绿眼赛茧蜂线粒体基因组全序列测定和分析
2020-10-15王丹阳王予彤于良斌韩海斌徐林波崔艳伟康爱国庞红岩
王丹阳, 王予彤,2, 于良斌, 韩海斌, 徐林波,*, 崔艳伟,康爱国, 庞红岩
(1.中国农业科学院草原研究所, 呼和浩特 010020; 2.内蒙古农业大学, 呼和浩特 010020;3.河北省张家口市康保县植保站, 河北张家口 076650)
线粒体(mitochondrion)是真核生物细胞中一种重要的胞质细胞器,在细胞代谢、凋亡、疾病和衰老中起着重要作用(Boore, 1999; Alcolado, 2005; Cameron, 2014)。线粒体基因具有结构简单、组成稳定、排列保守、母系统遗传、进化速率快等特点,被广泛应用于系统发育分析、种群遗传分化等研究中(Boore, 1999; Ballard and Rand, 2005; Weietal., 2010a; 李倩, 2014)。对于大多数昆虫而言,线粒体基因组通常是一个大小为15~20 kb的双链闭合环状分子,编码37个基因,包括13个蛋白编码基因(PCGs)2个核糖体RNA(rRNA)基因和22个转运RNA(tRNA)基因,以及1个长的非编码区(又称为控制区或AT富含区)(Wolstenholme and Clary, 1985; Boore, 1999; Sharkey and Chapman, 2017)。
膜翅目(Hymenoptera)是六足总纲中较为高等的类群,除了具有上述普遍特征外,其线粒体基因组还有重排率较高,基因重排程度在类群间有差异,广腰亚目(Symphyta)排列较细腰亚目(Apocrita)保守等特点(魏书军, 2009; Weietal., 2014)。茧蜂科(Braconidae)是膜翅目最大的科,已记录了1 040多个属,19 000余种(Wharton and van Achterberg, 2000; Weietal., 2010b; Lietal., 2016),其中有许多种类是农林害虫的重要寄生性天敌。茧蜂科是研究寄生虫模式演变的理想群体,而该家族报道完整的线粒体基因组较少(李倩, 2014; 宋胜楠, 2015)。而且,茧蜂科昆虫几乎全部营寄生生活(Dangerfield and Austin, 1998),且寄主范围极其广泛,是重要的天敌资源昆虫,对其深入、全面的研究才能更好地发挥其利用价值。
