炼锌废渣-修复植物-凋落物体系的生态化学计量学研究
2020-10-20周佳佳罗有发刘明凤吴兴玉孙航吴永贵
周佳佳 罗有发 刘明凤 吴兴玉 孙航 吴永贵

摘 要:為揭示金属冶炼废渣堆场生态修复多年后,废渣-植物-凋落物系统中养分循环和系统维持机制。该研究以实现生态修复6 a的黔西北铅锌冶炼废渣堆场上土荆芥(Dysphania ambrosioides)、芦竹(Arundo donax)、刺槐(Robinia pseudoacacia)、构树(Broussonetia papyrifera)和柳杉(Cryptomeria fortunei)五种优势修复植物为对象,分析它们的主要营养器官(细根、粗根、茎/干、枝、叶片)、地表凋落物、植被下方表层废渣(0~10 cm)中碳(C)、氮(N)、磷(P)含量及化学计量特征,探讨它们之间的相互关系。结果表明:不同植物、不同营养器官间C、N、P的含量具有显著差异(P<0.05),C平均含量在两种草本植物中为茎>叶片>根>凋落物,在三种乔木中为干>枝>细根>粗根>叶片>凋落物;N和P的分布在草本植物中分别为叶片>凋落物>根>茎和叶片>根>凋落物>茎,在三种乔木中均为叶片>细根>凋落物>粗根>枝>干。五种植物中,柳杉各营养器官及凋落物中C含量均高于其他植物,N、P含量呈相反的规律;刺槐中N含量最高。C∶N和C∶P在五种植物营养器官与凋落物中的变化规律跟N、P的分布相反,说明C∶N和C∶P分别主要受N和P含量影响。相关性分析指出,草本植物的N∶P受N和P共同影响,三种乔木的N∶P主要由N的分布决定,同时受到枝和叶片中P含量影响。五种植物中,仅豆科类刺槐的叶片N∶P大于16,在系统中生长受P限制,其他植物生长均受N限制,说明刺槐更能适应贫瘠的废渣环境,建议在修复贫瘠的废渣堆场时优先选择豆科类植物作为先锋植物,改善基质养分条件。植被下方表层废渣中C、N、P含量基本都低于植物各营养器官及凋落物,不同修复植物下方对应的表层废渣中C、N、P含量间具有显著差异(P<0.05),草本植物修复下的废渣中C、N、P含量低于乔木修复下的含量。废渣-植物-凋落物体系中N、P、N∶P之间的相关性分析显示,植物细根和凋落物中N、P含量与废渣中N、P含量及化学计量比关系更密切。
收稿日期:2019-06-22
中图分类号:Q948
文献标识码:A
文章编号:1000-3142(2020)08-1079-13
Abstract:This study aimed to reveal the nutrient cycling and stability mechanisms in slag-plant-litter system on the metal smelting slags yard for many years of ecological restoration. Five dominant plants (Dysphania ambrosioides, Arundo donax, Robinia pseudoacacacia, Broussonetia papyrifera and Cryptomeria fortunei) planted on the lead-zinc smelting slag which had been ecologically restored for six years in Northwest Guizhou were selected as the objective in this study. The contents of carbon (C), nitrogen (N), phosphorus (P) and their stoichiometric characteristics in the organs of these dominant plants (fine root, coarse root, stem, branch and leaf), floor litter and surface waste slag beneath plants (0-10 cm) were studied, and their relationships were also determined. The results were as follows:There were significant differences in C, N and P contents between different plants and different vegetative organs(P<0.05). In the herbaceous plants,, the average content of C was stem > leaf > root > litter, and in the three trees, trunk > branch > fine root > coarse root > leaf > litter, The distributions of N and P in the herbaceous plants were leaf > litter > root > stem, leaf > root > litter > stem, while in the three trees, the distribution of N and P was leaf > fine root > litter > coarse root >branch > trunk. C content in litter of Cryptomeria fortunei was higher than that in all organs of others, but the contents of N and P showed an opposite rule. The highest concent of N was found in Robinia pseudoacacia. The change of C∶N and C∶P in organs and litter was contrary to the distributions of N and P, indicating that C∶N and C∶P were mainly affected by the content of N and P. Correlation analysis indicated that N∶P of herbaceous plants was affected by N and P. N∶P in the three arbor components was mainly controlled by N content, and also affected by P content in branches and leaves. Among the N∶P leaves of the five plant leaves, only the N∶P of leaves in Robinia pseudoacacia was greater than 16, and the growth in the system was restricted by P, while the growth of the other four plants were mainly restricted by N, indicating that Robinia pseudoacacia is more suitable for the waste residue environment, it was recommended to select legumes as pioneer plants in the restoration of barren waste residue yards to improve the substrate nutrient conditions. The contents of C, N and P in the waste slag were lower than those in organs and litter of the plant. The contents of C, N and P in the surface waste slag of different plants were significantly different (P <0.05). The contents of C, N and P in the residue under herbaceous restoration were lower than those under the three trees. Correlation analysis between N, P and N∶P between waste residue and plant-litter showed that the N and P contents in fine roots and litter of plants were more closely related to the N and P contents and stoichiometric ratio in the waste residue.
