杂交水稻的系谱分析与杂种优势利用
2020-10-09杨金松黄志谋张再君邱东峰
杨金松,黄志谋,张再君,邱东峰
(1.湖北省农业科学院粮食作物研究所,湖北武汉430064;2.湖北省咸宁市农业科学院,湖北咸宁437000)
从20世纪70年代野败细胞质不育水稻被发现以来,杂交水稻研究取得了很大的成绩,诸如一系列三系败育细胞质的发现应用、湖北光敏感核不育水稻的发现利用及汕优63、Ⅱ优838、Ⅱ优725、两优培九、扬两优6号等系列品种的选育等,可以看出,从杂交水稻的种质创新到品种选育均取得了一系列的成果。同时亦应看到,从20世纪80年代至今,杂交水稻育成品种的产量出现了徘徊局面,例如:20世纪80年代用化学杀雄方法配制的杂交种赣化2号,其产量已达到14 117.25 kg/hm2[1],现在的育成品种超过这个产量的不多,不同时期选育的代表性品种诸如扬两优6号、两优培九、Ⅱ优725、Ⅱ优838、汕优63等之间产量差异较小[2];杂交水稻的三系和两系虽是在不同的年代配套成功并应用于生产,但并未出现两系杂交稻研究之初的乐观估计——实现两系杂交种比三系杂交种增产15%的目标[3],现实情况是两系杂交稻与三系杂交稻的推广杂交种相比产量上并无明显优势,两者并存发展并无取代的趋势。如何持续提高水稻杂交种的产量潜力值得讨论。本文从品种选育的系谱入手分析、探讨,仅供同行参考。
1 杂交种的亲本系谱分析
从20世纪70年代以来,利用野败细胞质及其他的不育胞质资源,选育了大量的三系不育系品种;自石明松先生发现了光敏核不育水稻资源后[4],又选育了大量的两系不育系品种。三系法杂交稻配套在前并应用于生产,尔后生产上出现大量两系法、三系法杂交种推广种植。分析这些品种的系谱可为育种研究提供一些启发。
1.1 不育系的系谱
20世纪70年代初,袁隆平和他的助手李必湖等在海南岛三亚发现花粉败育的野生稻株(称为野败型细胞质雄性不育,CMS-WA),之后不同的水稻不育细胞质陆续被发现。截至2002年,据不完全统计,共发现60多种水稻细胞质源不育类型资源,选育出600多对不育系和保持系品种[5]。在生产上应用的主要类型有野生稻细胞质类型(分为野败型、矮败型、红莲型等)、籼型栽培稻细胞质类型(主要有冈型、D型、印尼水田型、马协型等)、用籼粳杂交方法产生的粳型不育系(有籼型细胞质—BT型、粳型细胞质—滇型,二者区别在于细胞质的差异,细胞核均为粳型)。由于来源于不同细胞质的不育系众多,仅选取有代表性的品种系谱,呈列表1。