绿眼赛茧蜂Zelechlorophthalmus属膜翅目(Hymenoptera)茧蜂科(Braconidae)优茧蜂亚科(Euphorinae),主要分布于我国河北、内蒙古、辽宁、吉林、黑龙江、浙江、安徽、甘肃、宁夏、新疆等地(何俊华等, 2004)。绿眼赛茧蜂为内寄生,是草地螟Loxostegesticticalis、甜菜夜蛾Spodopteraexigua和斜纹夜蛾Spodopteralitura等农业重大害虫的主要天敌(李倩等, 2017)。目前国内仅有少量关于其形态学、生物学特性等方面的研究,尚无对绿眼赛茧蜂线粒体基因组全序列的研究报道(杜芹, 2015; 李倩等, 2017)。虽然茧蜂科线粒体基因组的信息已有报道,但是并不全面,而且茧蜂科昆虫几乎全部营寄生生活(Dangerfield and Austin, 1998),且寄主范围极其广泛,是重要的天敌资源昆虫,对其深入、全面的研究才能更好地发挥其利用价值。本研究对绿眼赛茧蜂线粒体全基因组进行了测定,结合GenBank中已收录的21种茧蜂科昆虫的线粒体基因组COX1序列进行比较分析,探讨茧蜂科各亚科、各属及种间的系统发育关系,旨在为绿眼赛茧蜂的种群遗传学、分子生态学以及茧蜂科的分子系统发育研究奠定基础。
1 材料与方法
1.1 供试昆虫
绿眼赛茧蜂成虫于2018年5月采自中国农业科学院草原研究所沙尔沁农牧交错区试验基地,室内以草地螟为寄主,在温度21~23℃、相对湿度50%~70%、光周期16L∶8D条件下,以15%蜂蜜水继代饲养繁育。
1.2 绿眼赛茧蜂总DNA的提取
取单头成虫用于总DNA提取,采用DNeasy DNA Extraction Kit试剂盒(QIAGEN),参照说明书进行总DNA的抽提,提取后采用Thermo Scientific NanoDrop 2000检测纯度及浓度,采用琼脂糖电泳和Agilent 2100 Bioanalyzer检测其完整性。
1.3 序列测定
