不同浓度赤霉素处理对辽东楤木种子萌发的影响
2020-10-09陈映彤于锡宏白国梁张雪刘汉兵赵恒田蒋欣梅
陈映彤,于锡宏,2#,白国梁,张雪,刘汉兵,赵恒田,蒋欣梅,2*
(1.农业农村部东北地区园艺作物生物学与种质创制重点试验室,东北农业大学,哈尔滨 150030;2.林下经济资源研发与利用协同创新中心,哈尔滨 150040;3.中国科学院东北地理与农业生态研究所,哈尔滨 150040)
辽东楤木(Araliaelata)也称刺嫩芽、刺老芽,是五加科楤木属植物,主要分布在我国东北部地区。辽东楤木是一种重要的药食同源植物,食用的嫩芽中含有多种氨基酸、维生素及大量的微量元素,能够为人体提供丰富的营养物质,同时具有很好的保健作用,具有活血化瘀、抗癌等功效,被誉为“山野菜之王”[1]。随着人们对这种药食同源类植物的需求升高,很多野生资源遭到破坏,大面积的辽东楤木资源消失,人工栽培成为解决辽东楤木资源匮乏的主要途径。然而,很多野生资源常常因种胚发育不完全,具有很强的休眠特性[2],辽东楤木也如此,人工栽培利用种子繁殖的过程中,解除种子休眠是首要的问题。打破种子休眠有很多种方法,包括低温层积处理、外源激素处理、光照、超声波及机械处理等[3]。生产中常常在上冻前采用低温层积处理——即沙藏方式,第二年早春取出后进行播种育苗;采用变温层积尽管可以有效提高辽东楤木种胚的后熟,但依然存在发芽率较低且发芽持续时间长的问题[4]。赤霉素作为一种生长素,是植物生长中最重要的物质之一,赤霉素含量降低往往意味着植株体的休眠甚至死亡,特别是对于种子来说,赤霉素含量的高低影响着种子内进行物质运输的速度,并且影响胚的分化和生长,适宜浓度的赤霉素对种子的萌发具有促进作用[5]。关于外源施用赤霉素对辽东楤木种子萌发方面的研究鲜有报道,为此,本试验通过研究不同浓度外源赤霉素处理变温层积后的辽东楤木种子,拟明确种子萌发过程中内含物质的变化规律,旨在为探究辽东楤木种子萌发机理提供理论依据,同时为辽东楤木种苗生产选用适宜的赤霉素浓度提供参考。
1 材料与方法
1.1 试验材料
本材料选用当年饱满成熟的辽东楤木种子,来自于农业农村部东北地区园艺作物生物学与种质创制重点实验室。
1.2 试验方法
将采集的辽东楤木种子洗净,用水浮法去除瘪粒。浸种48 h后选取籽粒饱满种子进行变温层积处理(沙藏层积用的细河沙经高温灭菌),即将层积的辽东楤木种子放在(12±1)℃下恒温箱中处理90 d、(4±1)℃下处理60 d。将经变温层积处理后的种子采用不同浓度外源赤霉素(GA3)浸种12 h后放在20 ℃条件下催芽。外源赤霉素设置5个浓度水平,分别为100、150、200、250、300 mg/kg(代码分别为T100、T150、T200、T250、T300),以清水作为对照(CK)。
