调控油茶果生刺盘孢bZIP转录因子CfAp1的生物学功能*
2020-10-17高亚兰何苑皋
高亚兰 何苑皋 李 河
(南方人工林病虫害防控国家林业和草原局重点实验室 森林有害生物防控湖南省重点实验室经济林培育与保护省部共建教育部重点实验室 中南林业科技大学 长沙 410004)
油茶(Camelliaoleifera)是我国特有的木本食用油料树种,与油棕、油橄榄和椰子并称为世界四大木本食用油料植物。油茶主要分布于我国长江流域以南的高山及丘陵地带,与粮食作物生态位互补;另外还有保持水土、涵养水源、调节气候等各种生态功能,在我国区域经济中具有重要地位(陈永忠等2013)。油茶种子可榨油供食用,是我国特有的优质植物油,富含角鲨烯和黄酮类物质等多种生理活性物质,具有降血脂、降胆固醇、延缓动脉粥样硬化、增强人体免疫力等营养保健作用(许俊道, 2018)。
油茶炭疽病是我国油茶种植区的主要病害,可引起油茶大量落果、落蕾,甚至整株衰亡,对油茶生长、产量和品质具有严重影响。我国油茶产区每年因炭疽病造成油茶籽减产20%左右,重病区可达40%以上。油茶病果种子含油量仅为健康种子的一半,甚至更低(靳爱仙等, 2009)。笔者课题组于2014年首次报道果生刺盘孢(Colletotrichumfructicola)是油茶炭疽病的病原菌(李河等, 2014),进一步研究发现我国油茶炭疽病是由多种刺盘孢属真菌侵染所致,病原菌种类主要有果生刺盘孢(C.fructicola)、暹罗刺盘孢(C.siamense)、 胶孢刺盘孢(C.gloeosporioides)、 山茶刺盘孢(C.camelliae)和卡斯特刺盘孢(C.karstii)等,其中果生刺盘孢分离率最高,为70%左右,分布在全国90%以上的油茶种植区(Lietal., 2016; 李河等, 2016; 2017; 李河, 2018)。致病力测试结果表明,果生刺盘孢对油茶叶的致病力最强 (李河等, 2019)。上述研究结果表明果生刺盘孢是我国油茶炭疽病的主要流行致病菌。
目前,主要采用化学杀菌剂防治油茶炭疽病(陈绍红等, 2007)。但由于病原菌易变,我国油茶炭疽病原菌已产生比较严重的抗药性(李河等, 2019)。因此,阐明油茶炭疽病原菌的致病分子机制,对开发新的杀菌剂靶标具有重要意义。然而,调控植物病原菌致病的基因网络十分复杂,往往多种基因协同作用。bZIP(Basic leucine zipper)转录因子是普遍存在于动植物及微生物中的一类转录因子,包含参与寡聚化作用的亮氨酸拉链区以及与特异DNA序列相结合碱性区,这2个区域以二聚体的形式结合DNA。bZIP转录因子在病原真菌中大量存在,主要参与调控病菌生长发育和致病等过程(Natorffetal., 2003; Etxebesteetal., 2008; Guoetal., 2010; Xiaoetal., 2010)。本文拟以油茶果生刺盘孢菌bZIP转录因子CfAp1为对象,研究其生物学功能,为揭示其致病分子机制提供参考。
1 材料与方法
1.1 供试菌株
野生型(wild type,WT)菌株为笔者实验室首次分离鉴定的油茶炭疽病优势流行致病菌果生刺盘孢菌CFLH16菌株,前期已完成全基因组测序。突变体及回补菌株为本试验获得。
1.2 CfAP1基因序列分析
根据稻瘟病菌(Magnaportheoryzae)中MoAp1蛋白的氨基酸序列(MGG_12814)在果生刺盘孢菌全基因组数据库中进行Blastp分析,获得与其同源的CfAp1蛋白序列。根据CfAp1蛋白的氨基酸序列在NCBI数据库中获得其他物种的同源蛋白的氨基酸序列,利用Mega7.0软件构建系统发育树。
1.3 CfAP1基因敲除突变体的获得
根据同源重组原理构建CfAP1基因敲除载体。以野生型菌株CFLH16的基因组DNA为模板,分别用引物对bZIP45-1F/bZIP45-2R和bZIP45-3F/bZIP45-4R进行PCR扩增获得上、下臂片段,PCR产物经纯化后作模板;利用引物Hyg F/Hyg R扩增潮霉素抗性筛选基因HPH,最后以上下臂片段和潮霉素基因片段为模板,以引物对bZIP45-1F/bZIP45-4R进行over-lap PCR扩增出CfAP1基因敲除载体,经纯化后用于果生刺盘孢菌的原生质体转化。
采用PEG介导法把敲除载体片段转化至果生刺盘孢菌的原生质体中,在含有潮霉素的TB3培养基中培养3天。利用基因内引物bZIP45-7F/bZIP45-8R和臂外引物bZIP45-5F/H885R分别进行PCR扩增筛选潮霉素抗性转化子,电泳验证。引物bZIP45-7F/bZIP45-8R不能扩增出目的条带,同时满足引物bZIP45-5F/H885R可扩增出目的条带的转化子为CfAP1基因敲除突变体。本文所涉及的引物见表1。
1.4 CfAP1基因敲除突变体回补菌株的获得