从表1可以看出,以野败型为代表的野生稻细胞质类型不育系的回交亲本均用到华南、长江中下游地区的早稻生态型的品种资源。野败型、矮败型、冈型、D型、马协型、印尼水田型不育系的细胞质源尽管来源不同,限于不育性的保持需要,其不育性(孢子体不育)的保持均要使用到华南、长江中下游地区的早稻生态型品种资源中的细胞质可育基因,以期可循环产生供生产利用的不育系。因此,早期三系不育系品种多为早稻生态型品种,近期略有改变,但亲本血缘成分变化不大。红莲型杂交稻不育性(配子体不育)的保持和恢复机制与野败型恰好相反,但配套互作亦要遵循相似的技术要求。杂交粳稻生产上面积不大,在此不作重点叙述。两系不育系列举了2个代表品种培矮64S、广占63S;两系不育系受不育株来源于粳稻和选育籼粳亚种间杂交种的技术路线,亲本都使用了广亲和资源材料,如培迪、轮选422,亲本使用上来源广泛,但亦显示了其无章法的实用主义特点。
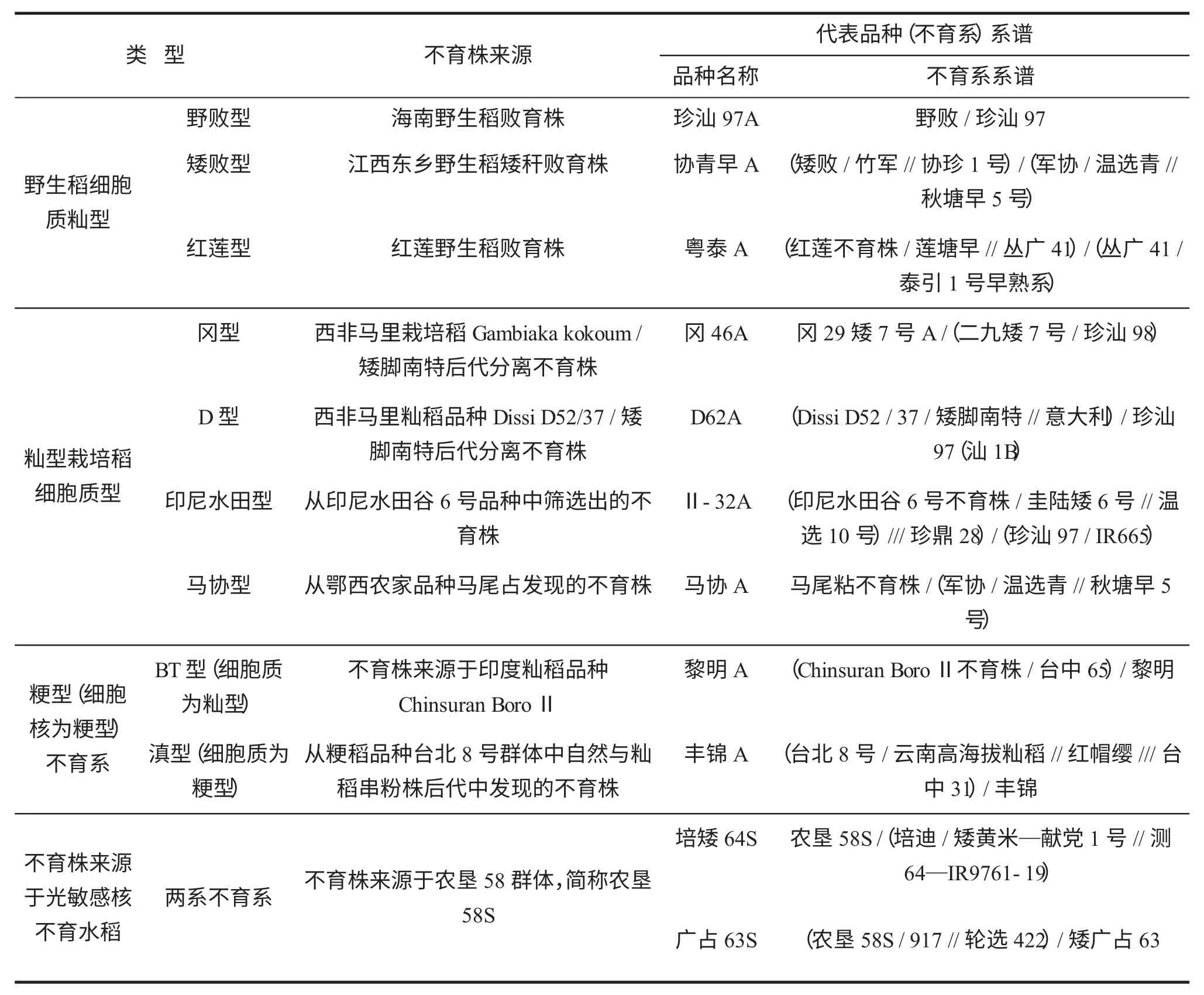
表1 三系杂交稻的不育细胞质类型系谱[6]
1.2 恢复系的系谱
三系不育系的育性恢复需要东南亚、南亚的中籼稻生态型品种资源中的育性恢复基因在配套中发挥遗传作用,以期产生供生产使用的可育杂交种。因此,恢复系的筛选、选育必须围绕着这些有恢复基因的特定种质资源进行研究工作。从图1可以看出,辐恢838、明恢63、绵恢725、蜀恢527等有代表性的恢复系概莫能外,追索其系谱血缘大多含有南亚、东南亚的有恢复基因的中籼稻生态型品种。野败类型的不育系育成以后,第一代的恢复系以常规品种直接测优筛选产生,前期的筛选主要有IR24、IR26、IR661、泰引1号、密阳46、IR9761-19-1等[7]。第二代恢复系以这些种质资源为基础开始进行杂交选育,早期选育的恢复系以明恢63为代表,用其与不同的不育系配组选育了一大批杂交种,主要以汕优63为代表;明恢63之后杂交选育出的代表性恢复系有绵恢838、绵恢725、蜀恢527等。几十年来,三系恢复系选育的技术路线变化不大,仅有变化为掺入一些不同来源的种质资源,如绵恢725有培迪、献党1号的血缘。生物技术应用于育种后,主要是用在单个质量性状改良上,诸如抗病性、米质等方面应用效果显著,在产量性状方面影响不大,其对恢复系主要系谱血缘影响亦小。
两系不育系育性转换受环境温度控制,配组相对自由。从代表性品种两优培九、扬两优6号、Y两优1号等品种来看,扬稻6号及其衍生系是两系恢复系的重要骨干亲本。从图2来看,扬稻6号亦含有IR24、IR26、皮泰等东南亚品种的血缘;IR24、IR26等是以野败为代表的三系杂交稻早期的主要恢复系及后来育种上利用的重要种质资源;皮泰是IR系列品种的重要骨干亲本。扬稻6号虽对野败为代表的三系不育系不具有较强的恢复能力,其与三系恢复系已有明显的遗传分化,但其血缘组成与三系恢复系具有共祖相似性。另有用三系的恢复系蜀恢527等与两系不育系配组产生的杂交种应用于生产。两系恢复系与以野败为代表的三系恢复系在亲本血缘上有着密切的联系[2]。
2 水稻杂种优势育种实践探讨