分别在催芽后的第10 d、30 d、50 d时,对种子中关键内含物质含量进行测定,其中可溶性糖含量测定采用蒽酮比色法[6]、可溶性蛋白质含量测定采用 G-250 法[6]、过氧化物酶(POD)活性测定采用愈创木酚法[7]、过氧化氢酶(CAT)活性测定采用高锰酸钾滴定法[7]、超氧化物歧化酶(SOD)活性测定采用 NBT 法[7]、内源激素(赤霉素、脱落酸)含量测定采用高效液相色谱法(HPLc)测定[6],同时随机选取100粒种子测定发芽率。3次重复。
采用Excel 2010和SPSS 21.0统计软件对试验数据进行计算和统计分析。
2 结果与分析
2.1 不同浓度赤霉素处理对辽东楤木种子发芽率的影响
利用赤霉素处理变温层积处理后的辽东楤木种子可以显著提高其发芽率,赤霉素一定浓度范围内,发芽率随着浓度的增加而增加。催芽后10 d开始陆续发芽;催芽后30 d开始,不同浓度赤霉素处理的辽东楤木种子发芽率显著提高,其中催芽后30 d时的发芽率尽管在25.67 %~27.99%,但赤霉素浓度在200~300 mg/kg的3个处理(T200、T250、T300)与T150之间均差异不显著,但显著高于T100处理;随着催芽时间延长,在催芽后50 d时各个处理发芽率均达到了50%以上,此时发芽率随着赤霉素浓度的增加而显著增加,浓度达到200 mg/kg时(T200)发芽率最高,达到了66.22%,比对照CK提高了 18.74 %,而后再随赤霉素浓度增加而发芽率显著降低。

图1 不同浓度赤霉素处理对辽东楤木种子发芽率的影响不同字母表示P<0.05水平(n=3)差异显著。下同。
2.2 不同浓度赤霉素处理对辽东楤木种子可溶性糖和可溶性蛋白质含量的影响
由表1可知,随着辽东楤木种子的萌发,可溶性糖含量逐渐降低而可溶性蛋白含量逐渐增加。可溶性糖含量在催芽后前30 d变化不明显,进入50 d时,随赤霉素浓度的增加,可溶性糖含量表现出先下降再上升趋势; 可溶性蛋白质含量在催芽后前10 d各个处理与对照之间均差异不显著,催芽后30 d和50 d时,均随赤霉素浓度的增加表现出先升高后下降趋势;其中200 mg/kg赤霉素处理(T200)下的可溶性糖含量在催芽后50 d最低,与其他处理及对照之间差异显著;T200处理下的可溶性蛋白质含量在催芽后30 d和50 d均最高,且与其他处理及对照之间差异显著。