Key words:lead-zinc smelting slag, ecological restoration, plant nutrient organ, litter, C, N and P, ecological stoichiometry
碳(C)是构成植物生命体的骨架元素,植物通过光合作用吸收同化C作为其生理生化过程的重要能量来源,氮(N)、磷(P)是植物的基本营养物质及重要生长限制因子(Elser et al.,2007)。C、N、P等基本营养元素在植物生长及其各种生理调节机制中发挥重要作用(Fan et al.,2015)。生态化学计量学(ecological stoichiometry)结合了生态学和化学计量学的基本原理,是研究多种化学元素(C、N、P)质量平衡与生态系统能量平衡间相互影响的一种理论(王绍强和于贵瑞,2008)。植物C∶N和C∶P可反映植物生长速率快慢和养分利用效率高低(Wang et al.,2014),叶片N∶P可以判断植物生长受N或P限制的情况(Elser et al.,2007)。土壤作为植物生长所需营养的主要来源,其全氮、全磷含量可在一定程度上调节植物营养器官中C、N、P的化学计量比(Yu et al.,2011)。植物凋落物作为联系植物与土壤的重要纽带,其养分状况可一定程度上反映土壤的养分供应能力及植物对养分的利用状况。Parton et al.(2007)研究表明,在植物凋落物矿化分解过程中,当凋落物的C∶N<40时,凋落物分解释放净N,且凋落物的C∶N和C∶P与其分解速率间呈显著负相关。生态化学计量学研究在国内虽然起步较晚,但在近20 a取得大量研究成果,主要集中在自然情况下的森林生态系统、农田生态系统及湿地,且大部分是研究植物叶片的碳氮磷含量及化学计量特征,而对非自然生态系统中土壤-植物-凋落物体系的生态化学计量特征研究较少。
黔西北土法炼锌活动已有300余年历史,煤炭作为燃料和还原剂的土法炼锌工艺,将煤炭和铅锌矿原料混匀装罐,经过高温煅烧得到粗锌。由于冶炼工艺粗放,每生产1 t锌会产生3.0~3.8 t废渣(煤渣1.0 t,矿渣1.5~2.0 t,罐渣0.5~0.8 t)(王海峰等,2000),且废渣中含有大量重金属(Pb、Zn、Cu、Cd等),常年随意堆放废渣对周边及下游的生态环境带来严重安全隐患。生态修复重建是矿山废弃地环境治理的根本途径,关于废渣堆场生态修复系统中废渣生物化学、微生物学特性及废渣-植物系统中重金属的迁移转化已有报道(邢容容等,2018; Luo et al., 2018, 2019)。对于废渣-植物-凋落物体系中营养元素的生物地球化学循环的研究较少,对生境恶劣的金属冶炼废渣堆场上修复植物的养分供求与限制的认识存在不足。此外,以往的研究较少有涉及恶劣生境中废渣-植物-凋落物体系中养分的生态化学计量特征。因此,本研究以课题组前期在黔西北威宁县猴场镇开展生态修复6 a的某土法炼锌废渣堆场上的五种修复植物为对象,结合生态化学计量学方法,系统研究废渣-植物-凋落物体系中C、N、P含量及化学计量特征,探讨该体系中养分间的相互关系,为类似的金属冶炼废渣堆场生态修复、植物筛选及养分控制提供理论依据。
1 材料与方法
1.1 研究区概况
研究區位于贵州省黔西北威宁县猴场镇群发村某土法炼锌废渣堆场生态修复示范区(104°43′45″ E,26°41′14″ N),平均海拔2 200 m,年平均气温约10 ℃,亚热带季风气候带,年降水量约890 mm,无霜期180 d。课题组于2012年初在研究区通过工程措施进行废渣场地平整,并通过添加有机改良剂对废渣进行必要的基质改良,同时搭配乔木(构树、刺槐、柳杉)和草本(芦竹)构成多元植物群落,随着废渣植生条件不断改善,土荆芥在该系统中自然着生,截至2017年12月,各类植物已在该堆场稳定生长6 a。
1.2 研究方法
1.2.1 样品采集 2017年12月初,在研究区开展样品采集工作,主要采集桑科的落叶乔木构树(Broussonetia papyrifera)、豆科的落叶乔木刺槐(Robinia pseudoacacia)、杉科的常绿乔木柳杉(Cryptomeria fortunei)、禾本科的多年生草本植物芦竹(Arundo donax)和藜科黎属土荆芥(Dysphania ambrosioides)。根据植物的分布特征将研究区划分为3个40 m × 40 m的样方,在每个样方中分别选取长势相同的3株土荆芥、芦竹、刺槐、构树和柳杉,每个样方中选取3株长势相当的土荆芥和芦竹整株拔起,分别采集植物的根系、茎和叶,并收集植株周边的地表凋落物进行混匀;在刺槐、构树和柳杉的1~1.5 m胸径处采集树干,在树冠不同方位采集枝和叶,挖取的根按直径大于2 mm为粗根和小于2 mm为细根分别采集,收集样方中不同植物的凋落物,将采集的样品用四分法缩分至剩300 g左右。废渣样品的采集:采集每种植物下方对应的表层(0~10 cm)废渣样品约500 g。将每个样方的植物各组分(细根、粗根、茎/干、枝、叶和凋落物)以及对应的表层废渣带回实验室。