设计引物对bZIP45-9F/bZIP45-10R,从果生刺盘孢菌中扩增含有启动子CfAP1基因回补片段,PCR产物经纯化后与线性pYF11共转入酵母感受态细胞XK-125形成pYF11::CfAP1互补载体,酵母细胞涂布于SD-Trp固体培养基上筛选,置于28 ℃、黑暗条件下培养3天,将SD-Trp固体培养基上长出的酵母置于YPD液体培养基中振荡培养12 h,利用引物bZIP45-7F/GFPR进行PCR鉴定阳性克隆,提取转化成功的质粒转入感受态大肠杆菌(Escherichiacoli)JM109中;经转化后的大肠杆菌细胞涂布于LB平板上,并置于37 ℃、黑暗条件下培养3天,然后将LB固体平板上长出的大肠杆菌置于LB液体培养基中振荡培养10 h,提取大肠杆菌质粒,利用引物bZIP45-9F/GFPR验证后送至测序验证;经测序验证正确后,采用PEG介导法将质粒转入ΔCfap1-8菌株原生质体中,对在含有博来霉素培养基上能生长的转化子进行绿色荧光筛选、PCR验证回补菌株。

表1 引物序列Tab.1 Primers used in this study
1.5 转录因子CfAp1的生物学表型测定
1.5.1 PDA培养基上生长测定 使用打孔器(Φ=8 mm)分别取野生型菌株CFLH16、突变体菌株ΔCfap1-8及基因回补菌株ΔCfap1/AP1的菌饼接种到PDA培养基上,并置于28 ℃黑暗倒置培养3天,每个菌株3个重复,试验重复3次。
1.5.2 突变体产孢率测定 用PDB摇瓶培养野生型菌株CFLH16、突变体菌株ΔCfap1-8及基因回补菌株ΔCfap1/AP1产孢(Wangetal., 2017)。使用血小板计数板计数法分别统计分生孢子产量,每个菌株3个重复,试验重复3次。
1.5.3 突变体对H2O2耐受性测定 将野生型菌株 CFLH16、突变体菌株ΔCfap1-8及基因回补菌株ΔCfap1/AP1活化后,从菌落边缘取菌饼(Φ=8 mm)分别接种到含0、2.5和5 mmol·L-1的H2O2的PDA培养基中央,置于28 ℃、黑暗倒置培养3天。每个菌株3个重复,试验重复3次。测量记录菌落直径,分别计算其在含不同浓度H2O2的PDA培养基上的抑制率。
1.5.4 突变体对渗透压胁迫测定 将野生型菌株 CFLH16、突变体菌株ΔCfap1-8及基因回补菌株ΔCfap1/AP1活化后,从菌落边缘取菌饼(Φ=8 mm)分别接种到含0、0.7 mol·L-1NaCl的PDA平板中央,置于28 ℃、黑暗倒置培养3天,测量记录菌落直径,计算在NaCl PDA培养基上的抑制率。每个菌株3个重复,试验重复3次。
1.5.5 突变体致病力测定 为了评估突变体菌株对油茶叶的致病力变化,将野生型菌株 CFLH16、突变体菌株ΔCfap1-8及基因回补菌株ΔCfap1/AP1活化后,从菌落边缘取菌饼(Φ=8 mm)接种至油茶叶片背面边缘,置于黑暗中28 ℃下保湿培养,观察并测量病斑大小。为明确伤口对侵染结果的影响,针刺油茶叶片,将菌丝块接种至油茶叶伤口处,黑暗中28 ℃下保湿培养,观察并测量病斑大小。每个菌株3个重复,试验重复3次。
1.5.6 突变体附着胞的形成率及膨压测定 将CFLH16、ΔCfap1-8及ΔCfap1/AP13种菌的分生孢子分别用无菌水洗涤3次,调节孢子液浓度至1×105个·mL-1,取10 μL滴至疏水玻片中央,28 ℃下保湿孵育8 h后,观察附着胞的形成并统计数量;取20 μL 2 mol·L-1的甘油浸泡附着胞,10 min后统计塌陷率。每个菌株3个重复,试验重复3次。
1.6 数据分析

图1 CfAp1结构域预测以及系统发育分析Fig.1 Domain prediction and phylogenetic analysis of CfAp1A: 通过SMART网站(http:∥smart.embl-heidelberg.de/)预测CfAp1蛋白的结构域;BRLZ为碱性亮氨酸拉链结构域,数字代表氨基酸的序号. B: Mega7.0软件的邻近相连法构建CfAp1及其他参考菌种同源蛋白氨基酸序列系统进化树,自举检验重复1 000次以获得各分支的支持率;进化树分支上的数字表示支持率;0.1为标尺,表示分支长度;每个参考菌株名称前的编号是该菌株CfAp1同源蛋白氨基酸序列的GenBank登录号。A: The structure of CfAp1 was predicted using the SMART website (http:∥smart.embl-heidelberg.de/); BRLZ: basic-leucine zipper domain; the number represents the serial number of the amino acid. B: The phylogenetic tree was constructed using the neighbour-joining (NJ) method and analysed with 1 000 bootstrap replicates in Mega7.0; the number on the branch is the support calculated with 100 bootstrap replicates in the NJ analysis; the scale bar indicates the branch length. The number in front of each reference taxon is GenBank accession number of amino acid sequence orthologous to CfAp1.