采用全基因组鸟枪法(whole genome shotgun, WGS)策略构建文库,利用第二代测序技术(next generation sequencing, NGS),基于Illumina MiSeq测序系统,对构建的文库进行双末端(paired-end, PE)测序。
1.4 基因组序列拼装与注释
采用A5-miseq v20150522(Coiletal., 2015)和SPAdesv3.9.0(Bankevichetal., 2012)对高质量的二代测序数据进行从头拼装,构建contig和scaffold序列。根据拼接序列的测序深度提取序列,将高测序深度的序列同NCBI上的nt库进行blastn(BLAST v2.2.31+)比对,挑出各拼接结果的线粒体序列。拼接结果整合:将以上不同软件得到的线粒体拼接结果利用mummer v3.1软件进行共线性分析,确定contig间的位置关系,进行contigs间gap的填补。使用pilon v1.18(Walkeretal., 2014)软件对结果进行校正以得到最终的线粒体序列。使用MEGA7.0对线粒体基因组各部分的碱基组成和相对同义密码子使用频率(relative synonymous codon usage, RSCU)进行统计。
将拼接得到的完整的线粒体基因组序列上传至MITOS网页服务器(http:∥mitos.bioinf.uni-leipzig.de)进行功能注释(Berntetal., 2013)。其中Genetic Code选择设置为05-inverterbrate,其余设置按照MITOS设置的默认参数,分别计算线粒体全基因组、蛋白编码基因、基因排列顺序、rRNA基因的碱基组成等。采用OGDRAM(ogdraw.mpimp-golm.mpg.del)在线可视化软件绘制线粒体全基因组圈图(Lohseetal., 2013)。
1.5 系统发育分析
以蜜蜂科的两个种小蜜蜂Apisflorea和大蜜蜂Apisdorsata为外群,选用已报道的21种茧蜂科昆虫和绿眼赛茧蜂的线粒体基因组COXⅠ基因的核苷酸序列,通过MEGAX的Align by Clustal-X程序进行序列比对(Kumaretal., 2016),利用MEGA-X软件采用最大似然法(ML)和邻接法(NJ)构建茧蜂科的系统发育树。系统树分支的置信水平用bootstrap test(Felsenstein, 1985)估计,重复抽样1 000次。
