低温胁迫对红果风铃木幼苗生理特性的影响
2020-10-20丁释丰袁森黄稚清秦新生冯志坚
丁释丰 袁森 黄稚清 秦新生 冯志坚
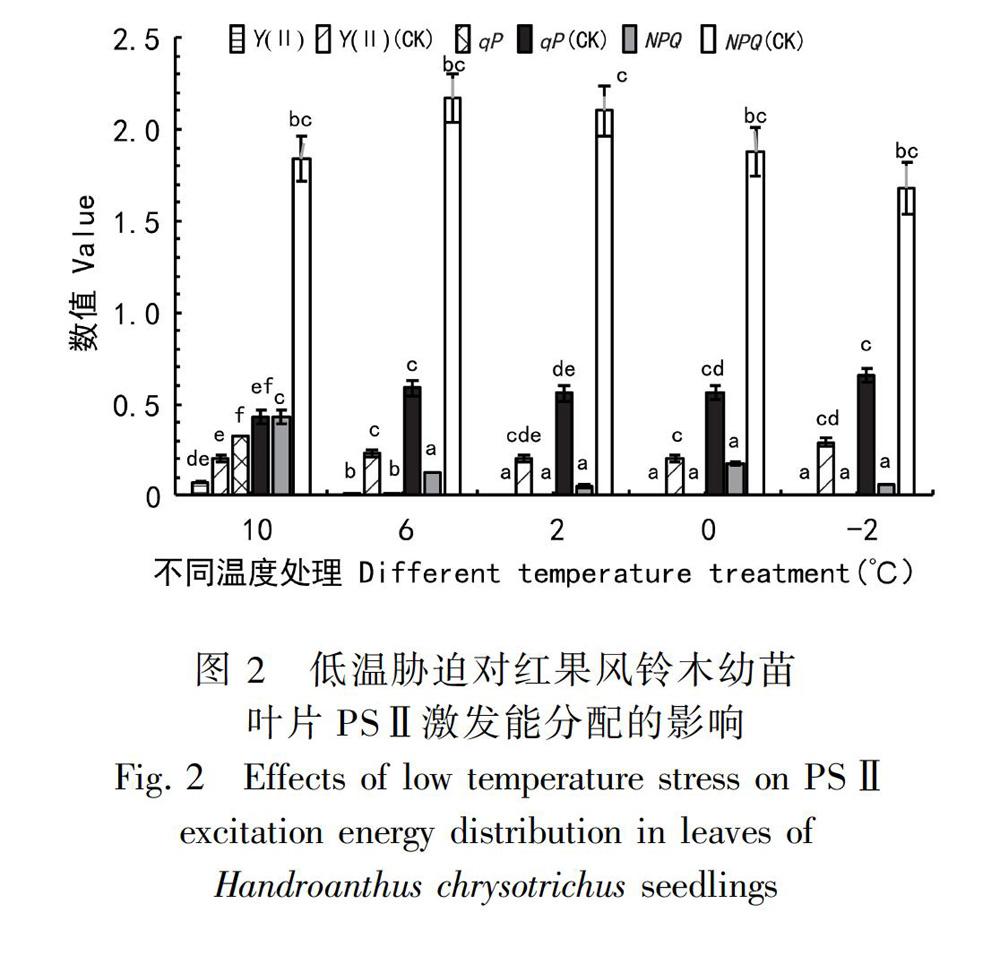
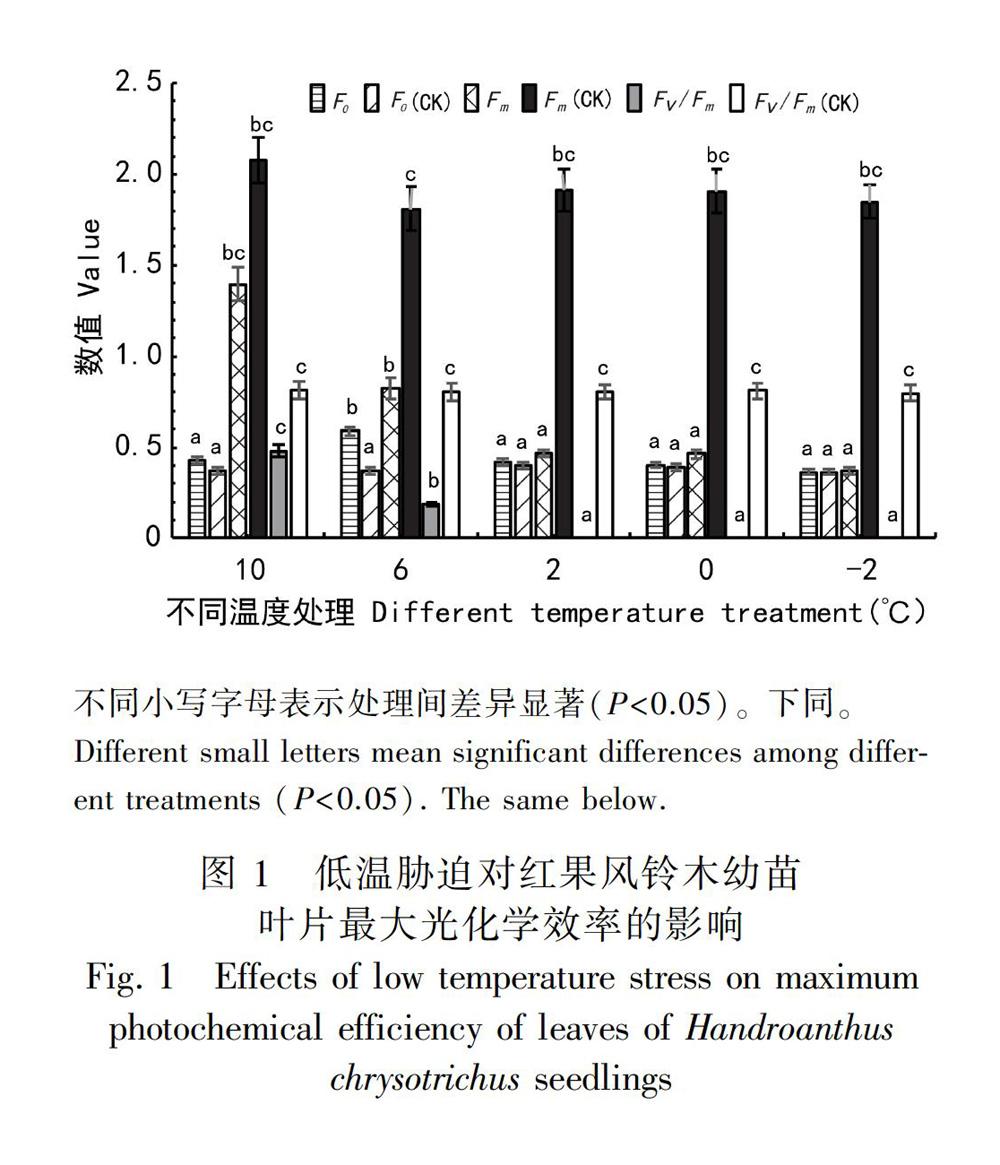
摘 要:为探究红果风铃木(Handroanthus chrysotrichus)对低温的耐受能力,以1年生红果风铃木幼苗为材料,人工模拟不同低温环境观察幼苗在低温下的受害情况,并测定SOD、MDA等生理指标和叶绿素荧光参数。结果表明:红果风铃木幼苗在遭受6 ℃及以上低温胁迫时,能够通过提高SOD活性、增加SS、Pro以及叶绿素含量增强植株抗寒力,进而缓解胁迫给植株带来的伤害;红果风铃木幼苗还能够提高NPQ耗散过剩的光能避免光合机构受低温破坏,但随着温度下降至2 ℃,PSⅡ反应中心的伤害愈发严重,光合作用能力受到显著影响。结合各项指标变化特征,推测红果风铃木幼苗能够有效通过自身调节缓解胁迫带来伤害的最低温度范围为2~5 ℃,具体临界点还需进一步验证。在遭受6 ℃及以上低温胁迫时,红果风铃木幼苗主要通过提高SOD活性和积累SS缓解低溫给植株带来的损伤;随着温度下降至2 ℃及以下低温时,主要通过提高SOD活性和积累Pro缓解低温给植株带来的损伤。综合分析认为,红果风铃木幼苗无法抵抗0 ℃及以下低温产生的冻害,但对0 ℃以上低温有一定的耐受力,研究结果有利于红果风铃木的抗性品种选育和推广种植,对探索植物抗寒生理机制具有一定意义。
关键词:红果风铃木, 低温胁迫, 生理指标, 叶绿素荧光参数
中图分类号:Q945
文献标识码:A
文章编号:1000-3142(2020)08-1151-08
Abstract:In order to explore the tolerance and reaction mechanism of Handroanthus chrysotrichus towards coldness, one year seedlings of H. chrysotrichus were put under artificially controlling low temperature treatments. Physiological indexes and chlorophyll fluorescence parameters were measured. The result shows that under low temperature stress of 6 ℃ or above, seedlings can improve their cold tolerance and protect themselves by enhancing SOD activities, SS contents and Pro contents. In addition, seedlings can also prevent their photosynthetic system from cold damaging through improving NPQ. When the temperature reaches to 2 ℃, both PSⅡ reaction center and photosynthetic system will be badly damaged. From these changes, we deduce that seedlings of H. chrysotrichus can resist to low temperature ranging 2-5 ℃ and prevent the meselves from coldness through self-adjustment. However, the critical point still needs to be determined. Our paper also finds out that when facing low temperature 6 ℃ or above, SS contents rapidly rise up. When the temperature reaches to 2 ℃ or below, Pro contents remarkably rise up. Perhaps seedlings of H. chrysotrichus can protect themselves from low temperature 6 ℃ or above by enhancing SOD activities and SS contents. Seedlings of H. chrysotrichus will mitigate damages of low temperature 2 ℃ or below by enhancing SOD activities and Pro contents. It is concluded that the seedlings of H. chrysotrichus could not resist the freezing damage caused by low temperature below 0 ℃, but have certain tolerance to low temperature above 0 ℃. The results are beneficial to both selecting resistent varieties and promoting cultivation. It is also good for exploring cold tolerant ability of plants.