1.2.2 样品制备及分析 将采集的植物样品先用自来水洗去尘土、去离子水冲洗干净,晾干后于105 ℃杀青30 min,70 ℃下烘干至恒重,用植物破碎机破碎、过80目筛后测定植物样品中C、N、P含量。将采集的废渣样品去除其中的石子和动植物残体后,在自然条件下置于室内自然风干,研磨,分别过60、100目筛后测定废渣的C、N、P含量。
植物不同器官、凋落物及废渣中C、N、P含量采用常规方法测定;全碳用重铬酸钾外加热法测定;全氮用半微量凯氏定氮法测定;植物与凋落物的全磷用H2SO4-H2O2消解;废渣的全磷用H2SO4-HClO4消解,消解液均用钼锑抗比色法测定。所有指标的实验过程均采用平行样和空白样进行全程质量控制。
1.2.3 数据处理 所有数据整理采用Microsoft Office Excel 2007软件,其中C、N、P的化学计量比均为质量比,所有数据均以算术平均值±标准误差表示,同时采用SPSS20中单因素方差分析(one-way ANOVA)及Duncans 新复极差分析法对不同植物修复下的废渣、不同植物的相同营养器官、相同植物的不同营养器官中C、N、P含量及化学计量特征进行显著差异性检验(P<0.05),并用Pearson相关性分析废渣-植物-凋落物中N、P含量及化学计量比之间的相关关系。
2 结果与分析
2.1 不同修复植物各器官及凋落物中C、N、P分布特征
从表1可以看出,各组分中C、N、P含量在不同植物间均存在显著性差异(P<0.05)。土荆芥的茎、构树和柳杉的干中C含量均高于其他组分,凋落物中C含量均低于叶片,C在两种草本植物中平均含量为茎>叶>根>凋落物,在三种乔木中依次为干>枝>粗根>细根>叶>凋落物;刺槐各组分中N含量均显著高于其他四种植物(P< 0.05),柳杉的细根、粗根、干、叶和凋落物中N含量均最低,除构树外,其他四种植物茎/干中N含量均显著低于别的组分(P<0.05),土荆芥和芦竹中N平均含量为叶>凋落物>根>茎,刺槐、构树和柳杉中N平均含量为叶>细根>凋落物>粗根>枝>干;构树的细根、粗根和凋落物中P含量均最高,而柳杉的粗根、细根、干和凋落物中P含量显著低于其他植物,三种乔木的不同组分中P的分配规律与N相同,两种草本植物各组分中P含量依次为叶>根>凋落物>茎。
2.2 五种修复植物各器官及凋落物中C、N、P化学计量特征
从表2可以看出,不同植物不同组分间的C∶N、C∶P和N∶P具有显著差异(P<0.05)。柳杉各组分的C∶N显著高于其他植物(枝除外),刺槐各组分的C∶N最低。C∶N在土荆芥和芦竹各组分中平均大小为茎>根>凋落物>叶,在三种乔木中为干>枝>粗根>凋落物>细根>叶,均与N在植物中的分布呈相反规律,可见,修复植物各组分的C∶N高低主要由N的含量决定。两种草本植物各组分中C∶P为茎>凋落物>根>叶,三种乔木各组分中C∶P的大小顺序与C∶N相同,五种植物的C∶P均与P的分布呈相反规律,说明植物各组分中C∶P主要由P含量决定。土荆芥和和芦竹各组分中N∶P平均大小为叶>凋落物>茎>根,刺槐、构树和柳杉不同组分中均值大小为叶>枝>细根>干>凋落物>粗根,各植物不同器官间N∶P差异显著,刺槐各组分中N∶P显著高于其它四种植物(P< 0.05)。
2.3 渣场修复植物下方废渣中C、N、P含量及化学计量特征
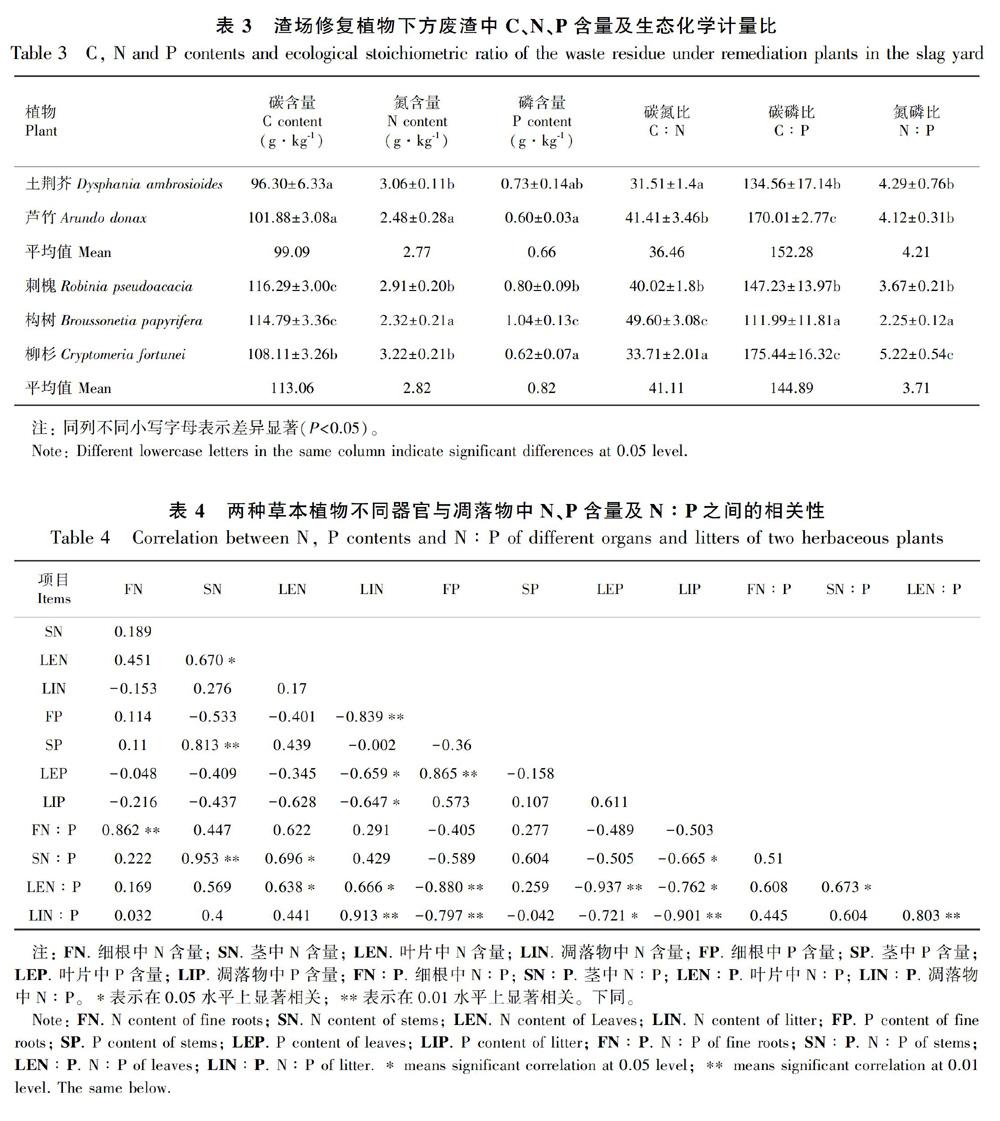
表3结果表明,修复植物下方的废渣中C、N和P含量均低于植物各营养器官和凋落物,不同植物生长下废渣中C、N、P含量差异显著(P<0.05)。废渣中C含量为96.30~116.29 g·kg-1,刺槐生长下的废渣中C含量最高;废渣中N含量范围为2.32~3.22 g·kg-1,五种植物对应的表层废渣中N含量依次为柳杉>土荆芥>刺槐>芦竹>构树;废渣中P含量为0.60~1.04 g·kg-1,构树生长下的废渣中P含量最高。
五种植物生长下废渣中C∶N为31.51~49.6,表现为构树>芦竹>刺槐>柳杉>土荆芥;C∶P的范围为111.99~175.44,柳杉和芦竹生长下,废渣中C∶P显著高于其他三种修复植物;废渣中N∶P的范围为2.25~5.22,构树和柳杉生长下废渣中N∶P分别最低和最高,不同修复植物生长下废渣的N∶P差异显著(P<0.05)。
2.4 废渣-植物营养器官-凋落物中N、P含量及化学计量特征间相关性分析
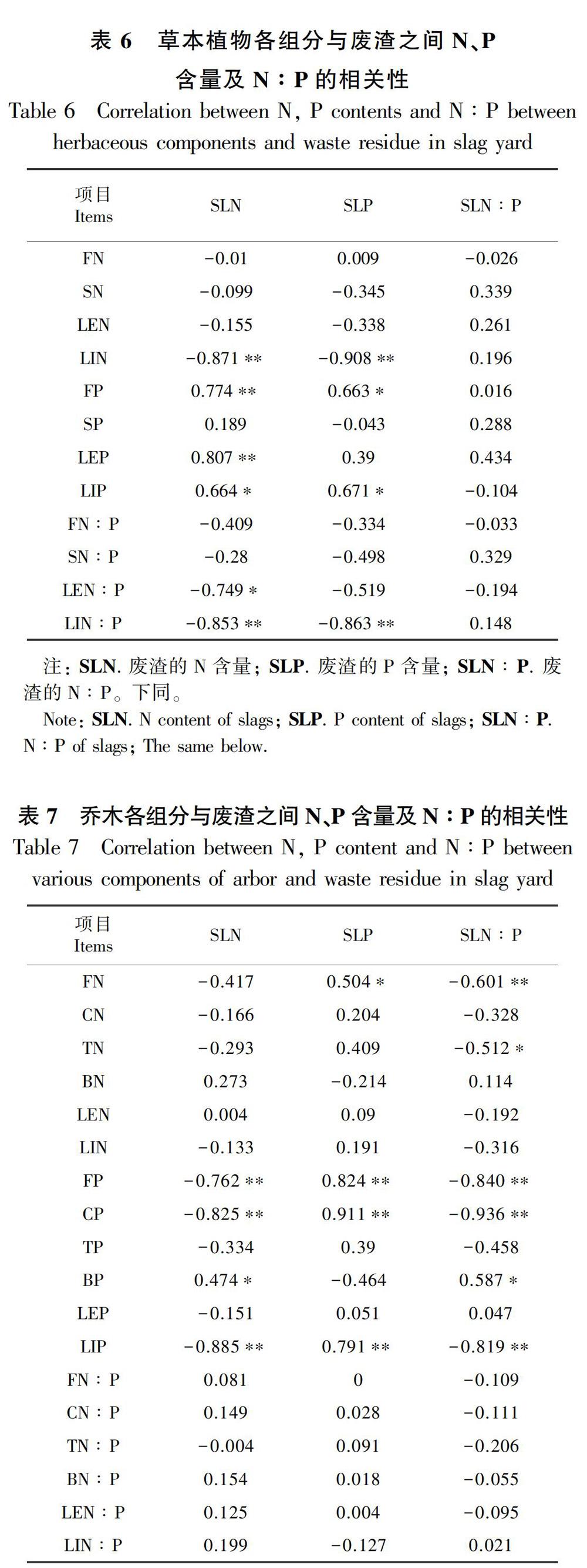
表4结果表明,两种草本植物的细根和茎中N∶P与N含量之间呈极显著正相关关系(P<0.01),叶片和凋落物中N∶P與N含量之间存在显著或极显著的正相关关系(P<0.05),与P含量之间呈极显著负相关关系(P<0.01)。由表5可知,相比研究中的两种草本植物,三种乔木各组分的N、P、N∶P之间联系更紧密。乔木不同组分的N含量之间存在极显著正相关关系(P<0.01),枝和叶片中P含量与各组分的N含量之间呈极显著负相关关系(P<0.01),分析各组分中N∶P发现,乔木各组分中N∶P均与N含量之间均呈极显著正相关关系(P<0.01), 与枝和叶的P含量之间存在极显著负相关关系(P<0.01)。相比植物各组分中P含量,各组分中N的分布与植物的N、P含量及N∶P之间相关性更强,说明该生态修复系统中植物的生长主要受N限制。