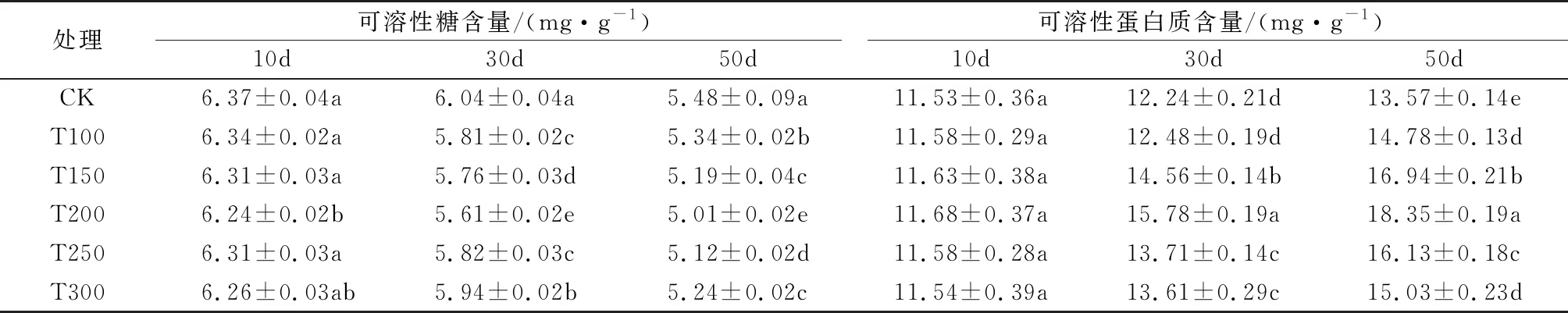
表1 不同浓度赤霉素处理对辽东楤木种子萌发过程中可溶性糖和可溶性蛋白含量的影响
2.3 不同浓度赤霉素处理对辽东楤木种子萌发过程中相关酶活性的影响
由图2、3、4可知,随着处理时间的增加,辽东楤木种子逐渐萌发过程中,过氧化物酶活性(POD)、过氧化氢酶活性(CAT)和超氧化物歧化酶活性(SOD)均逐渐升高。对于POD和SOD酶活性,赤霉素处理对辽东楤木催芽前30 d影响不大,均表现为各个处理以及对照之间均差异不显著,催芽后50 d时两种酶活性随着赤霉素浓度的增加表现出先升高后下降的趋势,其中T200处理处于高水平;对于CAT酶活性,赤霉素处理对辽东楤木催芽前10 d影响不大,催芽后30 d和50 d,随着赤霉素浓度的增加均表现出先升高后下降的趋势,其中T200处理处于较高水平。

图2 不同浓度赤霉素处理对辽东楤木种子萌发过程中过氧化物酶活性的影响
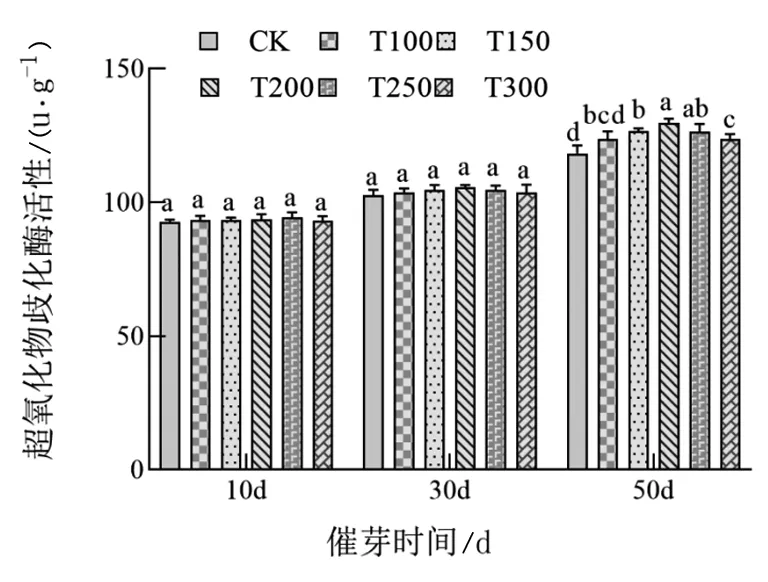
图3 不同浓度赤霉素处理对辽东楤木种子萌发过程中超氧化物歧化酶活性的影响
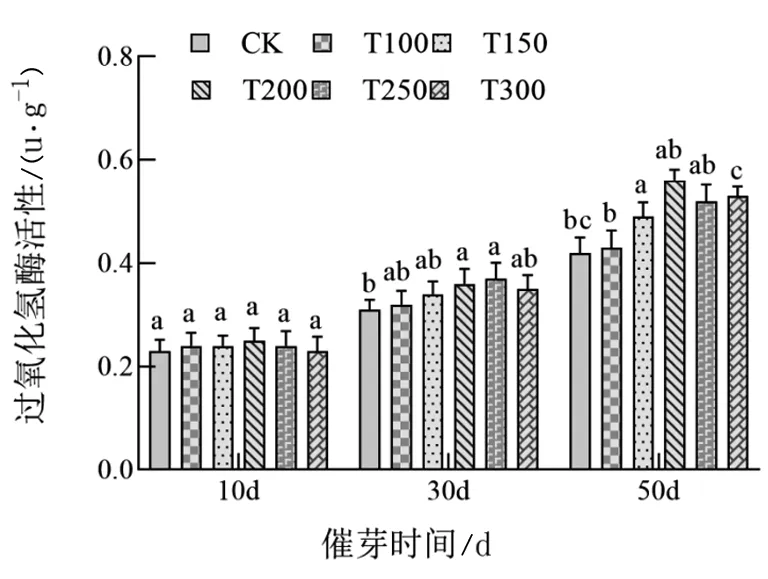
图4 不同浓度赤霉素处理对辽东楤木种子萌发过程中过氧化氢酶活性的影响
2.4 不同浓度赤霉素处理对辽东楤木种子萌发过程中内源激素含量的影响
由表2可知,随着辽东楤木种子的萌发,赤霉素含量逐渐升高,而脱落酸含量逐渐降低。赤霉素含量在催芽后10 d变化不明显,催芽后30 d和50 d时,随着外源赤霉素浓度的增加,均表现出先升高后降低的趋势;脱落酸含量在催芽后前10 d和30 d各处理及对照间均差异不显著,在催芽后50 d时,随着赤霉素浓度升高,脱落酸含量表现为先下降后上升的趋势;其中T200处理下赤霉素含量在催芽后30 d和50 d均最高,而脱落酸含量在催芽后50 d则最低,与对照之间均达到了显著水平。
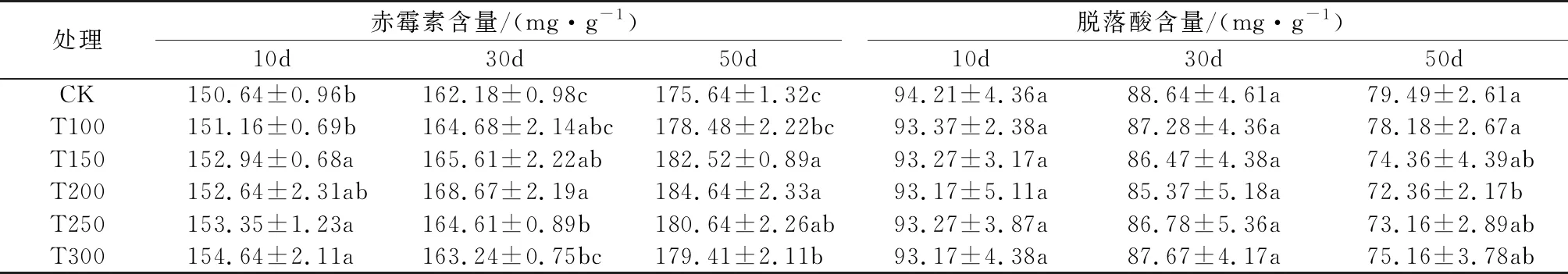
表2 不同浓度赤霉素处理对辽东楤木种子萌发过程中赤霉素含量和脱落酸含量的影响
2.5 辽东楤木种子发芽率与内含物变化相关性分析
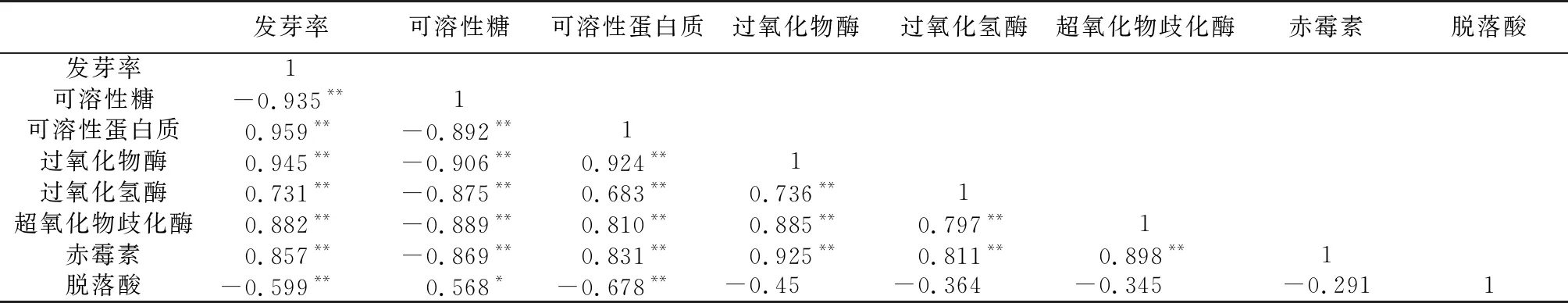
表3 辽东楤木种子萌发率与内含物变化相关性分析