2 结果
2.1 绿眼赛茧蜂线粒体基因组的基因组成
绿眼赛茧蜂线粒体基因组全长16 661 bp(GenBank登录号: MG822749),基因组的基因排布与已报道茧蜂科其他种类的(Weietal., 2014; Lietal., 2016)相似度高,呈双链闭合环状结构(图1)。基因组成包含13个蛋白质编码基因(PCGs)、22个tRNA基因、2个rRNA基因(rrnS和rrnL)和1个AT控制区(CR)。13个蛋白质编码基因中有4个(NAD1,NAD4l,NAD4,NAD5)在N链上,其余9个(Cob,NAD6,NAD3,COX3,ATP6,ATP8,COX2,COX1,NAD2)在J链上;22个tRNA基因中有10个位于N链,12个位于J链上;2个rRNA基因均位于N链上,其中rrnS基因长748 bp,位于tRNAIle(I)和tRNAVal(V)之间;rrnL基因长1 264 bp,位于tRNAVal(V)和tRNALeu-1(L1)之间(图1);控制区长度418 bp,位于tRNAGln(Q)和tRNAMet(M)之间。
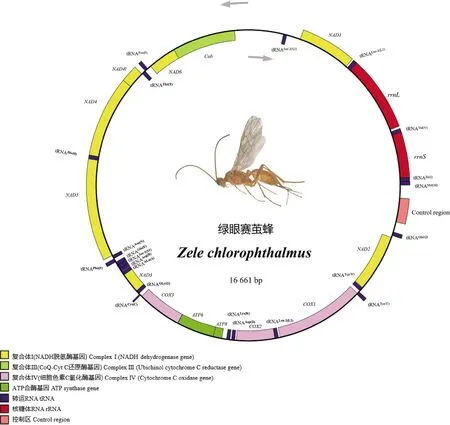
图1 绿眼赛茧蜂线粒体基因组结构
绿眼赛茧蜂线粒体基因组上的基因排列顺序与昆虫线粒体原始排列(魏书军, 2009; 魏书军和陈学新, 2011)存在一定差异。13个蛋白质编码基因和2个rRNA基因的排列顺序和方向均相同,7个tRNA基因发生重排。tRNACys(C)从tRNATrp(W)和tRNATyr(Y)之间移到了COX3和tRNAGly(G)之间,tRNAAsn(N)从tRNASer-1(S1)和tRNAArg(R)之间移到了tRNAPhe(F)和NAD5之间,tRNALys(K)和tRNAAsp(D)以及tRNASer-1(S1)和tRNASer-2(S2)发生了基因位置互换,tRNAIle(I)发生了倒置,tRNAMet(M)和tRNAGln(Q)发生了异位倒置。
