华南地区裸子植物与被子植物季节性水分利用的比较研究
2020-10-09叶慧莹赵平李艳琼
叶慧莹, 赵平, 李艳琼
(1. 中国科学院华南植物园, 广东省应用植物学重点实验室, 广州 510650; 2. 中国科学院大学, 北京 100049)
中国是世界上裸子植物物种最丰富的国家[1],种类高达197种,约占世界的17.6%[2]。有学者认为,在温暖湿润地区, 具有管胞的裸子植物输水效率低于具有导管的被子植物[3-4]。由于大部分裸子植物的叶片面积小,叶片较为密集,有更高的总叶面积,但由于叶片缺少导管而导致较低的光合速率[3],在水热条件良好的环境下,裸子植物的最高生长率通常要低于被子植物[3]。裸子植物有更强的冠层截流能力,使降水更难到达土壤中,导致土壤更加干旱[5]。被子植物比裸子植物有更高的细根密度和细根量,获取土壤中水分的能力更强[6]。从整树尺度上比较裸子植物和被子植物的水分利用特征,能从个体角度对两者有更深入直观的了解,为两者的起源发展作补充研究。
华南地区气候温暖湿润,植被多样性丰富,每日的蒸腾耗水量是巨大的,无论是裸子植物还是被子植物,为满足生命活动的需要,都要从土壤吸收水分,通过根系运输到各个器官,并以蒸腾作用的形式从叶片、枝条或树干向外界消散水汽。水力导度反映了树木从土壤到叶片的水分传输能力[7],可影响气孔的水分调节及气体交换,并影响植株的光合作用和生长速率[8]。在寒冷干旱背景下裸子植物与被子植物对环境的适应性已有研究[9-11], 但对我国华南湿润地区裸子植物与被子植物水分利用的对比研究并不多。与大多研究的试验地区不同的是,华南地区温暖湿润,更有生活在水中的裸子植物,在这样不受水分限制的情况下,裸子植物和被子植物的输水效率有何差异。
为深入了解裸子植物和被子植物蒸腾耗水的生理过程和差异,我们选择2种在华南地区常见的水生裸子植物池杉(Taxodium ascendens)和落羽杉(T.distichum),以及2种被子植物枫香树(Liquidambar formosana)和乐昌含笑(Michelia chapensis),利用Granier热消散探针[12-13]连续监测4树种的树干液流,计算整树水力导度和整树蒸腾速率,对比分析裸子植物与被子植物水分利用的季节差异及其与环境的关系,为园林绿化配置过程中树种的选择以及养护中的水分管理提供科学依据。
1 材料和方法
1.1 研究样地概况
研究样地在广东省广州市天河区中国科学院华南植物园内(23°11′ N, 113°21′ E),分别位于科研区试验田、研究生公寓旁和藤本植物园。该区属于典型的亚热带季风性湿润气候,夏季高温多雨,冬季温和少雨,有明显的干湿季。年均太阳辐射为4 365~4 600 MJ/m2[14],年均气温为21.4~21.9℃,年降雨量约为1 700 mm,全年约85%的降水集中在4-9月。
1.2 材料
选择华南地区常见的2种裸子植物池杉(T.ascendens)和落羽杉(T. distichum),2种被子植物枫香树(Liquidambar formosana)和乐昌含笑(Michelia chapensis)作为试验材料。每种树选择6株径级接近的样树,共计24株。
1.3 树形特征的测定
用激光测高仪(TruPulse200, USA Laser Technology)测量样树的树高(H);用胸径尺测量距离地面1.3 m处的直径(DBH);用软皮尺测量冠层投影的长和宽, 其乘积即为冠幅(AC);用生长锥钻取木条后,根据边材和心材颜色差异用直尺测量边材宽度,计算边材面积(AS)。
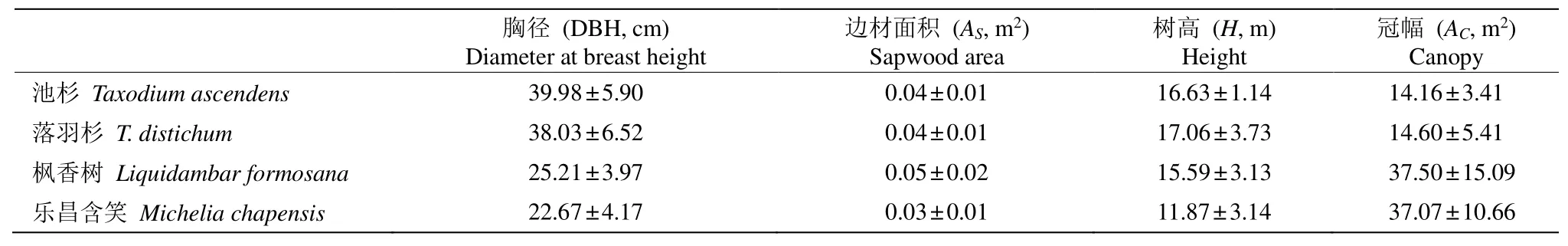
表1 样树的形态特征Table 1 Morphological characteristic of sample trees
1.4 环境因子的监测
本研究参考华南植物园小青山生态观测场的环境数据。光合有效辐射传感器[PAR,µmol/(m2·s)]安装在荷木林冠层上方约1 m处,空气温度(T, ℃)和相对湿度(RH, %)的传感器安装在观测塔内距地面高约18 m处,即近树冠高度的三分之二处。传感器均与DL2e数据采集仪(Delta-T公司, 英国)相连,测读频率为30 s,每10 min求1次平均值并存储。
