的时期不同,YS-5和YS-6在第4天后β-D-Gal酶活性迅速上升,YS-2和YS-4在第10天后活性迅速上升。对照组YS-4硬度与β-D-Gal酶活性呈显著负相关,对照组YS-2、YS-5和乙烯利处理组YS-2、YS-4的硬度与β-D-Gal酶活性呈极显著性负相关(表2)。如图11:C所示,贮藏期间果实β-D-Gal酶活性整体呈上升趋势。贮藏期结束时ZP-3的β-D-Gal酶活性最高,为22.95 mg·h-1·g-1,相反ZP-1最低,为18.06 mg·h-1·g-1。如图11:D所示,乙烯利处理促进ZP-1和‘恭城月柿在整个贮藏期间β-D-Gal酶活性的上升;而处理第8天后ZP-2和ZP-3酶活性受到抑制。说明乙烯利处理会抑制ZP-2和ZP-3贮藏后期β-D-Gal酶活性。ZP-1和ZP-2的硬度与β-D-Gal酶活性呈显著负相关关系(表2)。
3 讨论与结论
3.1 人工脱涩处理中柿果实软化与可溶性单宁含量变化的关系
单宁是果实涩味的来源,刚采收的涩柿果实中含有高浓度单宁,随着果实成熟,乙醛与可溶性单宁小分子结合成难溶性的缩合单宁大分子, 导致果实脱涩(殷学仁等,2011)。在贮藏后期,果实处于极度软化状态,可溶性单宁含量下降会加剧(张桂霞等,2009)。除此以外,缩合单宁与果胶发生相互作用,可以使果实涩味降低,果实脱涩(托尔坤·买买提,2017)。本研究中,可溶性单宁含量在贮藏期间随硬度的下降而下降,硬度与可溶性单宁含量呈极显著正相关关系。乙烯利处理后‘油柿硬度在0~4 d急剧下降,可溶性单宁含量在第4天后开始急剧下降;而‘农家栽培柿在12~16 d硬度和可溶性单宁下降速率增大,以上均验证了前人的研究结果。
3.2 柿果实软化与果胶含量变化的关系
果胶为组成细胞壁中胶层的主要物质,果实软化过程中先是中胶层的溶解,再是细胞壁的溶解,最终导致果实软化(王仁才等,2000)。在成熟软化前,果胶是以原果胶的形式存在,后熟期间中胶层的溶解是原果胶向可溶性果胶的转化,从而硬度下降;田建文等(1994)研究表明,果实的硬度与原果胶含量呈显著正相关关系,与可溶性果胶呈显著负相关关系。本研究中,‘油柿乙烯利处理组贮藏前期0~4 d原果胶含量大幅度下降,与0~4 d硬度急剧下降相对应,并且野生柿种质果实硬度与原果胶呈极显著正相关关系,与可溶性果胶呈极显著负相关关系,与前人研究结论一致。但Paull et al.(1996)研究木瓜发现,原果胶的迅速下降没有导致硬度的下降,YS-2可溶性果胶含量最高,原果胶下降最快,但硬度降幅最低,验证了原果胶的下降并不一定导致果实硬度的降低这一观点。
3.3 柿果实软化与细胞壁降解酶活性变化的关系
在果实软化过程中,细胞壁降解是引起果实软化的主要原因(范灵姣,2016)。然而,不同水解酶在不同果实中起到的作用不同,且在果实的不同发育阶段也有差异(Huber,1983)。魏建梅等(2009)、Ng et al.(2015)、刘思敏等(2018)研究认为β-D-Gal酶是导致果实软化的重要因素之一;而其他人研究发现,猕猴桃、青梅、番木瓜、鳄梨等PG酶活性随着果实软化程度的增加,酶活性会增强(Paull & Chen 1983;闫瑞香和王仁才,2000;陆胜民等,2000)。本研究中,Cx酶和PG酶活性在‘恭城月柿贮藏初期迅速升高之后一直保持较高的酶活性,抑制果实软化;ZP-1、YS-4和YS-5受Cx酶和β-D-Gal酶活性的调控,硬度的降低伴随着Cx酶活性的升高和β-D-Gal酶活性的降低,而YS-5正好相反;ZP-2、YS-2和YS-6果实软化受PG、Cx、β-D-Gal三种酶活性的调控,PG和Cx酶活性促进YS-2果实软化。不同柿种质果实硬度与相关酶活性相关性不同,不同柿果实软化过程中起主导地位的酶不同。
综上所述,作为广西野生柿种质资源,不同柿种质果实的贮藏性存在较大差异,影响果实软化的因素也不同。通过与广西主栽品种‘恭城月柿贮藏性的比较,证明以上特色野生柿可以作为研究果实软化机制的特色种质材料,同时也为提高柿果实贮藏性的育种提供种质基础。
参考文献:
ABWLES FB, BILES CL, 1991. Cellulase activity in developing apple fruit [J]. Sci Hortic, 47 (12):77-87.