Key words:Handroanthus chrysotrichus, low temperature stress, physiological index, chlorophyll fluorescence parameters
红果风铃木(Handroanthus chrysotrichus)属紫葳科(Bignoniaceae)风铃木属(Handroanthus),具有良好的观赏价值,春季黄花满树,夏秋枝叶繁茂,冬季枝干优美,在华南地区道路绿化、园林造景中应用广泛。近年来,对红果风铃木的研究多集中于栽培技术(Bittencourt & Giorgi, 2010;Sampaio et al., 2013;Dos et al., 2017)、观赏价值(Garofalo & Regalado, 2006;Rebelatto et al., 2013)。在生理研究方面,Brun et al.(2017)对城市中红果风铃木体内的微量营养元素水平进行了分析,Moura et al.(2016)研究了气候对红果风铃木体内蛋白质和碳水化合物含量的影响,但其对于逆境的耐受能力与响应机制的研究还处于一片空白。因此,对红果风铃木的耐寒能力及响应机制进行研究,既弥补了空白,有利于其耐寒品种的选育,又促进红果风铃木生产和推广。
当环境温度低于临界温度时,植物会受到伤害,温度越低植物受害越重。目前,国内外关于低温胁迫的研究越发详细,我国植物生理特性受低温胁迫影响的研究从最初主要集中于生理生化方面,逐步转向分子水平研究层面(杨阳等,2018)。应对低温胁迫植物的生理机制及其抗性表现在平衡渗透压、清除活性氧、光合效率以及叶片超微结构变化等多个方面(李文明等,2017)。但植物生理生化受低温胁迫的影响,因种类、地理、温度、时长以及温度降幅等不同而不同,且温度的变化能引起其他环境因子的变化,针对植物在应对多复合环境因子胁迫的响应还有待于进一步深入研究。我国华南地区分布的植物多为热带或亚热带植物,在冬季植株常遇寒潮而受冷害或冻害,严重时会导致植株死亡造成直接经济损失。因此,对华南地区主要经济树种的抗寒性进行研究,有利于抗性品种选育和种质改良创新,还可为植物抗性生理机制研究提供科学依据。
1 材料与方法
1.1 实验设计
材料为长势一致的1年生红果风铃木容器苗(广东国森林业有限公司提供)。共设置10 ℃(D1组)、6 ℃(D2组)、2 ℃(D3组)、0 ℃(D4组)、-2 ℃(D5组)5个实验组(D组),每组3个重复、每个重复3盆幼苗,对照组(CK组)在自然条件下培育。
用FYS-10智能人工气候室模拟低温环境,在48 h内从30 ℃缓冲到10 ℃(每6 h降低2.5 ℃)。10 ℃处理48 h后,取出D1组于常温下培育24 h后测定相关指标,并记录幼苗受害表现。其余4组继续在气候室中6 ℃处理48 h后,取出D2组测定其指标并记录幼苗受害表现,依次类推。样株采样结束后,在自然条件恢复培养30 d,观察植株脱离胁迫环境后的恢复情况。
1.2 指标测定
超氧化物歧化酶(SOD)活性测定采用氮蓝四唑(NBT)光还原法;可溶性糖(SS)含量测定采用蒽酮法;游离脯氨酸(Pro)含量测定采用酸性茚三酮显色法;丙二醛(MDA)含量测定采用硫代巴比妥酸(TBA)法(史树德等,2011)。
叶绿素荧光参数的测定用PAM-2500便携式调制叶绿素荧光仪,使用90°角光纤适配器暗反应30 min后,采用饱和脉冲模式,测量其慢速动力学曲线,测量时间为300 s。测定的指标包括最小荧光产量(F0)、最大荧光产量(Fm)、PSⅡ的最大光合量子产量(Fv/Fm)、PSⅡ的实际光合量子产量[Y(Ⅱ)]、光化学淬灭系数(qP)和非光化学淬灭系数(NPQ)。
2 结果与分析
2.1 低温胁迫对红果风铃木幼苗形态的影响