分别对两种草本植物和三种乔木中N、P含量及N∶P与废渣之间的关系进行相关性分析 (表6,表7)得出,草本植物的凋落物中N含量、N∶P均与废渣的N、P含量呈极显著负相关关系(P<0.01),凋落物中P含量与废渣的N、P含量呈显著正相关关系(P<0.05),细根的P含量也与废渣N、P含量呈显著正相关关系(P<0.05),草本植物各组分中N、P含量、N∶P均与废渣N∶P相关性不显著。三种乔木的细根、粗根和凋落物的P含量与废渣的N含量、N∶P均呈极显著负相关关系(P<0.01),而与废渣P含量呈极显著正相关关系(P<0.01)。纵观五种植物各组分与废渣的关系,只有根和凋落物与废渣之间具有更明显的相关性。
3 讨论
3.1 渣场修复植物不同器官及凋落物中C、N、P的含量特征
C、N、P元素是陆地生态系统中生物生长和生态过程的重要限制因子,也是生态化学计量学研究关注的主要元素(马永跃和王维奇,2011)。本研究中,五种修复植物各器官中C含量波动较小,在421.64~468.76 g·kg-1之间,与六种喀斯特林地植物的C含量接近(427.5 g·kg-1)(曾昭霞等,2015),但低于黄土高原西部针叶林各器官的C含量均值(501.1 g·kg-1)(孙美美等,2017)。N和P在各植物器官(细根、粗根、茎/干、枝、叶)中的含量具有显著差异,其平均值分别为7.69、1.80 g·kg-1,其中N含量低于喀斯特林地植物(21.2 g·kg-1)(曾昭霞等,2015),也低于全国753种陆地植物(18.6 g·kg-1)(Han et al.,2005),P含量高于喀斯特林地植物(1.2 g·kg-1)(曾昭霞等,2015)。与前人的研究结果相比,本研究中五种修复植物中C、N含量较低,可能是由于植物在非生长季(冬季)处于休眠状态,叶片光合作用降低,对养分的需求减少,进而使合成的有机产物减少,植物对养分的吸收也相应减少,另外,不同植物类型、生长环境以及植物自身节律等因素也可能是造成植物中C、N、P含量差异的重要因素。
就不同类型植物各器官中C、N、P含量而言,两种草本植物的根与三种乔木的细根中C、P含量接近,平均含量分别为425.84、2.35 g·kg-1,C含量均低于武夷山黄山松(陈晓萍等,2018)和中国植物细根的平均值(马玉珠等,2015),P含量均高于前两者的研究;N在植物细根中的含量为两种草本植物(6.36 g·kg-1)明显低于三种乔木(12.36 g·kg-1), 也低于前两者的研究,三种乔木细根中N含量则显著高于前两者的研究,对于出现不同类型植物和不同区域植物间的养分含量差异,一方面可能是植物结构特征和生长节律不同,植物的营养元素含量差异,另一方面,可能是由不同的采样时间与不同的生境带来的。枝和茎/干是植物重要的养分储存器官(Wilson,2011),五种修复植物茎/干的C含量与马任甜等(2017)研究的刺槐茎中C含量接近,N、P含量均低于前者的研究结果。五种植物叶片中C、N含量较稳定,其平均含量分别為421.64、13.92 g·kg-1,低于喀斯特峰丛洼地森林乔木(496.15、15.88 g·kg-1)(俞月凤等,2014),也低于黑岱沟露天煤矿上生长的优势植物叶片C含量(477.6、22.39 g·kg-1)(马任甜等,2016),这可能是由于样品采集时间为非生长季,植物呼吸作用消耗的C大于光合作用积累的C,同时废渣堆场生态修复年限尚短,生境仍然较恶劣,能为植物生长提供的养分有限,所以叶片中C、N含量较低。五种修复植物叶片的P含量在中国植物叶片P含量范围内(0.05~10.27 g·kg-1)(Han et al.,2005),高于俞月凤等(2014)和马任甜等(2016)的研究结果,但本研究中废渣的P含量较低,这可能有两方面的因素,在废渣基质方面,因为土法炼锌废渣养分贫瘠,在实现生态修复前向废渣中添加了改良基质,生态修复中植物吸收的磷可能主要来源于改良基质,但改良基质给植物提供磷的量有限,植物生长几年后,改良基质中磷可能已被植物利用殆尽,剩余的废渣中的磷含量就呈现较低的状况;在植物方面,废渣堆场实现生态修复年限较短,废渣的基质条件不同于一般的喀斯特土壤,植物在废渣的重金属及低磷胁迫下根系应激产生更多质子或有机酸(张振海等,2011;Dresler et al, 2014),从而增加了根际环境介质中磷的溶解性或生物有效性,增加植物对磷的吸收,此外,低磷胁迫下植物磷酸酶活性增强也可能使植物中P含量较高(柯野等,2018)。