绿眼赛茧蜂线粒体基因组有16处基因重叠,共107 bp,最长重叠位于COX1和tRNATyr(Y)之间,重叠序列长24 bp。基因间隔区域有18处,共1 886 bp,最长间隔854 bp,位于tRNASer-2(S2)和Cob之间;其次为NAD1和tRNASer-2(S2)之间,间隔236 bp。既无重叠又无间隔有4处(表1)。
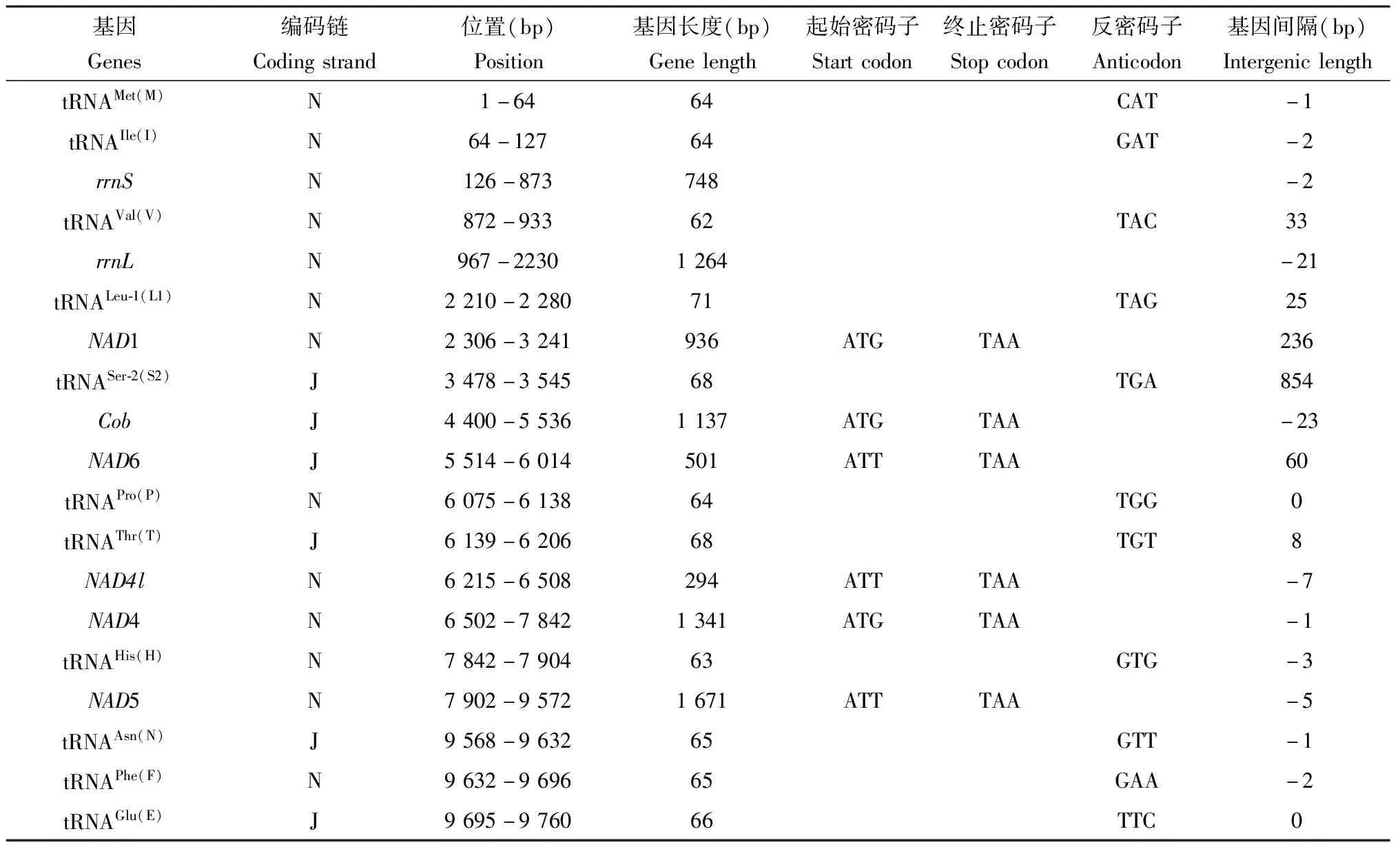
表1 绿眼赛茧蜂线粒体基因组特征分析
2.2 绿眼赛茧蜂线粒体基因组核苷酸组成
绿眼赛茧蜂线粒体基因组中,A, T, G和C含量分别为43.00%, 39.84%, 10.09%和7.08%,A含量最高而C含量最低(表2)。全基因组的A+T总量高达82.83%,G+C含量为17.17%,呈现明显的A+T偏好性,符合昆虫线粒体基因组中A+T含量偏向性的特征(夏靖等, 2011; 王菊平等, 2015; 钟健等, 2017)。绿眼赛茧蜂基因组全序列的AT偏斜(AT-skew)为0.038,GC偏斜(GC-skew)为0.176,表明A多于T,G多于C。从不同的基因来看,蛋白质编码基因的AT偏斜均为负值,J链上9个蛋白质编码基因的GC偏斜为负值。N链上4个蛋白质编码基因和rRNA基因的GC偏斜均为正值,与大多数昆虫的线粒体基因组AT/GC偏斜(彭艳等, 2017)一致。
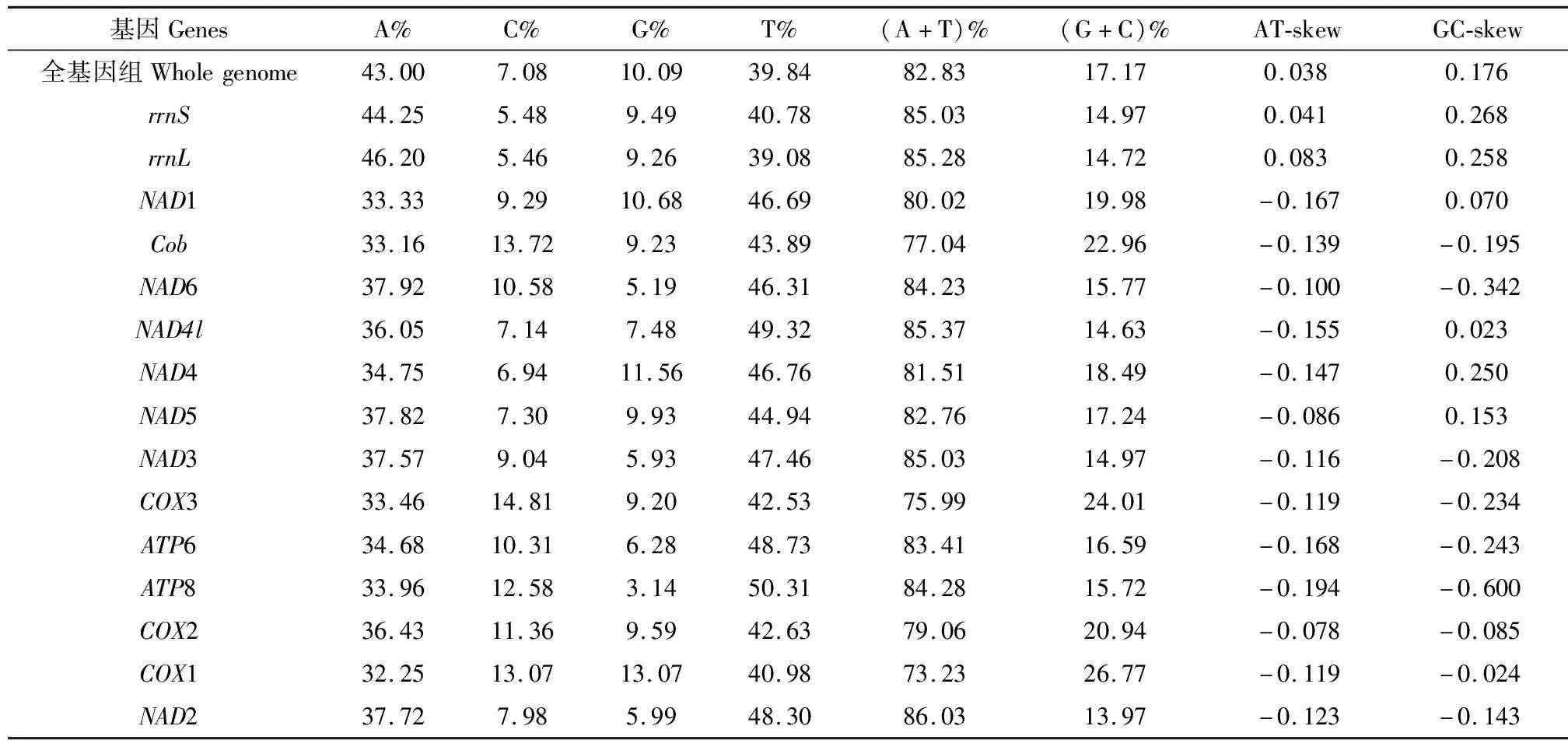
表2 绿眼赛茧蜂线粒体基因组的核苷酸组成
2.3 绿眼赛茧蜂线粒体基因组蛋白质编码基因(PCGs)
绿眼赛茧蜂线粒体蛋白质编码基因长度与其他膜翅目昆虫线粒体蛋白编码基因相似,13个蛋白质编码基因均以ATN为起始密码子,以TAA为终止密码子,其中NAD3以ATA为起始密码子,NAD6,NAD4l,NAD5,ATP8,COX1和NAD2以ATT为起始密码子,其余6种均以ATG作为起始密码子。绿眼赛茧蜂线粒体蛋白质编码基因的相对同义密码子使用频率(RSCU)见表3。绿眼赛茧蜂线粒体蛋白质编码基因对A和T有明显的偏向性,使用最多的5个密码子是AAA(Lys)525次、UAA(终止密码子)465次、UUA(Leu)459次、AAU(Asn)420次和UUU(Phe)394次。