对不同低温处理下的红果风铃木幼株形态变化进行观察记录(表1),低温胁迫下红果风铃木幼苗生长受到显著影响。其中,10 ℃低温胁迫下,红果风铃木幼苗便已受到明显伤害,主要表现为组织柔软,部分叶片萎蔫,并伴随脱水症状;随着胁迫程度加深,2 ℃低温胁迫下植株受到的伤害显著加深超1/2的叶片萎蔫,部分叶片脱水严重;-2 ℃低温胁迫下,植株全部叶片萎蔫,植株脱水严重,甚至干枯濒死。10 ℃是华南冬季寒潮常见低温,红果风铃木幼苗在这一阶段就已受到明显伤害,初步判断其对低温的耐受能力较差。
经30 d的复培,6 ℃及以上低温处理的组别可较快恢复正常生长;2 ℃组的植株虽然存活,但长势较差,说明2 ℃低温给植株带来了较重伤害,需较长时间才能够恢复;0 ℃组与-2 ℃组的植株在复培中无法恢复,植株死亡(图版 I)。推测红果风铃木原产于热带地区,生长环境温度多在0 ℃以上,导致其对冻害的耐受能力差。
2.2 低温胁迫对红果风铃木幼苗生理指标的影响
低温条件会影响植物的膜脂过氧化作用,使叶片中MDA含量增加,进而改变叶片内组织的生理结构,使其失去原有生理活性和功能,最终导致植物受到损害(李倩等,2016)。SOD是植物体内重要的保护酶,低温环境下能够避免超氧自由基对膜的伤害(Kanazawa et al., 2010)。随着低温胁迫程度的加深,红果风铃木幼苗叶片内MDA的含量呈不断增加的趋势,在10、6 ℃低温处理下平缓上升(P>0.05),在2、0和-2 ℃低温处理下迅速上升,且与CK组差异显著(P<0.05)。紅果风铃木幼苗叶片SOD活性呈现明显的波动(表 2),从10 ℃开始逐渐增强,但在2 ℃骤然下降至未低温处理前水平,又在-2 ℃时达到最大值(P<0.05)。综合分析MDA含量与SOD活性的变化,可以发现在6 ℃及以上低温胁迫下,红果风铃木幼苗能够有效通过体内的保护酶清除活性氧,但在2 ℃及以下低温胁迫下植株受害程度显著加深, 推断红果风铃木幼苗能够有效通过自身调节缓解低温胁迫带来伤害的临界范围在2~5 ℃之间。
为了各项生理过程在解除低温胁迫后能够正常进行,植物会在体内产生脯氨酸、可溶性糖等大量低分子化合物(郝建军等,2007)。可溶性糖是植物体内重要的渗透调节物质之一,植物体内的淀粉在遭受低温胁迫时会转化成可溶性糖以降低冰点缓解低温对膜系统的伤害,从而提高抗寒性(赵慧等,2017)。红果风铃木幼苗叶片中的SS含量在低温胁迫前期迅速上升,在10、6 ℃低温处理下迅速上升(P<0.05),随后出现波动,各实验组SS含量均显著高于CK组。低温处理下,红果风铃木幼苗叶片中的Pro含量和SS含量的变化相似,呈现出先上升后些微下降的趋势(表2),说明Pro的积累有利于提高植物的抗寒能力。低温处理开始后,Pro含量迅速上升,并在2 ℃时迅速达到了最大值(较6 ℃组上升了119.61%),随后又缓慢下降,各实验组的Pro含量均显著高于对照组(P<0.05)。
2.3 低温胁迫对红果风铃木叶片叶绿素含量的影响
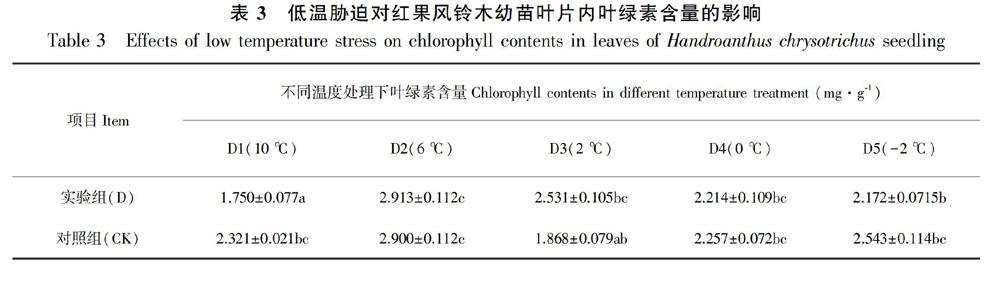
低温胁迫能加剧叶绿体的降解,并抑制其合成,因此叶绿素含量的稳定性有利于光合作用的进行,从而增强对低温胁迫的忍耐力(何跃君等,2008)。随着温度的下降,红果风铃木幼苗叶片内的叶绿素含量呈现出一定的波动(表 3)。10 ℃组植株叶片中的叶绿素含量较CK组有显著下降(P<0.05),但6 ℃组较10 ℃组显著上升了58.03%(P<0.05),与CK组含量接近。此后,在2 ℃之后叶绿素含量虽略高于CK组(P>0.05),但低温抑制了叶绿素色素合成酶的活性,叶绿素的合成速度小于被降解速度,-2 ℃组叶绿素含量已经明显低于CK组,植株全部叶片萎蔫失绿,濒临死亡。
2.4 低温胁迫对红果风铃木幼苗叶片叶绿素荧光参数的影响