空气水汽压亏缺(VPD, kPa),表征RH和T的协同效应[15],VPD=a×exp[bT/(T+c)](1-RH), 式中,a、b和c分别是0.611 kPa、17.502和240.97℃。
1.5 树干液流的测定
将1对长20 mm的Granier热消散探针平行安装在北面树干胸高边材里,上下探针垂直相隔10~15 cm。用塑料盖罩住探针以防止机械损伤,外部包裹太阳膜以减少太阳辐射和防止雨水渗入。用四芯屏蔽电缆连接探针和DL2e型数据采集仪,测读频率为30 s,每10 min求1次平均值并存储。上探针外面缠绕1层加热丝,通过供应0.12 A的直流电进行加热,下探针不加热与边材温度保持相近, 2个探针间产生温度差。在树干液流的影响下,2个探针间的温度差会发生改变,可用于计算树木的瞬时液流密度[Js, g/(m2·s)][16], 即单位时间通过单位边材面积的液流量,Js=119×[(ΔTm-ΔT)/ΔT]1.231, 式中,ΔTm为液流为0时的温差;ΔT为瞬时温差。利用美国杜克大学环境学院开发的Baseliner 3.0软件将温差数据转化为液流密度值。考虑液流沿边材的径向变化[17],0~4 cm处边材为高液流密度区,4 cm以上处边材为低液流密度区,低密度区平均液流量约为高密度区的45%,因此日均液流密度EA[kg/(m2·d)]采用加权平均值来表示[18-19]:EA=式中,Asi为单株样树边材总面积(m2);As1i为单株样树厚度0~4 cm部分的边材面积(m2);Jsi为单株样树的瞬时液流密度[g/(m2·s)];t为1 d的时间8.64×104s。

为比较4树种在单位时间内整树蒸腾的耗水量,即整树的蒸腾速率,选择天气晴朗,气象因子相接近的干季(3月11-13日)和湿季(7月15-18日)数据,分析4树种的整树蒸腾速率(ET,g/s)的日变化特征,ET=Js×As1+Jsi×As2×45%, 式中,Js为单株样树的瞬时液流密度[g/(m2·s)];As1为单株样树厚度0~4 cm部分的边材面积(m2),As2为单株样树厚度大于4 cm部分的边材面积(m2)。
累计单株整树日均耗水量(Qd, kg/d)根据Qd=[(Js×As1)+(Js×As2×45%)]×10-3×t计算。根据整树蒸腾速率(ET)计算整树水力导度[k, g/(s·MPa)][20],k=ET/(φs-φL-ρw·g·h), 式中,φs是凌晨叶片水势(MPa),φL是正午冠层叶片水势(MPa);ρw是水密度(998 kg/m3);h是树高(m);g是重力加速度(9.8 N/kg)。
1.6 叶片水势的测定
在2019年3月1-3日和7月15-17日的晴朗天气下,分别在黎明前(4:00或5:00)、正午(13:00或13:30)和日落后(19:00或20:00)测定叶片水势[19],每株样树分别剪取至少3枝带叶枝条,利用PMS压力室(PMS Instruments, Oregon, USA)即时测定叶片水势(φL, MPa)。
1.7 数据的统计分析
使用Microsoft Excel 2019和SPSS 20.0统计分析软件进行数据处理和分析,使用Origin Pro制图,采用Duncan检验、单因素方差分析和独立样本t检验进行数据的差异显著性分析。空气温度和相对湿度受太阳辐射的影响,VPD反映空气温度和湿度的共同效应,为了避免重复分析相互影响的环境因子,4种样树整树蒸腾(ET)与光合有效辐射(PAR)及VPD的相关关系采用偏相关分析法。
2 结果和分析
2.1 日均液流密度的干湿季变化
从图1: A可见,在干季(3月),池杉和落羽杉的日均液流密度(EA)显著低于枫香树和乐昌含笑(P<0.05),枫香与乐昌含笑的EA较高,分别为404.63和457.11 kg/(m2·d);池杉与落羽杉的较低,分别为71.74和123.83 kg/(m2·d)。从图1: B可见,在湿季(7月),裸子植物池杉和落羽杉与被子植物枫香树和乐昌含笑的EA没有显著差异(P>0.05),枫香与乐昌含笑的EA分别为745.87和1 154.66 kg/(m2·d);池杉与落羽杉分别为1 071.46和990.67 kg/(m2·d)。这说明在水热条件良好的情况下,裸子植物单位面积边材的输水效率与被子植物相近。