使用Microsoft Excel 2010对试验数据进行单因素方差分析,显著性差异水平设定为0.05,当P≤0.05时对比组间有差异显著性;显著性差异水平设定为0.01, 当P≤0.01时对比组间有差异极显著性。
2 结果及分析
2.1 CfAp1的鉴定及系统发育树分析
根据稻瘟菌中MoAp1蛋白的氨基酸序列(bZIP_Mo0045/MGG_12814),在油茶果生刺盘孢菌全基因组数据库中进行BLASTP比对,鉴定到1个同源的蛋白,命名为CfAp1。CfAP1基因全长1 804 bp,编码566个氨基酸。该蛋白含有1个碱性亮氨酸拉链结构域(BRLZ)和2个PAP1结构域以及2个未知功能的结构域(图1A)。系统发育分析发现,果生刺盘孢菌中的转录因子CfAp1氨基酸序列与胶孢刺盘孢菌的bZIP转录因子PAP1同源性最高(图1B),支持率高达到100%,而与稻温病菌距离较远。
2.2 CfAP1基因敲除突变体的获得及验证
敲除载体的构建策略见图2A。目标基因内引物bZIP45-7F/bZIP45-8R在野生型基因组中能扩增出单一的目的条带,突变体中扩增不出目的条带;采用目标基因上游引物bZIP45-5F和潮霉素基因上引物H855R通过PCR方法在突变体基因组中能扩增出单一的条带,而野生型中检测不到条带(图2B)。综合2对引物的检测结果,表明转化子Cfap1-8中CfAP1基因已经被成功敲除。为验证突变体的表型是否为CfAP1基因敲除引起,选择ΔCfap1-8作为受体菌株,将构建好的互补质粒导入突变体中,转化子在博莱霉素平板上筛选。抗博莱霉素的转化子通过PCR验证,获得CfAP1基因成功互补的转化子ΔCfap1/AP1。
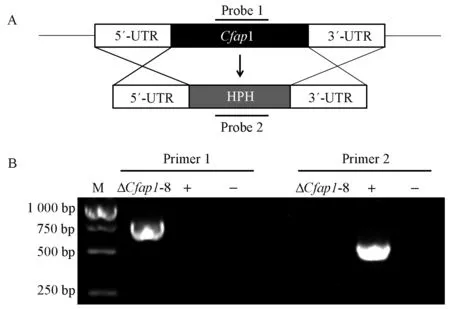
图2 CfAP1基因敲除突变体的获得Fig.2 Generation of the gene deletion mutants of CfAP1A: 基因敲除策略的模式图; B: Primer1是bZIP45-5F/H855R;Primer2是bZIP45-7F/ bZIP45-8R。M: 分子量为5 000 Marker;-: H2O阴性对照;+: WT阳性对照;ΔCfap1-8: 突变体。A: Schematic diagram of the deletion strategy for the CfAP1 gene. B: primer1 is bZIP45-5F/H855R, primer2 is bZIP45-7F/ bZIP45-8R; M: DL5000 marker; -: H2O negative control; +: WT positive control; ΔCfap1-8: mutant.
2.3 CfAP1基因对果生刺盘孢菌营养生长的影响
在PDA培养基上,果生刺盘孢菌突变体菌株ΔCfap1-8生长速率与野生型及回补菌株间无明显差异(图3A、3C),但气生菌丝显著减少(图3B)。以上结果表明CfAP1基因不影响果生刺盘孢菌的营养生长,但调控其气生菌丝的生长。
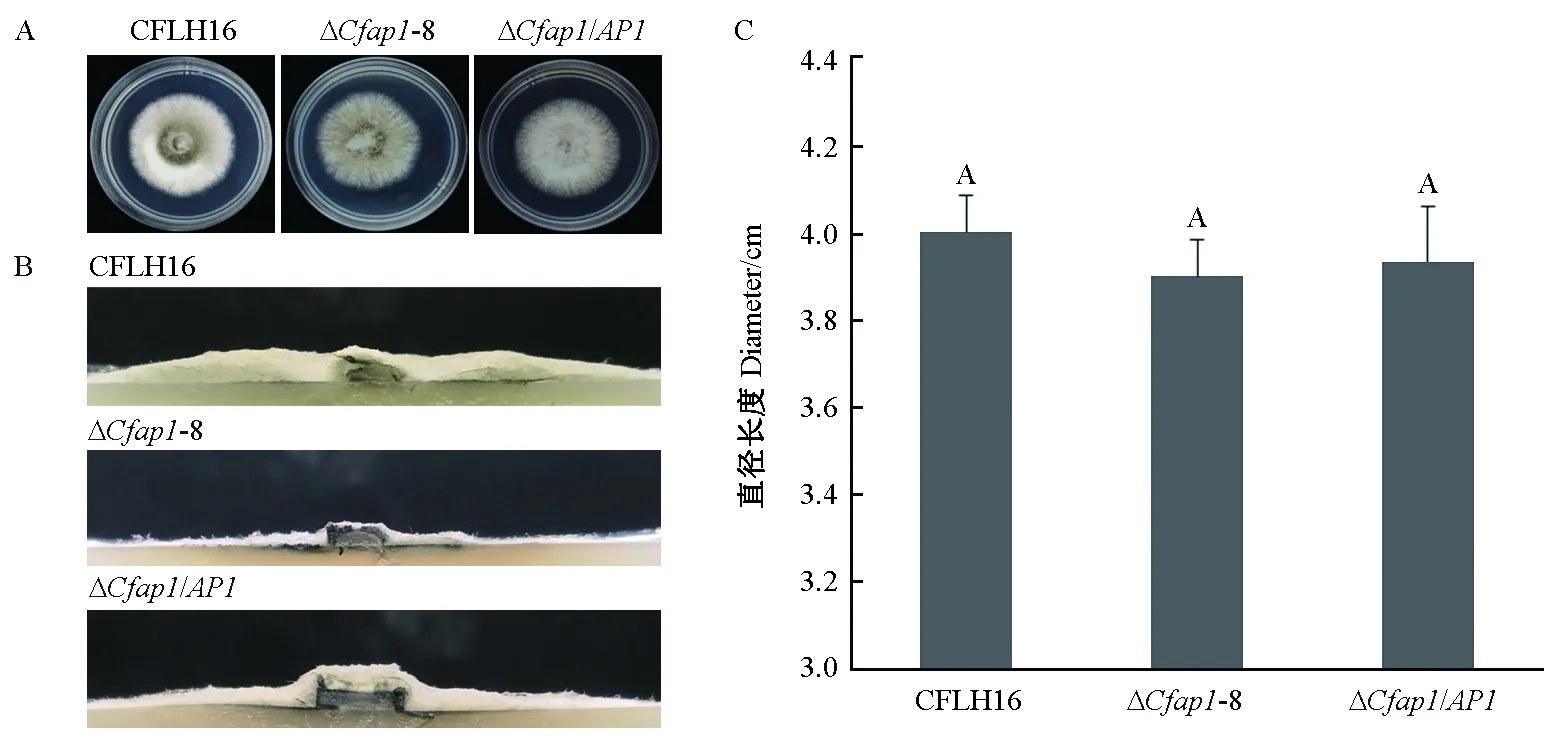
图3 CfAP1基因敲除突变体菌丝体生长Fig.3 Growth of CfAP1 gene-deletion mutantsA: 野生型菌株(CFLH16)、突变体及回补菌株在PDA培养基上28 ℃黑暗倒置培养3天的生长情况。 B: 野生型菌株(CFLH16)、突变体及回补菌株的菌落侧视图。C: 野生型菌株(CFLH16)、突变体及互补菌菌落直径差异统计分析。字母相同者表示在0.05水平上差异不显著。A: The wild-type strain (CFLH16),ΔCfap1-8, and complemented strains were inoculated on PDA media and cultured at 28 ℃ in the dark for 3 days. B: Colony side views are displayed. C: Statistical analysis of the colony diameter variations. Error bars are standard deviation and the same capitals represent no significant difference on P>0.05.