2.4.1 低温胁迫对红果风铃木幼苗叶片最大光化学效率的影响 叶绿素荧光信号具有丰富的光合作用信息,利用叶绿素荧光参数研究植物在胁迫条件下的光合作用可以推测出环境胁迫对植物的影响程度(Schubert et al., 2006),反映植株生长势和抗性强弱(杨猛等,2012)。随着处理温度的不断降低,红果风铃木幼苗F0不断上升,在6 ℃时达到最大值(P<0.05)随后下降,并在-2 ℃时降到最小值。F0的上升表明低温胁迫影响叶片光能的吸收,降低了原初光能转化效率,引起了光抑制;Fm呈不断下降的趋势,与CK组差异显著,2 ℃时降幅最大,表明在2 ℃及以下低温,PSⅡ反应中心受到了严重的损坏;Fv/Fm常用于度量植物叶片PSⅡ原初光能转换效率,随着处理温度的不断降低,红果风铃木幼苗的Fv/Fm不断降低,并在2 ℃时下降到零点,随后一直为零。比较各处理温度下的数值,发现10 ℃实验组较CK组显著下降了39.23%(P<0.05),6 ℃实验组比10 ℃实验组显著下降了64.50%(P<0.05)。此外,Fv/Fm为零表明植株的光合电子传递效率为零,致使PSⅡ反应中心激发能积累过剩,进而损伤PSⅡ反应中心,导致叶片光合作用能力下降(图 1)。
2.4.2 低温胁迫对红果风铃木叶片 PSⅡ激发能分配的影响 qP在一定程度上反映植物对光能的利用程度(Genty et al., 1989),随着处理温度的不断降低,红果风铃木幼苗叶片的qP也不断下降,各实验组的结果均显著低于同期CK组(P<0.05),并在2 ℃时降到零点。NPQ反映的是PSⅡ反应中心吸收的光能无法用于电子传递而通过热能散失掉的份额,将过剩光能耗散出来,可以避免对光合机构的损伤(张雷明等,2003;陈世茹等,2011)。随着处理温度的降低,红果风铃木幼苗叶片的NPQ也不断下降,10 ℃组比CK组显著下降了87.70%(P<0.05),随后持续缓慢下降(P>0.05),表明非辐射耗散不断减少。Y(Ⅱ)反映植物光合电子传递速率的快慢(And & Weis, 2003),Y(Ⅱ)和Fv/Fm的变化呈高度一致,两者都在10 ℃和6 ℃时显著降低,在2 ℃时降至零点,说明PSⅡ反应中心的实际光能转换效率在低温下受到了抑制,红果风铃木的光合机构受到破坏(图 2)。
3 讨论与结论
低温条件下,植物体内的生理生化反应会出现异常,主要表现在植物的渗透压调节机制、保护酶体系、活性氧平衡等方面(Ruelland et al., 2009;李猛等,2018)。活性氧和自由基造成植物体内的活性氧平衡被破坏,膜脂过氧化反应加剧导致植物体内MDA大量积累,膜功能受阻或丧失,造成代谢紊乱,从而对细胞产生伤害。有研究表明,乔木在低温胁迫条件下体内MDA的含量会显著增加(王纬航,2017;陈凯等,2019)。一方面,在低温胁迫下,红果风铃木幼苗体内MDA大量积累,因此造成了植株的伤害,这与前人的研究结果一致。另一方面,低温胁迫下为了维持植物体内活性氧和自由基的动态平衡,减轻自身所受伤害,植物体内的保护酶体系会发挥作用清除这些有害物质,使SOD等保护酶活性增强大,但超过保护酶的作用范围时其酶活性会下降(赵慧等,2017)。张文娇和王小德(2011)研究发现梅花在其低温耐受范围内,SOD的酶活性持续上升;章锦涛等(2017)在研究山茶的抗寒能力也发现随着胁迫温度的降低,植株体内SOD活性先升高后降低。红果风铃木体内的SOD等保护酶活性显著上升,仅在2 ℃时活性较低,随后酶活性又继续上升,说明遭受低温胁迫时,SOD是红果风铃木自我调节的主要物质之一,但其在遭遇临界点以下低温时反应具有一定的滞后性。在低温胁迫条件下,通过渗透压调节物质来调节植物体内细胞液浓度可帮助植物更好应对低温胁迫。雷雪峰等(2019)研究禾本科牧草中,发现植株体内Pro含量随着温度的降低而显著增强。SS、Pro等渗透压保护物质含量迅速增加以提高细胞液的浓度,从而降低细胞质的冰点,保护细胞质胶体不致遇冷凝固。植物到达抗寒性临界点后,SOD等酶活性会降低(庄倩倩等,2018),本研究中红果风铃木幼苗SOD活性在2 ℃骤然下降,结合其余指标的变化特征,推测红果风铃木幼苗能够有效通过自身调节缓解胁迫带来伤害的最低温度范围为2~5 ℃,具体临界点还需进一步实验验证。本研究还发现,在遭受6 ℃及以上低温胁迫时,红果风铃木体内SS含量快速上升,当温度下降至2 ℃及以下低溫胁迫时,随着不同的Pro合成途径开放,Pro含量显著增加。据此推测,红果风铃木幼苗在遭受6 ℃及以上低温胁迫时主要通过增强SOD活性和积累SS缓解低温给植株带来的损伤;随着温度下降至2 ℃及以下低温,主要通过增强SOD活性和积累Pro缓解低温给植株带来的损伤。