AHMED A, LABAVITCH J, 1980. Cell wall metabolism in ripening fruit I. Cell wall changes in ripening ‘bartlett pears [J]. Plant Physiol, 65(5):1009-1013.
BRUMMELL DA, 2006. Cell wall disassembly in ripening fruit [J]. Plant Biol, 33:103-119.
BUSE E, LATIES G, 1993. Ethylene-mediated posttranscriptional regulation in ripening avocado (Persea americana) mesocarp discs [J]. Plant Physiol, 102(2): 417.
CAO JK, JIANG M, ZHAO YM, 2007. Study on physiology and biochemistry of fruits and vegetables after harvest [M]. Beijing:China Light Industry. [曹建康, 姜微波, 趙玉梅, 2007. 果蔬采后生理生化实验指导 [M]. 北京:中国轻工业出版社.]
DENG LB, 2013. Genetic diversity of persimmon germplasm resources and its resistance to corner spot disease in Guangxi [D]. Nanning:Guangxi University. [邓立宝, 2013. 广西柿种质资源遗传多样性及其对角斑病抗病性研究 [D]. 南宁:广西大学.]
DENG LB, HE XH, LI TW, et al., 2012. Investigation on germplasm resources and genetic diversity of persimmon in northwest plateau of Guangxi [J]. Acta Hortic Sin, 39(2):215-224. [邓立宝, 何新华, 李天文, 等, 2012. 广西西北部高原地区柿种质资源调查及遗传多样性分析 [J]. 园艺学报, 39(2):215-224.]
DENG LB, LIANG QZ, HE XH, 2015. Investigation and analysis of genetic diversity of diospyros germplasms using SCoT molecular markers in Guangxi [J]. PLoS ONE, 10(8):e0136510.
FAN LJ, 2016. Regulation of ascorbic acid on postharvest softening of persimmon fruit and its mechanism [D]. Nanning:Guangxi University. [范灵姣, 2016. 抗坏血酸对柿果实采后软化的调控作用及其研究机制 [D]. 南宁:广西大学.]
GAO ZQ, 2008. Study on persimmon processing technology of Anxi D. oleifera [D]. Fuzhou:Fujian Agriculture and Forestry University. [高志强, 2008. 安溪‘油柿柿饼加工工艺的研究 [D]. 福州:福建农林大学.]
HUBER DJ, 1983. The role of cell wall hydrolases in fruit softening [J]. Hortic Rev, 5:169-219.
ITAMURA H, 1986. Relationships between fruit softening, respiration and ethylene production after deastringent treatment by alcohol in Japanese persimmon (Diospyros kaki Thunb. var. ‘Hiratanenashi) fruits harvested at various stages [J]. Jpn Soc Hortic Sci, 55(1):89-98.
KANG IK, CHANG KH, BYUN JK, 1998. Changes in activities of cell wall hydrolases during ripening and softening in persimmon fruits [J]. J Korean Soc Hortic Sci, 39 (1):55-59.
LIU SM, HUANG SJ, LU D, et al., 2018. Relationship between postharvest redox potentials and ethylene synthesis and related enzyme activities in persimmon fruits [J]. Guihaia, 38(10):1326-1334. [刘思敏, 黄思婕, 陆笛, 等, 2018. 柿果实采后氧化还原电位与乙烯合成和相关酶活性的关系 [J]. 广西植物, 38(10):1326-1334.]
LU SM, XI QF, ZHANG YZ, 2000. Changes of softening and cell wall components and degradation enzyme activity of plum fruit after harvest [J]. Chin Agric Sci, 36(5):595-598. [陆胜民, 席琦芳, 张耀洲, 2000. 梅果采后软化与细胞壁组分及其降解酶活性的变化 [J]. 中国农业科学, 36(5):595-598.]
LUO ZS, 2005. Changes of cell wall component metabolism and ultrastructure in persimmon fruit during post-harvest softening [J]. Chin Physiol Mol Biol, 31(6):651-656. [罗自生, 2005. 柿果实采后软化过程中细胞壁组分代谢和超微结构的变化 [J]. 植物生理與分子生物学学报, 31(6):651-656.]