2.4 CfAP1基因参与调控果生刺盘孢菌分生孢子的形成
在真菌侵染过程中分生孢子起着十分重要的作用(Huangetal., 2016; Fangetal., 2018; Yangetal., 2018)。本文研究CfAP1基因对果生刺盘孢菌分生孢子的形成情况,结果发现突变体ΔCfap1-8只能产生较少量分生孢子,与野生型和回补菌株相比差异显著(图4),表明CfAP1基因参与调控果生刺盘孢菌分生孢子的形成。
2.5 CfAP1基因缺失降低了果生刺盘孢菌对H2O2胁迫应答
CfAP1基因敲除突变体对H2O2耐受性较野生型显著降低,突变体ΔCfap1-8在含2.5、5 mmol·L-1H2O2的PDA平板上菌丝生长受到明显抑制(图5A);敲除突变体菌丝生长抑制率较野生型及回补菌株分别增加13.5%(2.5 mmol·L-1)和22.7%(5 mmol·L-1)(图5B),表明CfAP1基因参与果生刺盘孢菌对外源H2O2胁迫的应答。
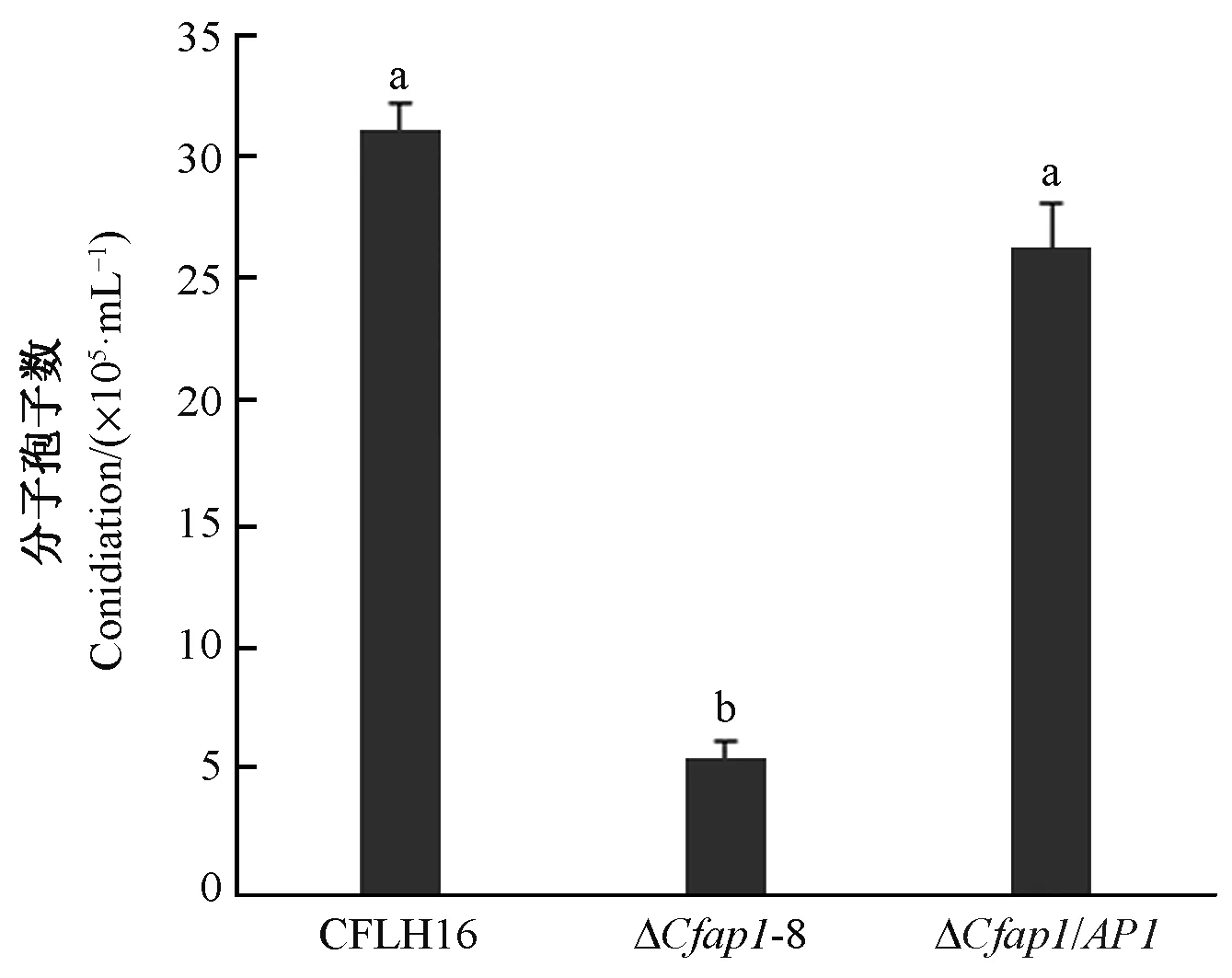
图4 CfAP1基因缺失突变体分生孢子数量统计分析Fig.4 The conidial quantitative statistical diagram of CfAP1 gene deleted mutants.不同小写字母者表示在0.01水平上差异极显著。 Error bars are standard deviation and different lowercases represent significantly difference at P≤0.01.