此外,红果风铃木幼苗在6~10 ℃低温条件下可以通过提高叶绿素的含量提高植物代谢能力来补偿逆境受到的伤害,进而提高耐受低温的能力,但随着胁迫温度的降低和时间的延长,高强度的低温胁迫会损伤光合色素,导致叶绿体结构分解、淀粉粒消失、质壁分离、细胞器崩溃降解等现象,进而对植物体造成损害(李晓靖和崔海军,2018)。由于叶绿素的分解,进而影响了叶绿素荧光参数。前人研究也有发现,低温胁迫使当归和白及叶片叶绿素含量以及Y(Ⅱ) 、PSⅡ、光化qP等叶绿素荧光参数显著降低(崔波等,2019;张牡丹等,2019),损害PS反应。本研究中,在低温胁迫下,红果风铃木幼苗叶片的光合作用能力也受到了显著影响。在轻度低温胁迫时红果风铃木幼苗能够提高NPQ耗散过剩的光能保护光合机构,但在6 ℃时Fv/Fm、Y(Ⅱ)、NPQ和qP开始急剧下降,Fv/Fm、Y(Ⅱ)和qP更是在2 ℃降至零点,说明PSⅡ反应中心的原初光能转换效率和实际光能转换效率在低温下都受到了极大的抑制,且在2 ℃以下对PSⅡ反应中心的伤害愈发严重。
综合低温胁迫对红果风铃木幼苗生理特征的影响,并结合其脱离胁迫后的复培情况,表明红果风铃木幼苗在遭受低温胁迫后具有一定的自我修复机制,对2 ℃及以上低温具有较强的耐受性,即具有一定抗冷性;对0 ℃及以下的低温所引起的冻害没有忍耐力。红果风铃木在华南地区进行种植推广时,冬季应做好必要的防护措施,避免低于2 ℃及以下低温带来经济价值和观赏价值的损失。
参考文献:
AND GHK, WEIS E, 2003. Chlorophyll fluorescence and photosynthesis:The basics [J]. Ann Rev Plant Physiol, 42(42):313-349.
BITTENCOURT NS, GIORGI MCI, 2010. Self-fertility and polyembryony in South American yellow trumpet trees (Handroanthus chrysotrichus and H. ochraceus, Bignoniaceae):A histological study of postpollination events [J]. Plant Syst Evol, 288(1-2):59-76.
BRUN FGK, BRUN EJ, DIONATAN G, et al., 2017. Nutrition facts and limits for micronutrients in tree species used in urban forestry [J]. An Acad Brsil Cienc, 89(3):1881-1893.
CHEN K, JIANG XZ, MA SY, et al., 2019. Physiological response of leaves to low temperature stress and evaluation of cold resistance in seedling stage of Parashorea chinensis Wang Hsie. [J]. J Univ NW For, 34(3):67-73. [陈凯,江秀章,马松亚,等,2019. 望天树苗期叶片对低温胁迫的生理响应及抗寒性评价 [J]. 西北林学院学报,34(3):67-73.]