LUO ZR, 1996. Current status of germplasm resources and utilization of persimmon [J]. J Huazhong Agric Univ, 15 (4):381-382. [罗正荣, 1996. 柿属植物种质资源及其利用研究现状 [J]. 华中农业大学学报, 15(4):381-382.]
NG JK, SCHRODER R, BRUMMELL DA, et al., 2015. Lower cell wall pectin solubilisation and galactose loss during early fruit development in apple (Malus × domestica) cultivar ‘Scifresh are associated with slower softening rate [J]. J Plant Physiol, 176:129-137.
PAULL RE, CHEN NJ, 1983. Post harvest variation in cell wall degrading enzymes of papaya during fruit ripening [J]. Plant Physiol, 72:382-385.
PAULL KE, GROSS K, QIU YX, 1996. Changes in papaya cell walls during fruit ripening [J]. Postharvest Biol Technol, 7(4): 359-370.
PRABHA TN, BHAGYALAKSHMI N, 1998. Carbohydrate metabolism in ripening banana fruit [J]. Phyt Chem, 48(6):915-920.
SMITH D, ABBOTT J, GROSS K, 2002. Down-regulation of tomato β-galactosidase 4 results in decreased fruit softening [J]. Plant Physiol, 129(4):1755-1762.
SHEN SG, 1991. Physiological and biochemical changes of red Fuji apple during fruit development [J]. Acta Hortic Sin, 18 (1):1-3. [申曙光, 1991. 红富士苹果果实发育期间生理生化变化的研究 [J]. 园艺学报, 18 (1):1-3.]
TAIRA S, 1996. Fruit analysis:Astringency in persimmon [M]. Mod Meth Pant Anal, 18:97-110.
TIAN JW, HE PC, XU MX, 1994. Study on the relationship between physical and chemical indexes in post-ripening persimmon [J]. Acta Hortic Sin, 21 (1):41-46. [田建文, 贺普超, 许明宪, 1994. 火柿后熟中各理化指标间的关系研究 [J]. 园艺学报, 21(1):41-46.]
TORKUN MMT, 2017. Study on interaction between persimmon tannins and pectin [D]. Wuhan:Huazhong Agricultural University. [托尔坤·买买提, 2017. 柿单宁与果胶相互作用研究 [D]. 武汉:华中农业大学.]
WANG H, CHEN YH, LIN HT, et al., 2018. Effects of different concentrations of 1-mcp treatment on persimmon fruit preservation in Anxi after harvest [J]. Acta Trop Agric, 39(10):2060-2066. [王慧, 陈燕华, 林河通, 等, 2018. 不同浓度1-MCP处理对采后安溪‘油柿果实的保鲜效应 [J]. 热带作物学报, 39(10):2060-2066.]
WANG RC, XIONG XY, TAN XH, et al., 2000. Changes of postharvest hardness and cell wall ultrastructure of delicious kiwi fruit [J]. J Hunan Agric Univ (Nat Sci Ed), (6):457-460. [王仁才, 熊兴耀, 谭兴和, 等, 2000. 美味猕猴桃果实采后硬度与细胞壁超微结构变化 [J]. 湖南农业大学学报(自然科学版), (6):457-460.]
WEI JM, MA FW, GUAN JF, et al., 2009. Cell wall metabolism and its regulation during fruit ripening and softening of jingbai pear [J]. Chin Agric Sci, 42 (8):2987-2996. [魏建梅, 马锋旺, 关军锋, 等, 2000. 京白梨果实后熟软化过程中细胞壁代谢及其调控 [J]. 中国农业科学, 42(8):2987-2996.]
YAN RX, WANG RC, 2000. Physiological and biochemical mechanisms of fruit softening and senescence [J]. J Hunan Agric Univ, 26(3):230-234. [闫瑞香, 王仁才, 2000. 果實软化衰老的生理生化机制 [J]. 湖南农业大学学报, 26(3): 230-234.]
YIN XR, SHI YN, MIN T, et al., 2011. Expression of ethylene transduction elements in persimmon fruits [J]. Acta Hortic Sin, 38(Suppl.):2499. [殷学仁, 石艳娜, 闵婷, 等, 2011. 柿果实脱涩过程中乙烯转导元件的表达研究 [J]. 园艺学报, 38(增刊):2499.]
ZHANG GX, WANG YC, WEI X, et al., 2009. Changes of tannin and soluble solids in persimmon fruit during softening [J]. J Anhui Agric Sci, 37(14):6599-6600. [张桂霞, 王英超, 魏欣, 等, 2009. 柿果软化过程中单宁和可溶性固形物含量的变化 [J]. 安徽农业科学, 37(14):6599-6600.]
(责任编辑 何永艳)