植物生长所需的氮磷有90%来源于往年植物归还到土壤中的养分,凋落物作为生态系统中养分归还的主要形式,对维持土壤肥力和促进生态系统正常的物质循环和养分平衡有重要作用。本研究中,草本植物和乔木的凋落物的C、N、P含量较稳定,其平均含量分别为401.56、9.36、2.06 g·kg-1,其中C含量与喀斯特林地植物凋落物(396.2 g·kg-1)相当(曾昭霞等,2015),但低于喀斯特刺梨林和核桃林(422.67、433.89 g·kg-1)(王璐等,2018),N、P则分别低于和高于前两者的研究,凋落物中低C、N高P的分配格局在一定程度秉承了植物中C、N、P的分配特点。
刘超等(2012)研究表明,植物各器官都具有其独特的结构物质和功能属性,同一植物不同器官的C、N、P含量存在差异。在土荆芥和芦竹中,C含量为茎>叶>根>凋落物,在三种乔木中C含量为干>枝>细根>粗根>叶>凋落物,N、P含量在两种草本植物中的分布不完全一致,其中N含量为叶>凋落物>根>茎,P含量分布则为叶>根>凋落物>茎,在三种乔木中N、P分布均为叶>细根>凋落物>粗根>枝>茎/干,其整体规律与赵亚芳等(2014)研究结果一致。因为植物的茎/干和枝主要起支撑地上部分和运送水分、无机盐离子的功能,新陈代谢较叶片慢,所以N、P含量比叶片低(Martin et al.,1998)。茎/干和枝中具有较高的C含量可能是由于前期合成并储存于植物枝干的碳物质含量高于植物枝干向根部运送所亏损的量。三种乔木细根的C、N、P含量均高于粗根,与马玉珠等(2015)关于全国细根碳氮磷的研究结论一致,这主要与细根的生理功能有关,细根是植物地下部分最活跃且对土壤环境变化最敏感的部分,是植物从土壤中获取物质和能量的主要器官(孙文泰等,2016),由于在非生长季,植物蒸腾作用较弱,细根吸收的N、P元素未及时转移至植物的其他器官而富集在细根部分。本研究中土荆芥、芦竹、构树和柳杉的凋落物中P含量均低于叶片,这与聂兰琴等(2016)、陆远鸿等(2019)的研究结论一致,而刺槐凋落物的P含量高于叶片,这与赵月等(2016)对非生长季杉木不同器官中营养元素的研究结果相似。植物叶片中N、P平均含量在所有组分中最高,这是因为叶片是植物光合作用的主要器官,其新陈代谢旺盛,对N、P的需求较大。
在不同類型植物中,C、N、P的含量及化学计量特征的差异反应了不同植物对C、N、P的适应策略,结构性元素C在五种植物各器官中的含量表现为柳杉显著高于其他植物,限制性元素N、P的含量相反,基本表现为柳杉最低,因为柳杉属于常绿植物,也是针叶类植物,相比其他几种植物,柳杉对C、N、P的适应策略明显不同,这与皮发剑等人(2017)的研究结论相似,即C在不同类型植物中的含量表现为常绿树种高于落叶树种,针叶树种高于阔叶树种,而N、P的含量则相反。
3.2 渣场修复植物不同器官及凋落物中C∶N、C∶P、N∶P特征
在同种植物不同营养器官中,C、N、P的分布特征与化学计量比对研究物种的生态策略和环境适应性有重要的生态学和植物生理学意义(贺合亮等,2017)。本研究中,两种草本植物的根与三种乔木的细根的C∶N分别为68.80和45.37,与武夷山黄山松细根接近(陈晓萍等,2018),三种乔木细根和粗根的C∶N均略低于全国植物细根和粗根的C∶N(59.15和121.98),而C∶P和N∶P均远低于中国植物细根和粗根的比值(马玉珠等,2015),这与五种修复植物中P含量较高有关。修复植物细根的C∶N、C∶P小于粗根,可能是细根较粗根具有更大的比表面积,生理活性更强,生命周期较短,对N、P的需求较大。植物叶片N∶P被广泛应用于判断系统中植物生长的限制因子。Koerselman & Meuleman(1996)研究表明,当植物叶片N∶P<14时,植物生长受N限制,N∶P>16时,植物生长受P限制,14
潘复静等(2011)研究表明凋落物的N∶P是制约凋落物分解和养分循环的重要因子,N∶P较高则凋落物分解缓慢,反之,凋落物分解较快,尤其当N∶P>25或P含量小于0.22 g·kg-1时,凋落物分解更难(Güsewell & Verhoeven,2006)。五种植物凋落物的N∶P(2.79~9.00)均小于25,说明五种修复植物凋落物的理论分解速率较快,但植物凋落物的分解速率还与重金属污染、微生物种群及其活性密切相关(薛银婷等,2018),因此判断系统中凋落物的分解速率还需进一步研究。比较植物各营养器官与凋落物中C∶N、C∶P和N∶P发现,两种草本植物与三种乔木各组分的C∶N、C∶P均与植物各组分中N、P的分布特征呈完全相反的规律,说明N、P含量是引起植物C∶N和C∶P变化的主要因子,这与牛得草等(2013)
的研究结论一致,也符合C作为植物结构性元素,N、P为植物限制性元素的基本特性。植物各组分中N∶P的主要控制因子可由相关性分析得出,两种草本植物各组分中N∶P均与其N、P含量的相关性达到显著或极显著水平,说明土荆芥和芦竹的N∶P由N和P共同控制,而三种乔木各组分的N∶P与N含量之间达到极显著正相关,同时与枝和叶的P含量呈显著或极显著负相关,说明研究区三种乔木各组分中的N∶P主要受N含量控制,同时植物的枝和叶中的P含量也会影响N∶P。
3.3 废渣中C、N、P含量及化学计量特征