对辽东楤木种子发芽率与内含物含量之间进行相关性分析表明(见表3):辽东楤木种子发芽率与可溶性糖、脱落酸呈极显著负相关关系,与可溶性蛋白质含量、赤霉素含量以及3种酶活性(过氧化物酶、过氧化氢酶、超氧化物歧化酶)呈极显著正相关关系。可溶性糖与可溶性蛋白质、赤霉素、3种酶活性均呈极显著负相关性,与脱落酸呈显著正相关关系。可溶性蛋白质与3种酶活性及赤霉素呈极显著正相关,而与脱落酸呈极显著负相关。3种酶之间以及3种酶与赤霉素之间均呈极显著正相关关系。
3 讨论与结论
种子休眠的类型多且复杂,大致为2种情况,一种是种子已经成熟,但由于外界条件不适宜而不得不暂停萌发;另一种是种子的生理后熟和形态后熟未完成发育或受内源激素物质抑制而无法萌发[8]。辽东楤木种子萌发困难往往是由于其种胚发育不完全引起的休眠造成的。本试验采用12 ℃下90 d结合4 ℃下60 d的变温层积处理促进辽东楤木种胚后熟,经显微观察层积后辽东楤木的种胚占有率达到46%,但从发芽持续时间来看,在催芽后50 d时经过赤霉素处理的种子最高发芽率的仅为66%,说明尽管选用的种子为经过水浮法选出的籽粒饱满的种子,但种子个体之间种胚发育的程度不同,虽然经过了150 d的层积处理,但依然会存在种胚尚未完全成熟的种子,采用一定浓度外源赤霉素浸种对种胚的发育有促进作用,由于赤霉素的有效性一般在2周左右[9],本试验中只在层积结束后对辽东楤木种子处理了1次赤霉素,这也是种子发芽时间周期长的原因之一。关于辽东楤木种子形态自然成熟过程中种胚发育情况以及赤霉素处理辽东楤木种子的使用次数还有待于进一步研究。
植物种子解除休眠的过程,是一系列内源激素参与调控的过程,赤霉素(GA3)和脱落酸(ABA)是2种联系紧密又极受关注的一对拮抗机制激素。其中赤霉素作为促进激素,在解除种子休眠方面有显著的作用,主要表现为软化种胚周围组织,促进种胚生长;同时赤霉素能够提高各类酶活性、促进种子内物质的分解与合成[10]。一定浓度的赤霉素能够有效的克服种子休眠,使种子迅速萌发[11]。赤霉素处理在不同程度上提高了天女木兰种子和节节麦种子的发芽率,在天女木兰种子萌发过程中,内源赤霉素含量不断积累,呈现上升的趋势;节节麦种子内源赤霉素含量明显增高[12-13]。种子休眠及解除与脱落酸水平的降低密切[14],随种子的萌发,脱落酸含量可自我调节而明显下降[15]。赤霉素和脱落酸在种子内的平衡决定了之后种子是休眠还是萌发。本试验表明,种子萌发过程中赤霉素增加而脱落酸减少,更有利于辽东楤木种子的萌发,其中。200 mg/kg的外源赤霉素浸种效果最佳,能够显著提高发芽率。