2.6 CfAP1基因参与应答果生刺盘孢菌的渗透压胁迫
CfAP1基因敲除突变体对NaCl耐受性较野生型显著降低,突变体ΔCfap1-8在含0.7 mol·L-1NaCl 的PDA平板上菌丝生长受到显著抑制(图6A);敲除突变体菌丝生长抑制率较野生型及回补菌株增加8.8%(图6B),这表明CfAP1基因参与应答果生刺盘孢菌的渗透压胁迫。
2.7 CfAP1基因参与调控果生刺盘孢菌的致病力
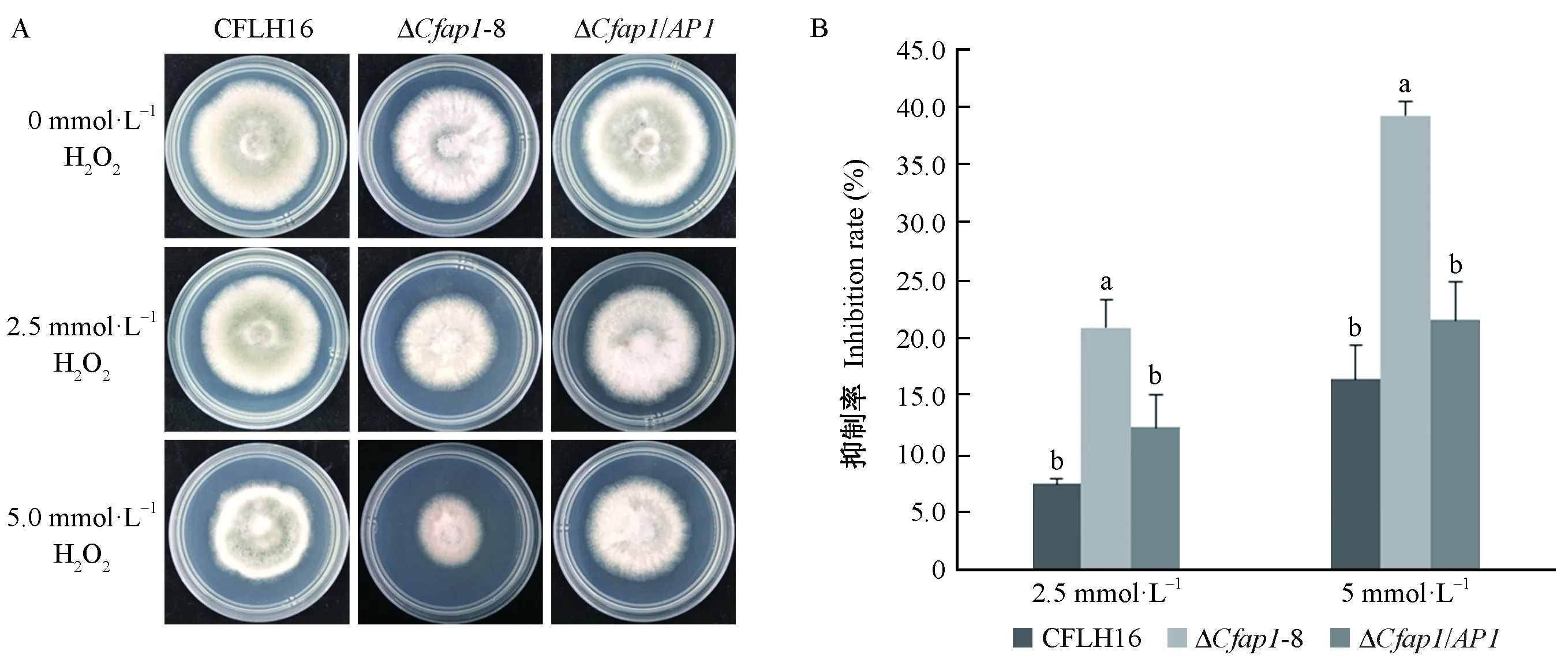
图5 CfAP1基因缺失突变对氧压胁迫的敏感性测定Fig.5 Oxygen stress sensitivity test of CfAP1 gene-deletion mutantsA: 野生型、ΔCfap1-8突变体和回补菌株分别接种于含有0、2.5和5 mmol·L-1的H2O2的PDA培养基上的菌落生长情况。 B: 菌株在不同的氧压胁迫培养基中生长抑制率统计分析. 误差线采用的是标准偏差;小写字母不同者表示在0.01水平上差异极显著.A: The wild-type strain (CFLH16),ΔCfap1-8, and complemented strains were inoculated on PDA plates containing 0、2.5 and 5 mmol·L-1 H2O2,respectively. B: Statistical analysis of the inhibition rate of the indicated strains on different stressors. Error bars are standard deviation and different lowercases represent significantly difference at P≤0.01.