五种修复植物对应的表层废渣中C、N、P含量分别为107.47、2.8、0.76 g·kg-1,其中C含量高于桂西北喀斯特森林土壤(曾昭霞等,2015),N、P含量低于前者,这与土法炼锌以煤炭为燃料和还原剂的工艺技术有关,较高的有机碳含量可能是冶炼过程中未完全燃烧或氧化的煤炭造成的假象,且后续的研究也证实,废渣中有机碳全量较高,但具有活性的易氧化有机碳占废渣有机碳不足20%。废渣中较低的N、P含量可能是由于废渣自身养分含量低(邢容容等,2018),且修复时间短暂(6 a),根系对废渣养分的改善有限,废渣中微生物活性较低,凋落物分解释放的养分有限。两种不同类型植物修复下表层废渣中C、N、P含量均表现为草本植物修复作用下低于乔木,因为乔木相比草本植物具有更高的生物量,会产生更大量的凋落物回归废渣,从而增加废渣中养分的含量。
土壤中C、N、P比值可以指示土壤中有机质组成和预测有机质分解速率(王绍强和于贵瑞,2008)。本研究中,五种修复植物对应的表层废渣中C∶N、C∶P、N∶P分别为39.25、147.85、3.91。其中:废渣中C∶N高于广西喀斯特土壤(15.3)(曾昭霞等,2015);C∶P低于喀斯特峰丛洼地不同森林类型土壤(209.2)(俞月凤等,2014);N∶P接近原生林土壤(3.9)(曾昭霞等,2015),但远低于喀斯特峰丛洼地土壤(23.8)(俞月凤等,2014)。产生不同差异可能是研究的植被类型、土层深度、采样时节、研究区域分布等因素不同造成的。Bengtsson et al.(2003)研究指出,土壤C∶N与土壤中有机质的分解速率成反比,当C∶N高时,微生物需要输入N素满足自身生长,分解速率降低,反之,C∶N较低时,多余的N会进入土壤中,微生物分解速率提高。本研究中,废渣C∶N相比其他的研究结果偏高,可能是废渣中较高的有机碳假象造成的,不能准确指示废渣中有机质的分解快慢。
4 结论
通过对土法炼锌废渣堆场生态修复系统中五种修复植物的营养器官-凋落物-废渣体系中C、N、P含量及化学计量特征进行研究,结论如下:
(1)渣场五种修复植物不同器官的C含量较稳定,在土荆芥和芦竹中,C含量为茎>叶>根>凋落物,在构树、柳杉和刺槐中为干>枝>细根>粗根>叶>凋落物;N、P在植物中分布差异显著,在土荆芥和芦竹中,N含量为叶片>凋落物>根>茎,P含量为叶片>根>凋落物>茎,在构树、柳杉和刺槐中N、P的分布一致,均为叶>细根>凋落物>粗根>枝>干。废渣中C、N、P含量均低于植物各器官和凋落物(茎/干中P含量除外),草本植物修复下废渣中C、N、P含量低于乔木修复下的含量。
(2)兩种不同类型的植物各组分中C∶N、C∶P的大小与N、P 在各组分中的分布呈相反的规律,说明植物中C∶N和C∶P主要由N和P含量决定;相关性分析得出,土荆芥和芦竹的N∶P由N、P共同决定,构树、柳杉和刺槐的N∶P主要受N含量控制,也受枝和叶中P含量影响。
(3)五种修复植物中只有豆科植物刺槐的生长受P限制,其余植物生长均受N限制,说明豆科植物刺槐更能适应贫瘠的废渣基质环境;此外,相关性分析也得出植物在该系统中生长主要受N限制,所以在往后的贫瘠金属冶炼废渣的修复中,建议优先选择豆科植物作为先锋植物,从而达到缓解基质养分贫瘠,改善基质植生条件的作用。
(4)对废渣-植物-凋落物体系中N、P、及N∶P进行相关性分析得出,植物细根和凋落物与废渣的养分之间关系密切。
参考文献:
BENGTSSON G, BENGTSON P, KATARINA F MANSSON, 2003. Gross nitrogen mineralization, immobilization, and nitrification rates as a function of soil C/N ratio and microbial activity [J]. Soil Biol Biochem, 35(1):143-154.
CHEN XP, GUO BQ, ZHONG QL, et al.,2018. Response of fine root carbon, nitrogen, and phosphorus stoichiometry to soil nutrients in Pinus taiwanensis along an elevation gradient in the Wuyi mountains [J]. Acta Ecol Sin, 38(1):273-281. [陈晓萍,郭炳桥,钟全林,等,2018. 武夷山不同海拔黄山松细根碳、氮、磷化学计量特征对土壤养分的适应 [J]. 生态学报,38(1):273-281.]
DRESLER S, HANAKA A, BEDNAREK W, et al., 2014. Accumulation of low-molecular-weight organic acids in roots and leaf segments of Zea mays plants treated with cadmium and copper [J]. Acta Physiol Plant, 36:1565-1575.