2个基因型不同的水稻品种杂交产生的F1代在产量等性状方面优于双亲的现象被称为水稻杂种优势。以共祖为特点的杂种优势群理论的育种实践在玉米上成效显著,可为杂交水稻育种借鉴。一般认为,以野败为代表的三系杂交稻的杂种优势产生模式主要以东南亚、南亚的IR系列等中稻型品种(系)和我国的华南及长江中下游地区早稻型品种(系)的这2个生态型不同品种(系)杂交产生的[9]。从扬稻6号及其衍生系、培矮64S、广占63S等品种的系谱来看,两系杂交稻恢复系和不育系都含有一些三系恢复系亲本(东南亚品种)血缘,可以说两系杂交稻的杂种优势来源类似于三系杂交稻。
两系、三系杂交稻研究多年以来,育成杂交种产量出现徘徊、增加缓慢的现象,寻其原因,可从杂种优势群的分析看出一些端倪,有学者认为:三系恢复系和不育系(保持系)基本上分属两大杂种优势群,即南亚、东南亚中籼生态型品种群和中国华南、长江流域地区早籼生态型品种群[9-14]。多年来,三系杂交稻的育种基本上是围绕着这2个杂种优势群工作的,使用的品种资源不多,近期虽有改进,但多表现在抗性和米质等单基因上。种质资源方面创新不够,延用骨干亲本也不够丰富。据统计,截至2010年,利用明恢63作为骨干亲本,先后至少育成了617个新恢复系,其中543个恢复系配组的922个组合通过省级以上品种审定,167个组合通过了国家级品种审定。从1990—2009年,以明恢63作为骨干亲本衍生的杂交种累计推广面积达8 101.3万hm2,占全国杂交水稻推广面积的28.22%[13]。没有挖掘、寻找新的杂种优势群,三系亲本无新的杂种优势群依托,故而杂种优势群内的遗传多样性相对贫乏,这可能是制约三系杂交稻育种产量进一步提高的原因[9]。两系杂交稻育种虽突破了三系杂交稻核质互作的限制,但受到三系杂交稻育种经验的惯性思维影响,在不育系和恢复系的选育上都用到了三系恢复系已使用的种质资源。有研究认为,两系和三系杂交种的杂种优势主要是来自不育系和恢复系的品种间差异,与不育类型并没有必然联系[10]。两系杂交稻的杂种优势来源可能并没有突破三系杂交稻的杂种优势模式。多年以来,在水稻杂种优势群创新研究上乏善可陈,这可能是制约杂交水稻育种产量上升的重要原因。
3 杂交水稻育种未来发展探讨
两系杂交稻及三系杂交稻育种为水稻生产的产量提高作出了很大贡献。三系杂交稻育种实践限于细胞核与细胞质的互作配套,在不育系及恢复系上有选择性的使用亲本并已形成两大杂种优势群,育种成效较大,但近期产量育种陷于瓶颈;两系杂交稻育种可能限于三系杂交稻育种实践的惯性思维及种质资源创新性不够,育种产量增加未见明显突破。对于育种从业者来讲,品种的产量没有最高,只有更高。杂交水稻如何在育种上进一步提高产量值得思考。
3.1 利用生物技术手段,助力杂交水稻产量育种