CHEN SR, YU LQ, YI J, et al., 2011. Influence of chlorophyll fluorescence characteristics on alfalfa seedlings under cryogenic stress [J]. Acta Prat Sin, (4):596-600. [陈世茹,于林清,易津,等,2011. 低温胁迫对紫花苜蓿叶片叶绿素荧光特性的影响 [J]. 草地学报,(4):596-600.
CUI B, CHENG SL, YUAN XY, et al., 2019. Effects of low temperature stress on photosynthesis and chlorophyll fluorescence parameters of Bletilla striata (Thunb.) Reichb.f. [J]. J Trop Crop, 40(5):891-897. [崔波,程邵丽,袁秀云,等,2019. 低温胁迫对白及光合作用及叶绿素荧光参数的影响 [J]. 热带作物学报,40(5):891-897.]
DOS SRSM, SILVERIRA RLR, STEFANEL CM, et al., 2017. Agar does not affect in vitro rhizogenesis and ex vitro acclimatization of Handroanthus chrysotrichus [J]. Cerne, 23(2):185-191.
GAROFALO J, REGALADO R, 2006. Long-term survial of golden trumpet tree and copperpod on rockland soil in South Florida [J]. Hortscience, 41(3):502-502.
GENTY B, BRIANTAIS J, BAKER NR, 1989. The relationship between the quantum yield of photosynthetic electron transport and quenching of chlorophyll fluorescence [J]. Bba-Gen Subjects, 990(1):87-92.
HAO JJ, KANG ZL, YU Y, 2007. Experimental techniques of plant physiology [M]. Beijing:Chemical Industry Press. [郝建军,康宗利,于洋,2007. 植物生理学实验技术 [M]. 北京:化学工业出版社.]
HE YJ, XUE L, REN XR, et al., 2008. Effects of low temperature stress on physiological characteristics of six seedlings [J]. Chin J Ecol, 27(4):524-531. [何躍君,薛立,任向荣,等,2008. 低温胁迫对六种苗木生理特性的影响 [J]. 生态学杂志,27(4):524-531.]
KANAZAWA S, SANO S, KOSHIBA T, et al., 2010. Changes in antioxidative enzymes in cucumber cotyledons during natural senescence:comparison with those during dark-induced senescence [J]. Physiol Plant, 109(2):211-216.
LEI XF, MA AS, LI HC, et al., 2010. Comparison of physiological response and cold resistance of 8 species of Gramineae forage under low temperature stress [J]. Jiangsu Agric Sci, 9(51):1-5. [雷雪峰,马爱生,李海翠,等,2019. 8种禾本科牧草低温胁迫的生理响应及抗寒性比较 [J]. 江苏农业科学,9(51):1-5.]
LI M, L TH, XIN QJ, et al., 2018. Research progress on evaluation and regulation of low temperature tolerance of melon vegetables [J]. Acta Hort Sin, 45(9):1761-1777. [李猛,吕亭辉,邢巧娟,等,2018. 瓜类蔬菜耐低温性评价与调控研究进展 [J]. 园艺学报,45(9):1761-1777.]
LI Q, WANG YY, WANG YX, 2016. Effects of temperature stress onchlorophyll content, soluble sugar and MDA in Trifolium repens L. [J]. Chin Agric Sci Bull, 32(19):83-86. [李倩,王月异,王玉祥,2016. 低温处理对白三叶叶绿素、可溶性糖和丙二醛等的影响 [J]. 中国农学通报,32(19):83-86.]
LI WM, XIN JP, WEI CY, et al., 2017. Research progress in cold resistance of plants [J]. Jiangsu Agric Sci, 45(12):6-11. [李文明,辛建攀,魏驰宇,等,2017. 植物抗寒性研究进展 [J]. 江苏农业科学,45(12):6-11.]
LI XJ, CUI HJ, 2018. Research progress on the physiological response of plants to environmental stress [J]. J Shandong For Sci Technol, 48(6):90-94. [李晓靖,崔海军,2018. 低温胁迫下植物光合生理研究进展 [J].山东林业科技,48(6):90-94.]
MOURA BB, ALMEIDA PH, BALLIANA AG, et al., 2016. Effects of changes in micro-weather conditions on structural features, total protein and carbohydrate content in leaves of the Atlantic rain forest tree golden trumpet (Tabebuia chrysotricha) [J]. Braz J Biol, 77(3):535-541.
REBELATTO D, LEAL TDS, MORAES CPD, 2013. Fenologia de duas espécies de ipê em área urbana do município de araras, so paulo, brasil [J]. Revsbau, 8(1):1-16.