无伤叶片在5天后可在接种野生型和回补菌株处观察到典型病斑,而接种ΔCfap1-8突变体菌饼只能观察到较小病斑,统计分析表明病斑直径差异显著(P≤0.01,图7A、7C)。有伤叶片3天后可在接种野生型及回补菌株伤口处观察到病斑,而突变体菌株只能引起较小的病斑,且病斑大小差异显著(P≤0.01,图7B、7D)。以上结果表明,CfAP1基因参与调控果生刺盘孢菌的致病力,并且伤口有助于病菌对油茶叶片的侵染。
2.8 CfAP1基因参与调控果生刺盘孢菌附着胞的形成及膨压
野生型CFLH16附着胞形成数量为突变体ΔCfap1-8附着胞形成量的2.6倍(图8A、8B),统计分析差异显著(P≤0.01)。附着胞主要依靠膨压产生的机械穿透力侵入植物角质层,因此推测ΔCfap1-8致病力减弱与附着胞膨压有关。采用2 mol·L-1浓度的甘油检测附着胞的膨压(图8C)。CfAP1基因敲除突变体ΔCfap1-8附着胞塌陷率较野生型和回补菌株高出37.3%,统计分析差异极显著(P≤0.01)。以上结果表明,CfAP1基因参与调控果生刺盘孢菌附着胞的形成及其内部膨压。
3 讨论
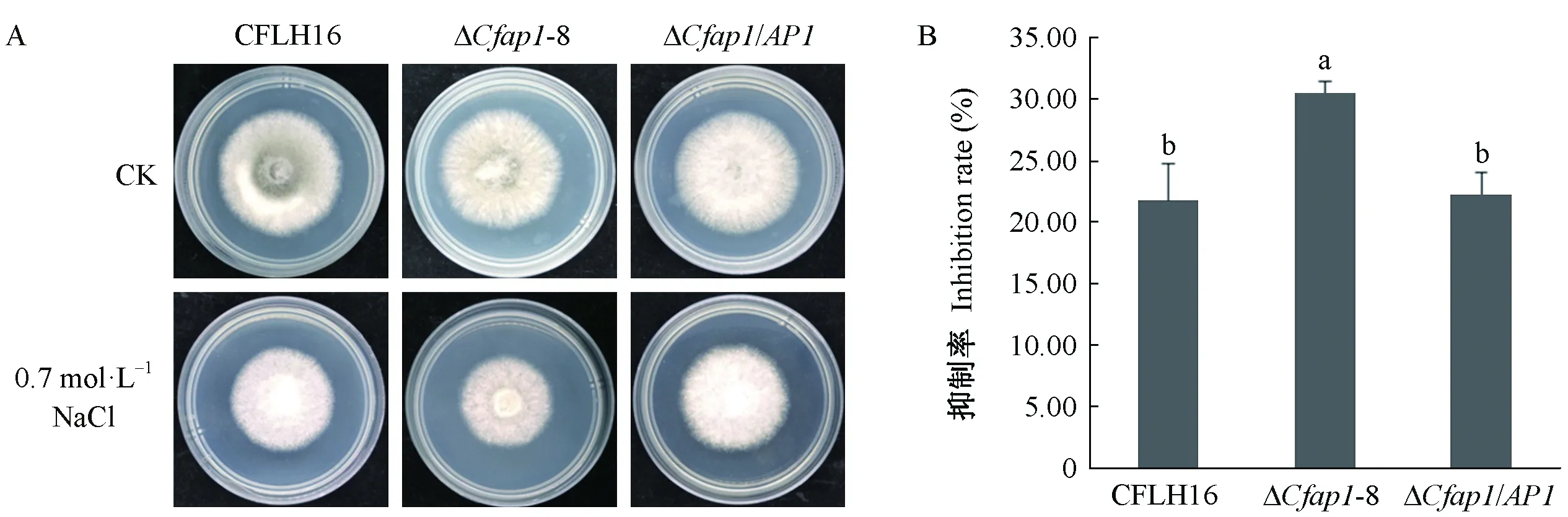
图6 CfAP1基因缺失突变体对渗透压胁迫的敏感性测定Fig.6 Osmotic sensitivity test of CfAP1 gene-deletion mutantsA: 野生型、ΔCfap1-8突变体和回补菌株接种在含有0.7 mol·L-1 NaCl的PDA培养基上的菌落生长情况。 B: 菌株在不同的渗透压胁迫培养基中生长抑制率统计分析. 误差线采用的是标准偏差; 小写字母不同者表示在0.01水平上差异极显著. A: The wild-type strain (CFLH16),ΔCfap1-8, and complemented strains were inoculated on PDA plates containing 0.7 mol·L-1 NaCl. B: Statistical analysis of the inhibition rate of the indicated strains on different stressors. Error bars are standard deviation and different lowercases represent significantly difference at P<0.01.
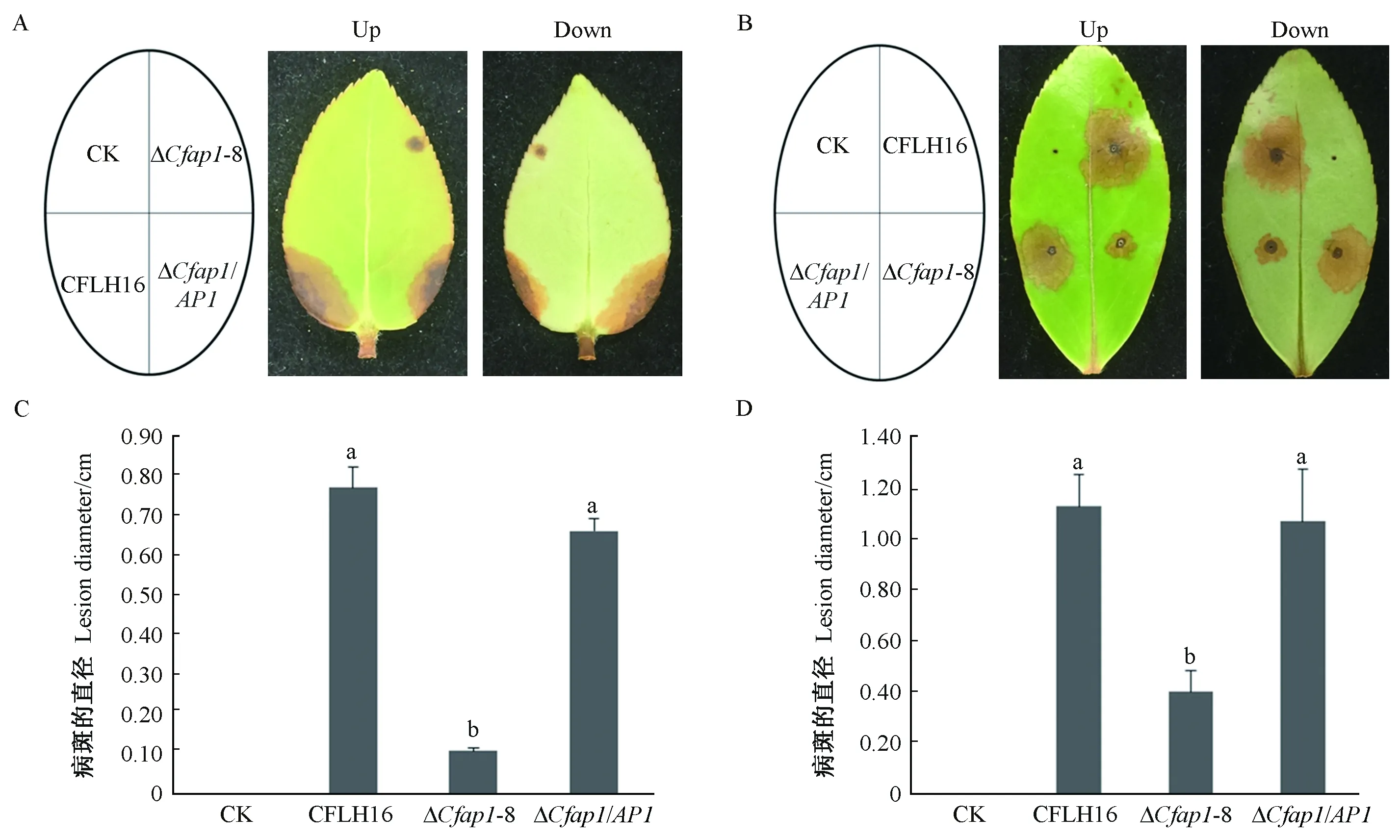