纵观作物育种的历史,重大的突破不外乎2条途径,即新技术发明应用和新材料的发现应用。生物技术的日益发展为杂交水稻产量育种发展提供了许多新的可能。邓兴旺等使用基因编辑技术,利用np1突变体r的雄性不育基因,连接相关基因启动子,采用转基因手段整合形成可循环使用的新类型水稻保持系,可分离出保持系及稳定的不育系品种,重复此分离过程形成体系,被称为第三代杂交水稻育种新技术,并已应用于杂交水稻育种[11-12]。新类型不育系的不育性遗传受隐性单核基因控制,可与常规品种杂交产生可育的F1代。利用第三代水稻杂交技术,不用杂交转育手段即可使常规水稻品种转化成新的不育系,可使任何2个常规水稻品种自由杂交形成可供生产使用的水稻杂交种,因此,可摆脱两系、三系杂交水稻的不育基因的转育或回交保持的羁绊。可效仿杂交玉米育种的经验,以品种间的杂种优势为筛选目标,以品种系谱来源为基础,重新构建以共祖为特点的水稻杂种优势群,形成品种(系)系谱来源清晰、血缘平行不交叉的杂种优势群,并且不断给杂种优势群内补充新的种质资源,从而不断提高品种的杂种优势,在育种上持续提高杂交水稻的产量。
3.2 加强水稻远缘杂交研究,提高水稻的杂种优势
水稻的远缘杂交产生的杂种优势巨大,近期的“粳不籼恢”模式取得了一些进展,这个现象值得鼓励和重视。要加强力量持续进行籼粳杂交在理论基础和应用实践方面的研究,以期获得更大突破。广泛进行籼稻、粳稻与爪哇稻等品种之间的远缘杂交研究,寻找新的水稻杂种优势群,同时强化水稻杂种优势理论研究也是必要的。
3.3 整合杂交水稻研究力量,为提高杂交水稻产量作贡献
目前国家层面有水稻研究所及杂交水稻研究中心,大部分省(市)级农业科学院有水稻研究所或水稻研究团队,省以下的地(市)级农业科学院亦有专门的水稻研究团队,这些水稻研究机构或团队为全国的水稻杂交种改良及杂交水稻生产作出了重要贡献。但随着科学技术的发展,这种机构设置已不能适应目前生产需要,主要表现在应用研究内容低水平重复,选育品种同质化现象严重,造成大量的人力资源浪费。全国几大稻区内的不同省份之间存在的此种现象更严重。因此有必要从顶层设计上整合全国的杂交水稻研究力量,夯实基础研究,并按稻作区划清晰明确各级研究单位或团队的研究任务以期提高工作效率,切实提高杂交水稻育种水平。可在全国范围内按稻区分布重新组合研究力量,做到各级分工明确,上、下游单位之间工作内容贯通,各单位之间研究任务区分明确,进一步在国家范围内建立跨上下游单位、跨省(市)的高效率的杂交水稻研究大团队。国家级单位侧重于基础理论研究与种质资源创新,规划指导省级及以下单位的研究工作;省级单位主要从事应用基础研究、新品种选育等方面工作,同一稻区的省级研究机构之间研究内容要建立互动、互补机制,研究内容各有侧重,以减少重复性工作;省级以下的研究机构主要从事品种生产推广试验、示范、试种及品种栽培等方面的研究工作,以期在生产上最大限度地发挥品种产量潜力。通过整合国家各层面单位的杂交水稻研究力量,提高工作效率,以期选育出更多、更优秀的水稻杂交种,为提高杂交水稻产量作出更大的贡献。