ELSER JJ, BRACKEN MES, CLELAND EE, et al., 2007. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems [J]. Ecol Lett, 10(12):1135-1142.
FAN HB, WU JP, LIU WF, et al., 2015. Linkages of plant and soil C∶N∶P stoichiometry and their relationships to forest growth in subtropical plantations [J]. Plant Soil, 392(1/2): 127-138.
GSEWELL S, VERHOEVEN JTA, 2006. Litter N∶P ratios indicate whether N or P limits the decomposability of grami-noid leaf litter [J]. Plant Soil, 287:131-143.
HAN WX, FANG JY, GUO DL, et al., 2005. Leaf nitrogen and phosphorus stoichiometry across 753 terrestrial plant species in China [J]. New Phytol, 168:377-385.
HE HL, YANG XC, LI DD, et al., 2017. Stoichiometric characteristics of carbon, nitrogen and phosphorus of Sibiraea angustata shrub on the eastern Qinghai-Xizang Plateau [J]. Chin J Plant Ecol, 41(1):126-135. [贺合亮, 阳小成, 李丹丹, 等, 2017. 青藏高原东部窄叶鲜卑花碳、氮、磷化学计量特征 [J]. 植物生态学报, 41(1):126-135.]
HERBERT DA, MATHEW W, EDWARD BR, 2003. A model analysis of N and P limitation on carbon accumulation in amazonian secondary forest after alternate land-use abandonment [J]. Biogeochemistry, 65(1):121-150.
KE Y, XIE L, LAN L, et al., 2018. Absorption of poorly soluble phosphorus by sugarcane seedlings and its response to low phosphorus stress [J]. Guihaia, 39(12):1673-1680. [柯野, 谢璐, 蓝林, 等, 2018. 甘蔗幼苗对难溶性磷的吸收及其根系对低磷胁迫的响应 [J]. 广西植物, 39(12): 1673-1680.]
KOERSELMAN W, MEULEMAN AFM, 1996. The vegetation N∶P ratio:A new tool to detect the nature of nutrient limitation [J]. J Appl Ecol, 33(6):1441-1450.
LIU C, WANG Y, WANG N, et al., 2012. Advances research in plant nitrogen, phosphorus and their stoichiometry in terrestrial ecosystems:A review [J]. Chin J Plant Ecol, 36(11): 1205-1216. [刘超, 王洋, 王楠, 等, 2012. 陆地生态系统植被氮磷化学计量研究进展 [J]. 植物生态学报, 36(11):1205-1216.]
LU YH, CAO Y, XU LM, et al., 2019. Stoichiometric characteristics of plants, litter and soils in desertification area of Poyang Lake [J]. Chin J Ecol, 38(2):329-335. [陸远鸿, 曹昀, 许令明, 等, 2019. 鄱阳湖沙化土地植物-凋落物-土壤化学计量特征 [J]. 生态学杂志, 38(2):329-335.]
LUO YF, WU YG, QIU J, et al., 2019. Suitability of four woody plant species for the phytostabilization of a zinc smelting slag site after 5 years of assisted revegetation [J]. J Soils Sed, 19(2):702-715.
LUO YF, WU YG, WANG H, et al., 2018. Bacterial community structure and diversity responses to the direct revegetation of an artisanal zinc smelting slag after 5 years [J]. Environ Sci Poll Res, 25:14773-14788.
MA RT, AN SS, HUANG YM, 2017. C, N and P stoichiometry characteristics of different-aged Robinia pseudoacacia plantations on the loess plateau, China [J]. Chin J Appl Ecol, 28(9): 2787-2793. [马任甜, 安韶山, 黄懿梅, 2017. 黄土高原不同林龄刺槐林碳、氮、磷化学计量特征 [J]. 应用生态学报, 28(9):2787-2793.]
MA RT, FANG Y, AN SS, et al., 2016. Ecological stoichiometric characteristics of leaves and litter of plants dominant in Heidaigou opencast coal mining area [J]. Acta Pedol Sin, 53(4):1003-1014. [马任甜, 方瑛, 安韶山, 等, 2016. 黑岱沟露天煤矿优势植物叶片及枯落物生态化学计量特征 [J]. 土壤学报, 53(4):1003-1014.]
MA YY, WANG WQ, 2011. Carbon, nitrogen and phosphorus content and the ecological stoichiometric ratios of paddy field soil-plants in Minjiang River estuary [J]. Subtr Agric Res, 7(3): 182-187. [马永跃, 王维奇, 2011. 闽江河口区稻田土壤和植物的C、N、P含量及其生态化学计量比 [J]. 亚热带农业研究, 7(3):182-187.]
MA YZ, ZHONG QL, JIN BJ, et al., 2015. Spatial changes and influencing factors of fine root carbon, nitrogen and phosphorus stoichiometry of plants in China [J]. Chin J Plant Ecol, 39(2):159-166. [马玉珠, 钟全林, 靳冰洁, 等, 2015. 中国植物细根碳、氮、磷化学计量学的空间变化及其影响因子 [J]. 植物生态学报, 39(2):159-166.]
MARTIN J G, KLOEPPEL BD, SCHAEFER TL, et al., 1998. Aboveground biomass and nitrogen allocation of ten deciduous southern appalachian tree species [J]. Can J For Res, 28(11):1648-1659.
NIE L Q, WU Q, YAO B, et al., 2016. Leaf litter and soil carbon, nitrogen, and phosphorus stoichiometry of dominant plant species in the Poyang Lake wetland [J]. Acta Ecol Sin, 36(7):1898-1906. [聂兰琴, 吴琴, 尧波, 等, 2016. 鄱阳湖湿地优势植物叶片-凋落物-土壤碳氮磷化学计量特征 [J]. 生态学报, 36(7):1898-1906.]
NIU DC, LI Q, JIANG SG, et al., 2013. Seasonal variations of leaf C∶N∶P stoichiometry of six shrubs in desert of Chinas Alxa Plateau [J]. Chin J Plant Ecol, 37(4):317-325. [牛得草, 李茜, 江世高, 等, 2013. 阿拉善荒漠区6种主要灌木植物叶片C∶N∶P化学计量比的季节变化 [J]. 植物生态学报, 37 (4):317-325.]
PAN FJ, ZHANG W, WANG KL, et al., 2011. Litter C∶N∶P ecological stoichiometry character of plant communities in typical karst peak-cluster depression [J]. Acta Ecol Sin, 31(2): 335-343. [潘復静, 张伟, 王克林, 等, 2011. 典型喀斯特峰丛洼地植被群落凋落物C∶N∶P生态化学计量特征 [J]. 生态学报, 31(2):335-343.]