图7 CfAP1基因缺失突变体对油茶叶片的致病力Fig.7 Pathogenicity of CfAP1 gene deleted mutants to oil-tea leavesA、B: 野生型菌株、回补菌株和突变体的菌饼分别接种在无伤和有伤的油茶叶片上; C:无伤叶片病斑大小; D: 有伤叶片病斑大小。小写字母不同者表示在0.01水平上差异极显著。 A and B: the wild type strain CFLH16, CfAP1 deletion mutant and the complemented strains were inoculated unwounded leaves and on wounded leaves; C: Statistical analysis of the disease lesion size of wounded leaves; D: Statistical analysis of the disease lesion size of unwounded leaves. Error bars are standard deviation and different lowercases represent significantly difference at P≤0.01.
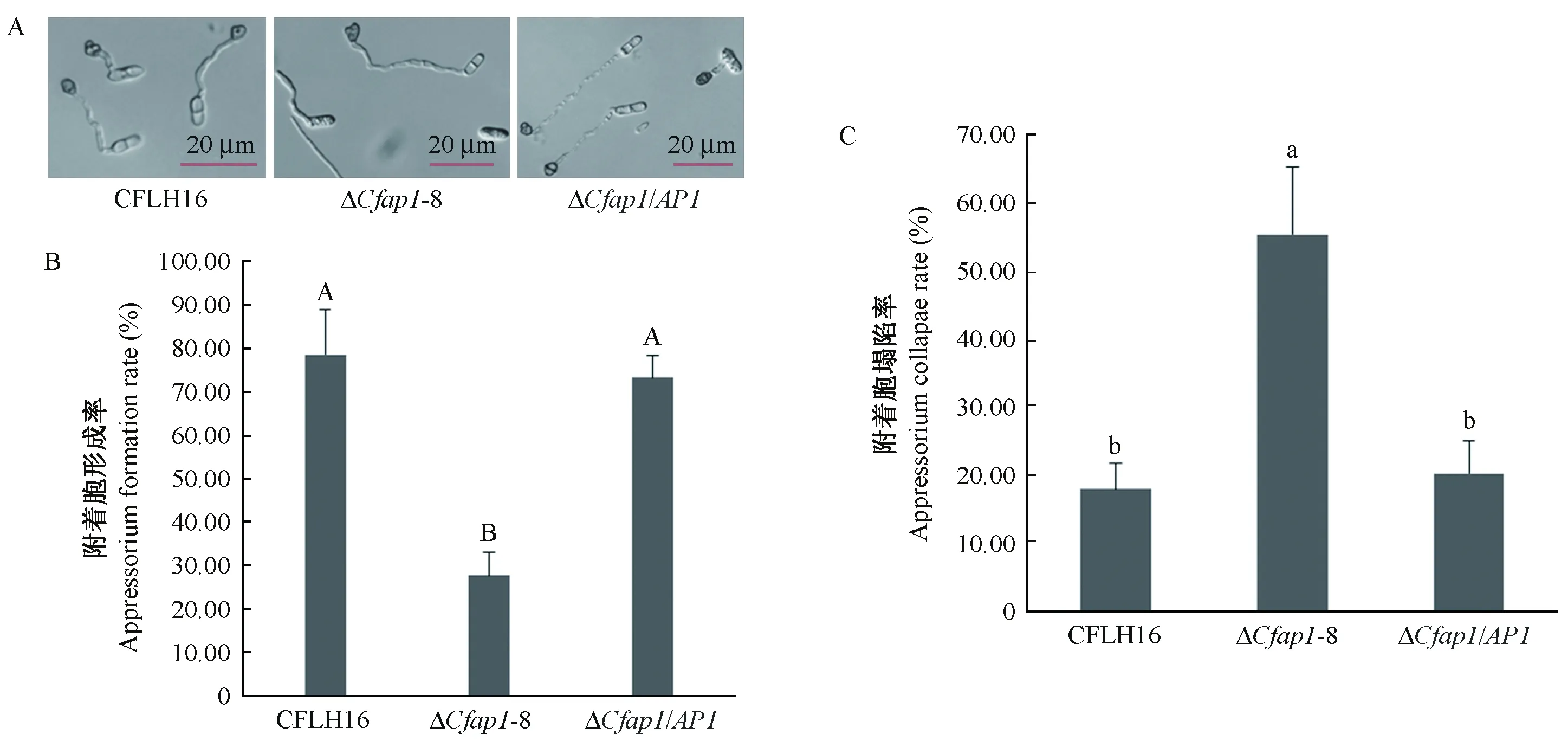
图8 CfAP1基因缺失突变体附着胞Fig.8 Appressorium in CfAP1 gene deleted mutantsA: 突变体ΔCfap1-8能形成少量附着胞; B: 附着胞形成率; C: 附着胞塌陷率。误差线采用的是标准差;大写字母不同者表示在0.05水平上差异显著,小写字母不同者表示在0.01水平上差异极显著。A:The CfAP1 gene deletion mutant can form a small amount appressorium; B: Statistical analysis of the rate of appressorium formation on glass surface; C:Appressorium collapse rate. Error bars are standard deviation and different capitals represent significantly difference at P≤0.05, different lowercases represent significantly difference at P≤0.01.