种子后熟过程中,种皮透性会增强,种子中抑制剂含量降低,使种子逐步达到可萌发条件。当种子打破休眠,首先会分解种胚中大量贮藏的可溶性糖和可溶性蛋白质,为胚的伸长和新陈代谢提供更多的能量[16]。GA3处理种子后, 可使活力毛竹可溶性蛋白质含量升高[17],使华重楼种子可溶性蛋白质含量呈下降趋势[18],可使杜鹃花瓣中可溶性糖含量显著提高[19]。本试验中,辽东楤木种子经200 mg/kg浓度的赤霉素处理后,可溶性糖含量逐渐下降而可溶性蛋白质含量逐渐上升,说明一定浓度的赤霉素能够促进辽东楤木种子可溶性糖的水解和可溶性蛋白质的合成。种子萌发过程中可溶性糖的降低,可能是在外源赤霉素的作用下,利用糖代谢循环的速度加快,呼吸消耗的可溶性蛋白质高于淀粉水解的速度。大多数可溶性蛋白质为代谢酶类,种子萌发过程中,酶类含量的变化在一定程度上能够说明种子代谢活性的强弱。因此可溶性蛋白质逐渐升高,可能是由于种子萌发的相关代谢酶的积累。可见,200 mg/kg赤霉素处理能够促进辽东楤木种子可溶性糖的水解和可溶性蛋白质的合成。
过氧化物酶(POD)、过氧化氢酶(CAT)、超氧化物歧化酶(SOD)是种子内部抗氧化保护系统的重要保护酶,相互之间共同作用来保护种子内部细胞[20]。CAT具有防止过氧化的功能,广泛的存在于生物体中。POD、SOD是植物体内具有生理活性的重要的活性氧自由基清除酶,其中POD能够水解细胞代谢过程中产生的H2O2,从而保护细胞组织[21],与植物的呼吸作用、木质素的合成以及生长素的氧化等都有着密切的联系。SOD是生物体内清除自由基的主要物质,能够防止自由基损害细胞并修复受损细胞[22];SOD可以催化超氧自由基歧化反应生成H2O2,产生的H2O2再由CAT和POD清除[23]。本试验中,层积后的辽东楤木种子在200mg/kg外源赤霉素处理后3种酶活性逐渐升高,且在50d时过氧化物酶、过氧化氢酶和超氧化物歧化酶的酶活性均达到最高值,可见,赤霉素处理辽东楤木种子后可提高POD、CAT及SOD活性,减少了自由基对萌发过程中种子细胞的损伤,进而促进了辽东楤木种子的萌发。这也与前人在露蕊乌头种子、福建青冈种子及丝瓜种子的研究结果相一致[24-26]。总之,一定浓度外源赤霉素处理辽东楤木种子,对打破种子休眠、提高种子发芽率有促进作用。其中,以200mg/kg浸种效果最好。高水平的酶活性(POD、CAT、SOD)、可溶性蛋白质含量、赤霉素含量以及低水平的可溶性糖含量、脱落酸含量有利于辽东楤木种子的萌发。