只有Met和Trp仅有一种密码子,分别为AUG和UGG。除了Met和Trp外,绿眼赛茧蜂蛋白质编码基因使用频率最高密码子第3位偏好使用A和U,与一些膜翅目昆虫(Fanetal., 2017; 彭艳等, 2017)相似,蛋白质编码基因的氨基酸百分比最高者为Ile(15.3%),其次为Leu(13.2%), Lys(11.2%)和Phe(8.6%)。
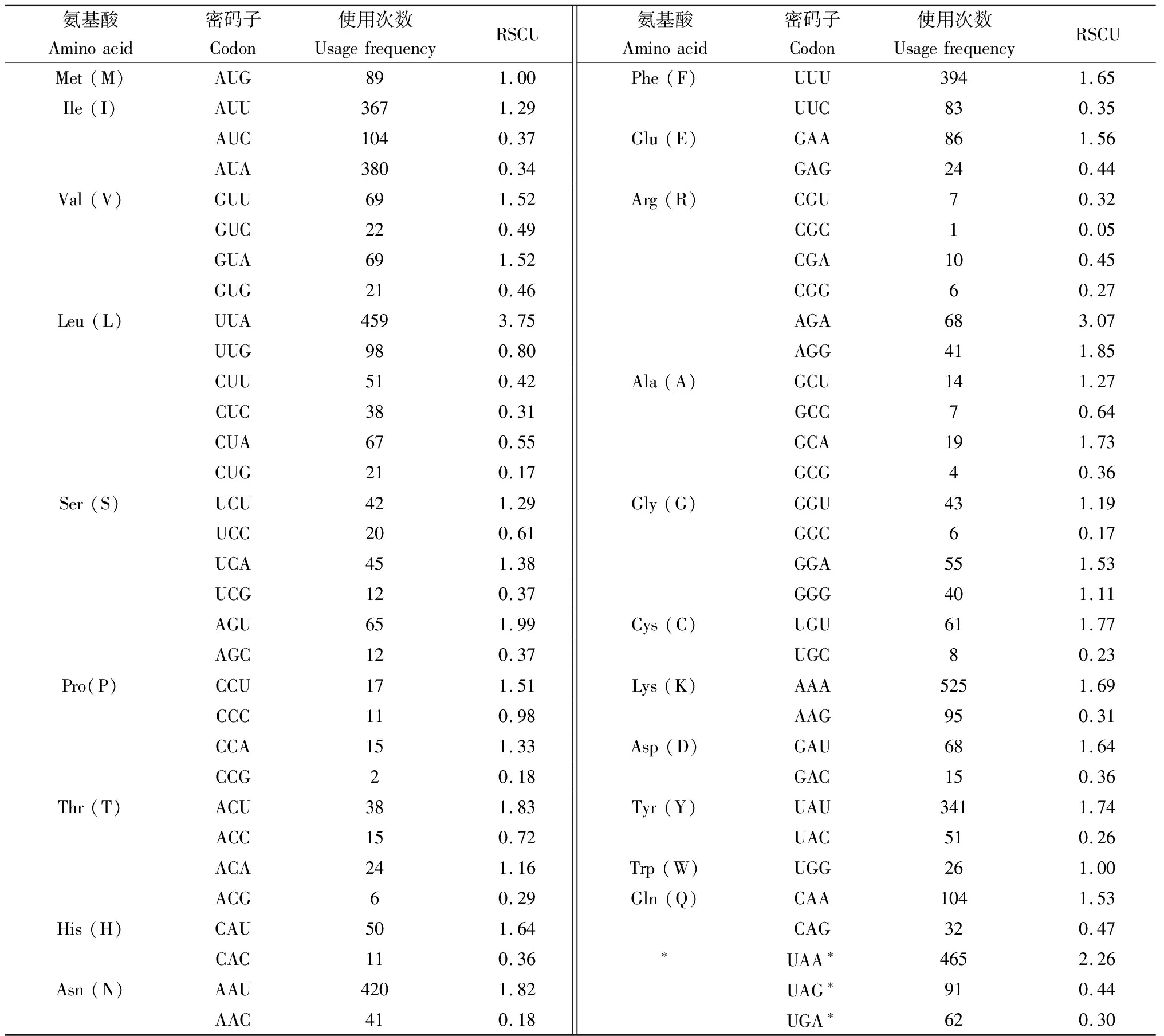
表3 绿眼赛茧蜂线粒体基因组相对同义密码子使用频率(RSCU)
2.4 绿眼赛茧蜂线粒体基因组tRNA基因和rRNA基因
绿眼赛茧蜂线粒体基因组的22个tRNA基因与其他膜翅目昆虫线粒体基因组对应的tRNA反密码子相同,除tRNALeu和tRNASer有两个tRNA结构外,其他都只有一个与之对应。除tRNAHis(H)缺失TΨC环和tRNACys(C)仅剩DHU臂和反密码子臂外,其他tRNA基因都能形成典型的三叶草式二级结构(图2)。在三叶草结构中还发现有6处错配:tRNAAla(A)有1处,错配于氨基酸臂上;tRNAPhe(F), tRNAGly(G)和tRNAVal(V)各有1处,位于DHU臂上;tRNAGln(Q)有2处,分别位于DHU臂和反密码子臂上,发生错配的碱基均为G-U。

图2 绿眼赛茧蜂线粒体基因组tRNA基因二级结构
绿眼赛茧蜂线粒体基因组的2个rRNA基因与大部分昆虫线粒体rRNA基因相比类似,具有明显的AT碱基偏向性,rrnS与rrnL的长度分别为748 bp和1 264 bp(表1),A+T的含量分别为85.03%和85.28%(表2)。
2.5 系统发育分析
以蜜蜂科为外群,基于已报道的21个茧蜂科种和绿眼赛茧蜂的线粒体基因组COX1蛋白编码基因序列,采用最大似然法(ML)和邻接法(NJ)利用MEGA-X软件构建茧蜂科的系统发育树(图3和4)。