RUELLAND E, VAULTIER MN, ZACHOWSKI A, et al., 2009. Cold signalling and cold acclimation in plants [J]. Adv Bot Res, 49:35-150.
SAMPAIO DS, COSTA ME, MENDES RC, 2013. Temperature effect in the number of seedlings per seed in cultivated specimens of Handroanthus chrysotrichus (Bignoniaceae) [J]. Iheringia Serie Bot, 68(2):279-283.
SCHUBERT H, ANDERSSON M, SNOEIJS P, 2006. Relationship between photosynthesis and non-photochemical quenching of chlorophyll fluorescence in two red algae with different carotenoid compositions [J]. Mar Biol, 149(5):1003-1013.
SHI SD, SUN YQ, WEI L, 2011. Experimental guidance of plant physiology [M]. Beijing:China Foretry Publish. [史樹德,孙亚卿,魏磊,2011. 植物生理学实验指导 [M]. 北京:中国林业出版社.]
WANG WH, 2017. Physiological studies on drought and cold resistance of Sophora of Sophora japonica, Sophora japonica Linn. var. japonica f. pendula Hort., Sophora japonica L. cv. Golden Stem and Sophora safflower [D]. Hohhot:Inn Mongolia Agricultural University. [王纬航, 2017. 槐属观赏乔木国槐、龙爪槐、金枝国槐和红花国槐抗旱性抗寒性生理研究 [D]. 呼和浩特:内蒙古农业大学.]
YANG M, WEI L, ZHUANG WF, et al., 2012. Effects of low-temperature stress on electric conductivity and fluorescence parameters of maize seedling [J]. J Maize Sci, 20(1):90-94. [杨猛,魏玲,庄文锋,等,2012. 低温胁迫对玉米幼苗电导率和叶绿素荧光参数的影响 [J]. 玉米科学,20(1):90-94.]
YANG Y, WANG HL, GAO P, 2018. Effects of low-temperature stress on leaf morphology and physiological characteristics of columbine [J]. Jiangsu Agric Sci, 46(23):151-153. [杨阳,王红丽,高平,2018. 低温胁迫对耧斗菜叶片形态和生理特性的影响 [J]. 江苏农业科学,46(23):151-153.]
ZHANG JT, WANG H, WANG S, et al., 2017. Effect of external application of abscisic acid on physiological and biochemical indexes of Camellia oleifera under low temperature stress [J]. J Anhui Agric Univ, 44(1):142-145. [章锦涛,王华,王松,等,2017. 外施脱落酸对低温胁迫下山茶花生理生化指标的影响 [J]. 安徽农业大学学报,44(1):142-145.]
ZHANG LM, SHANGGUANG ZP, MAO MC, et al., 2003. Effect of long-term nitrogen application on chlorophyll fluorescence parameters of dryland wheat during grain filling stage [J]. Chin J Appl Ecol, 14(5):695-698. [张雷明,上官周平,毛明策,等,2003. 长期施氮对旱地小麦灌浆期叶绿素荧光参数的影响 [J]. 应用生态学报,14(5):695-698.]
ZHANG MD, FENG Y, SHI ZZ, et al., 2019. Regulation of chlorophyll content and fluorescence characteristics of Angelica sinensis seedlings by extracellular ATP under drought and low temperature stress [J]. J Trad Chin Med, 44(7):1305-1313. [張牡丹,冯媛,石珍珍,等,2019. 干旱和低温胁迫下细胞外ATP对当归幼苗叶绿素含量及其荧光特性的调节 [J]. 中国中药杂志,44(7):1305-1313.]
ZHANG WJ, WANG XD, 2011. Effects of low temperature stress on physiological characteristics of five different plum blossom varieties [J]. Jiangsu Agric Sci, 39(3):203-205. [张文娇,王小德,2011. 低温胁迫对5个不同梅花品种生理特性的影响 [J]. 江苏农业科学,39(3):203-205.]
ZHAO H, ZHAO YB, GUO JB, et al., 2017. Research progress on tolerance of plants to low temperature stress [J]. Seed, 36(5):47-50. [赵慧,赵一博,郭江波,等,2017. 植物耐受低温胁迫研究进展 [J]. 种子,36(5):47-50.]
ZHUANG QQ, CHEN SP, LIU HZ, 2018. Effects of low temperature stress on three physiological indexes of root system from Hosta ventricosa [J]. J NE For Univ, 46(3):33-36. [庄倩倩,陈少鹏,刘洪章,2018. 低温胁迫对紫萼玉簪根系3种生理指标的影响 [J]. 东北林业大学学报,46(3):33-36.]
(责任编辑 周翠鸣)