果生刺盘孢是油茶炭疽病的优势流行致病菌。本研究在果生刺盘孢菌中鉴定到了1个具bZIP结构域的转录因子CfAp1,并对该转录因子的生物学功能进行了研究。转录因子CfAp1参与调控油茶果生刺盘孢的生长发育、产孢、响应环境胁迫和致病力,本研究结果为开发果生刺盘孢的新型杀菌剂提供了潜在靶标。笔者实验室前期对油茶果生刺盘孢部分基因的功能进行研究,结果表明CfPMK1、CfHAC1和CfSNF1等基因参与调控油茶果生刺盘孢生长发育、胁迫应答和致病力(李河等2018;姚权等2019;Zhangetal., 2019),这与转录因子CfAp1的功能相似,说明参与调控果生刺盘孢致病的基因网络十分复杂,可能存在多种基因的协同作用,需要深入研究致病基因及相关网络调控致病的分子机制,为制定油茶炭疽病防治策略提供理论依据。
病原菌在侵染植物的过程中会受到各种环境因子的胁迫。本研究发现,果生刺盘孢菌CfAP1基因缺失突变体ΔCfap1-8对H2O2十分敏感,说明转录因子CfAp1参与应答H2O2胁迫,这与Guo(2011)对稻瘟病菌MoAp1基因的研究结果一致。另外,在渗透压胁迫试验中发现,ΔCfap1-8对0.7 mol·L-1的NaCl耐受性较野生型和回补菌株显著性降低,表明果生刺盘孢菌的CfAP1基因参与外源NaCl胁迫的应答。李河(2018)等的研究发现果生刺盘孢的CfPMK1基因也参与NaCl胁迫应答,这表明在该菌中不止一种基因参与渗透压胁迫应答。
致病力是病原菌所具有损害寄主植物和引起病变的能力。本研究发现,果生刺盘孢菌CfAP1基因缺失突变体ΔCfap1-8致病力显著降低,说明CfAp1转录因子参与调控果生刺盘孢菌的致病力。但在稻瘟病菌中,基因MoAP1缺失会导致稻瘟菌丧失致病性(Guoetal., 2011),表明CfAp1转录因子和MoAp1转录因子都参与调控致病力,但对病菌的致病力的调控程度不同,AP1基因对稻瘟病菌的致病力影响更大。
分生孢子在病原菌繁殖和致病过程中起着重要作用。果生刺盘孢主要通过分生孢子进行传播侵染,利用附着胞内部的黑色素层产生的巨大膨压推动侵染钉刺穿寄主表皮细胞致使寄主植物感病(Weiretal., 2012)。本研究发现,果生刺盘孢菌CfAP1基因敲除突变体ΔCfap1-8菌株分生孢子形成能力显著降低,说明CfAp1转录因子参与调控分生孢子的形成。李河(2018)研究发现,CfPMK1基因也参与油茶果生刺盘孢菌分生孢子的形成,这表明在该菌中多种基因参与调控其分生孢子的形成。Guo等(2011)的研究发现,稻瘟病菌中MoAP1基因缺失会导致稻瘟病菌的分生孢子明显变长、变细,而本研究中转录因子CfAp1参与调控产孢率但并不影响分生孢子的形态,这表明AP1基因在不同的物种中功能并不完全相同。
分生孢子产生的附着胞是病原菌侵染植物的重要结构。在适宜的环境条件下,分生孢子会萌发,芽管顶端膨大形成附着胞,附着胞分泌黏状物,黏附于寄主植物表面,形成纤细针状感染菌丝侵入寄主角质层吸取营养。张俊祥(2019)研究发现,CgCMK1基因缺失导致苹果炭疽叶枯病菌(C.gloeosporioides)不能形成附着胞,从而导致了苹果炭疽叶枯病菌完全丧失了致病性。李河(2018)研究发现,CfPMK1基因缺失导致油茶果生炭疽病菌完全不能形成附着胞,同样完全丧失了致病力。在本研究中,CfAP1基因突变体的附着胞形成率减少了51%,进一步对附着胞膨压测定发现,CfAP1基因敲除突变体附着胞膨压显著减小,这可能是其致病力下降的重要原因。然而,Sun等(2016)对胶孢刺盘孢菌CgAP1基因研究发现,该基因既不影响胶孢刺盘孢附着胞的产生,也不影响附着胞的膨压。这是否由于CfAP1基因在不同病原种类中的生物学功能也存在差异,还需要进一步研究。研究果生刺盘孢转录因子CfAp1调控的靶基因以及它们的生物学功能,揭示CfAp1转录调控的分子机制,为油茶炭疽病的防治提供依据。
4 结论
果生刺盘孢是油茶炭疽病的优势流行致病菌,本研究发现bZIP转录因子CfAp1参与调控油茶果生刺盘孢的生长发育、产孢、响应环境胁迫和致病力,该结果为开发果生刺盘孢的新型杀菌剂提供了潜在靶标。