最大似然法分析结果(图3)表明,茧蜂科各个种群分类关系为蚜茧峰亚科Aphidiinae+(长体茧蜂亚科Macrocentrinae+毛茧蜂亚科Doryctinae+探茧峰亚科Ichneutinae+(窄径茧蜂亚科Agathidinae+折脉茧蜂亚科Cardiochilinae))+优茧峰亚科Euphorinae;邻接法分析结果表明,优茧峰亚科Euphorinae+(蚜茧峰亚科Aphidiinae+((窄径茧蜂亚科Agathidinae+折脉茧蜂亚科Cardiochilinae)+(探茧峰亚科Ichneutinae)+(长体茧蜂亚科Macrocentrinae+毛茧蜂亚科Doryctinae)))。综合比较,两种方法构建的系统发育树大致相同。在系统发育树中,茧蜂科主要分为3支,蚜茧峰亚科为一支,优茧蜂亚科为一支,剩余的一支包括长体茧蜂亚科、毛茧蜂亚科、探茧蜂亚科、折脉茧蜂亚科和窄径茧蜂亚科。窄径茧蜂亚科(Agathidinae)和折脉茧蜂亚科(Cardiochilinae)在两种方法中都为姐妹群。绿眼赛茧蜂聚于优茧蜂亚科赛茧蜂属,与传统形态分类学结果一致,其与雪跗赛茧蜂Zeleniveitarsis亲缘关系较近。
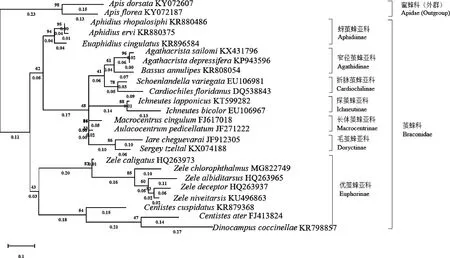
图3 最大似然法构建的基于线粒体COX1序列的茧蜂科22个种的系统发育树(1 000次重复)
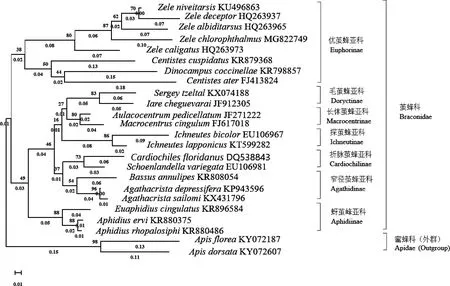
图4 邻接法构建的基于线粒体COX1序列的茧蜂科22个种的系统发育树(1 000次重复)
3 讨论
亚库巴果蝇Drosophilayakuba线粒体基因组的排列方式被认为是昆虫线粒体基因组最原始的排列方式(魏书军和陈学新, 2011),绿眼赛茧蜂线粒体基因组与其相比,存在基因重排现象。本研究通过与茧蜂科其他种的线粒体基因组比较分析,发现绿眼赛茧蜂线粒体基因组重排多出现于tRNA基因中,除tRNALeu-2(L2)处于COX1和COX2之间较稳定,没有出现重排外,其余tRNA基因都曾出现重排现象。膜翅目分为广腰亚目(Symphyta)和细腰亚目(Apocrita)两个类群,广腰亚目的基因排列相比细腰亚目保守,重排较少(宋胜楠, 2015),但细腰亚目存在大量重排,包括移位、原位倒置、异位倒置(Weietal., 2010b),而绿眼赛茧蜂属于细腰亚目。膜翅目线粒体具有较高的重排速率,主要集中发生于AT富含区NAD2,NAD2-COX1,COX2-ATP8和NAD3-NAD5区(魏书军, 2009)。