PARTON W, SILVER WL, BURKE IC, et al., 2007. Global-scale similarities in nitrogen release patterns during long-term decomposition [J]. Science, 315:361-364.
PI FJ, SHU LX, YU LF, et al, 2017. Study on ecological stoichiometry characteristics and correlation of plants within different organs of 10 dominant tree species in Karst Region of Central Guizhou [J]. Ecol Environ Sci, 26(4):628-634. [皮发剑, 舒利贤, 喻理飞, 等, 2017. 黔中喀斯特10种优势树种根茎叶化学计量特征及其关联性 [J]. 生态环境学报, 26(4):628-634.]
SUN MM, GUAN JH, YUE JW, et al., 2017. Carbon, nitrogen, and phosphorus stoichiometry in plant organs and soil of coniferous forests on the western Loess Plateau [J]. J Soil Water Cons, 31(3):202-208. [孙美美, 关晋宏, 岳军伟, 等, 2017. 黄土高原西部针叶林植物器官与土壤碳氮磷化学计量特征 [J]. 水土保持学报, 31(3):202-208.]
SUN WT, DONG T, LIU XL, et al., 2016. The relationship between root distribution of apple and soil physical properties by different ground covering approaches [J]. Agric Res Arid Areas, 34(2):88-95. [孙文泰, 董铁, 刘兴禄, 等, 2016. 覆盖处理苹果细根分布与土壤物理性状响应关系研究 [J]. 干旱地区农业研究, 34(2):88-95.]
WANG HF, ZHANG YF, ZHANG ZK, 2000. The current situation of and counterpart measures for the zinc industry of Guizhou [J]. Guizhou Sci, 18(3):229-233. [王海峰, 张元福, 张中可, 2000. 贵州锌工业现状及发展对策 [J]. 贵州科学, 18(3):229-233.]
WANG L, YU YH, XING RR, et al., 2018. Ecological stoichiometry characteristics of carbon, nitrogen, phosphorus, and potassium of different economic tree species in the karst frigid and arid area [J]. Acta Ecol Sin, 38(15):5393-5403. [王璐, 喻阳华, 邢容容, 等, 2018. 喀斯特高寒干旱区不同经济树种的碳氮磷钾生态化学计量特征 [J]. 生态学报, 38(15):5393-5403.]
WANG M, MURPHY MT, MOORE TR, 2014. Nutrient resorption of two evergreen shrubs in response to long-term fertilization in a bog [J]. Oecologia,174(2):365-377.
WANG SQ, YU GR, 2008. Ecological stoichiometry characte-ristics of ecosystem carbon, nitrogenand phosphorus elements [J]. Acta Ecol Sin, 28(8):3937-3947. [王绍强, 于贵瑞, 2008. 生态系统碳氮磷元素的生态化学计量学特征 [J]. 生态学报, 28(8):3937-3947.]
WILSON BF, 2011. Tree branches as populations of twigs [J]. Can J Bot, 67(2):434-442.
XING RR, WU YG, LUO YF, et al., 2018. Effect of pioneer phytoremediation on indigenous zinc smelting slag matrix nutrients and microbial properties [J]. Res Soil Water Cons, 25(5):103-111. [邢容容, 吴永贵, 罗有发, 等, 2018. 先锋修复植物对土法炼锌废渣基质养分及微生物学特性的影响 [J]. 水土保持研究, 25(5):103-111.]
XUE YT, LIN YH, HE XB, et al., 2018. Effects of lead on the decomposition of Phyllostachys pubescens leaf litter in western Hunan Province [J]. J Chongqing Norm Univ (Nat Sci Ed), 35(1):117-123. [薛银婷, 林永慧, 何兴兵, 等, 2018. 铅污染对湘西地区毛竹凋落物分解的影响 [J]. 重庆师范大学学报(自然科学版), 35(1):117-123.]
YU Q, ELSER JJ, HE NP, et al., 2011. Stoichiometric homeostasis of vascular plants in the Inner Mongolia grassland [J]. Oecologia, 166(1):1-10.
YU YF, PENG WX, SONG TQ, et al., 2014. Stoichiometric characteristics of plant and soil C, N and P in different forest types in depressions between karst hills, Southwest China [J]. Chin J Appl Ecol, 25(4):947-954. [俞月凤, 彭晚霞, 宋同清, 等, 2014. 喀斯特峰丛洼地不同森林类型植物和土壤C、N、P化学计量特征 [J]. 应用生态学报, 25(4):947-954.]
ZENG ZX, WANG KL, LIU XL, et al., 2015. Stoichiometric characteristics of plants, litter and soils in karst plant communities of Northwest Guangxi [J]. Chin J Plant Ecol, 39(7): 682-693. [曾昭霞, 王克林, 刘孝利, 等, 2015. 桂西北喀斯特森林植物-凋落物-土壤生态化学计量特征 [J]. 植物生态学报, 39(7):682-693.]
ZHANG ZH, CHEN Y, HAN SF, et al., 2011. Effect of P deficiency stress on soybean root system and its secretion of H+ and organic acid [J]. Chin J Soil Crop Sci, 33(2):35-40. [张振海, 陈琰, 韩胜芳, 等, 2011. 低磷胁迫对大豆根系生长特性及分泌H+和有机酸的影响 [J]. 中国油料作物学报, 33(2):35-40.]
ZHAO Y, WANG GJ, CHEN C, et al., 2016. Relationship of N∶P stoichiometry of different organs and soil of Cunninghamia lanceolata in Huitong [J]. J Centr S Univ For Technol, 36(11):73-79. [趙月, 王光军, 陈婵, 等, 2016. 杉木不同器官与土壤的N∶P生态化学计量相关性 [J]. 中南林业科技大学学报, 36(11):73-79.]
ZHAO YF, XU FL, WANG WL, et al., 2014. Seasonal variation in contents of C, N and P and stoichiometry characteristics in fine roots, stems and needles of Larix principis-rupprechtii [J]. Chin Bull Bot, 49(5):560-568. [赵亚芳, 徐福利, 王渭玲, 等, 2014. 华北落叶松根茎叶碳氮磷含量及其化学计量学特征的季节变化 [J]. 植物学报, 49(5):560-568.]
(责任编辑 蒋巧媛)