膜翅目茧蜂科昆虫线粒体基因组中有大量的基因间隔区和基因重叠区的现象并不少见。离颚细蜂科的Vanhorniaeucnemidaru存在36处基因间隔,5处基因重叠,基因间隔总长达1 777 bp,基因重叠共31 bp(Castroetal, 2006);盖拉头甲肿腿蜂Cephalonomiagallicola基因间隔总长度比绿眼赛茧蜂基因间隔长,共有22处,长2 543 bp,基因重叠4处,共36 bp(魏书军, 2009);半闭弯尾姬蜂Diadegmasemiclausum也有13处基因间隔,共1 846 bp,14处基因重叠,共61 bp(魏书军, 2009)。比较发现,基因重排发生的区域基因间隔较大,而基因重叠区基因重排发生较少。茧蜂科昆虫常有一些较短的基因间隔区,但也存在例外,绿眼赛茧蜂有两处较长的基因间隔,NAD1与tRNASer-2(S2)之间有236 bp的基因间隔,tRNASer-2(S2)与Cob之间有854 bp的基因间隔,推断是由于tRNASer-2(S2)和tRNASer-1(S1)发生了位置互换产生的。发生重排的基因左右都有较大的基因间隔区,因此推断基因间隔可能是昆虫线粒体基因组产生重排的原因,具体原因还需要更进一步的研究。
通过不同方法构建系统发育树,综合分析发现优茧蜂亚科和蚜茧峰亚科的单系性得到很高的支持。窄径茧蜂亚科和折脉茧蜂亚科互为姐妹群。宽鞘茧蜂属Centistes、瓢虫茧蜂属Dinocampus和赛茧蜂属Zele同属于优茧蜂亚科(Euphorinae),这与传统分类方法的结果(陈学新等, 2000)一致,但宽鞘茧蜂的两个种并没有聚到一起,还需要进一步研究。两种方法都将窄径茧蜂亚科、折脉茧蜂亚科、探茧蜂亚科、长体茧蜂亚科和毛茧蜂亚科聚于一支,说明这5种亚科之间的亲缘关系较近,而与优茧蜂亚科和蚜茧峰亚科关系较远。
本研究对绿眼赛茧蜂线粒体基因组全序列的测定丰富了茧蜂科线粒体基因组的序列信息,为分子进化信息系统增添了有益的数据信息,并且为线粒体基因的结构和组成提供了基础数据,为后续茧蜂科系统发生关系奠定基础。