2.2 整树蒸腾速率的干湿季变化
在干湿季的晴朗天气下,4种样树的整树蒸腾速率(ET)均呈现出明显的日变化,与光合有效辐射(PAR)、空气水汽压亏缺(VPD)呈现出一致的变化趋势(图2: A, B)。从图2: C, D可见,干湿季池杉和落羽杉均在14:00左右达到蒸腾速率峰值,而枫香树和乐昌含笑则在13:00左右达到峰值,裸子植物的ET峰值出现时间要晚于被子植物,这可能是裸子植物与被子植物木质部结构不同而导致的。干季池杉和落羽杉的ET显著低于枫香树和乐昌含笑(P<0.05), 其中枫香树的最高,池杉的最低。湿季池杉和落羽杉的ET显著高于枫香树和乐昌含笑(P<0.05),其中池杉的最高,乐昌含笑的最低。

图1 裸子植物与被子植物日均液流密度(EA)的干(A)湿(B)季变化。CS: 池杉; LYS: 落羽杉; FX: 枫香树; LCHX: 乐昌含笑。以下图表同。Fig. 1 Changes in daily sap flow density (EA) of gymnosperms and angiosperms in dry (A) and wet (B) seasons. CS: Taxodium ascendens; LYS: T. distichum;FX: Liquidambar formosana; LCHX: Michelia chapensis. The same is following Figures and Table.
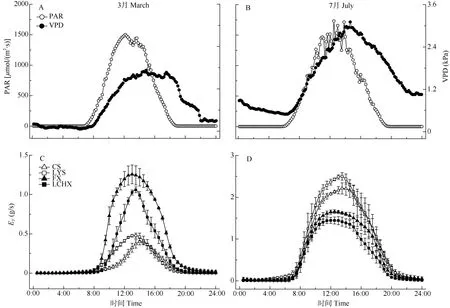
图2 光合有效辐射(PAR)、空气水汽压亏缺(VPD)、裸子植物和被子植物的整树蒸腾速率(ET)的干(C)湿(D)季日变化Fig. 2 Daily changes in photosynthetical active radiation (PAR), vapor pressure deficit (VPD) and whole tree transpiration rate (ET) of gymnosperms and angiosperms in dry (C) and wet (D) seasons
2.3 整树水力导度的干湿季差异
如图3所示,在干季(3月),2种裸子植物池杉和落羽杉的整树水力导度(k)都显著低于2种被子植物枫香树和乐昌含笑(P<0.05);池杉与落羽杉间、枫香树与乐昌含笑间没有显著差异。在湿季,4种样树的整树水力导度比干季都显著增加,池杉和落羽杉的整树水力导度显著高于枫香树和乐昌含笑(P<0.05 );裸子植物间和被子植物间有显著差异(P<0.05)。
2.4 偏相关分析
偏相关分析可见,4种样树的整树蒸腾速率(ET)都与光合有效辐射(PAR)和空气水汽压亏缺(VPD)存在偏相关关系,池杉的ET在干湿季与PAR的偏相关程度都要低于VPD的。而落羽杉、枫香树和乐昌含笑的ET在干湿季与PAR的偏相关程度都要高于VPD的。因此,PAR与VPD对不同树种的ET存在不同的影响。

图3 裸子植物与被子植物整树水力导度(k)的干湿季变化。柱上不同小写字母表示干湿季差异显著;不同大写字母表示不同树种同一季节差异显著。Fig. 3 Change in whole tree hydraulic conductance (k) of gymnosperms and angiosperms in dry and wet seasons. Different small letters indicate significant difference between dry and wet seasons at 0.05 level, while different capital letters indicate significant difference among different species at the same season at 0.05 level.

表2 整树蒸腾速率(ET)与PAR和VPD的偏相关关系Table 2 Partial correlations of whole tree transpiration rate (ET) with PAR and VPD
对4种样树的日均耗水量(Qd)和环境变量(PAR或VPD)进行拟合,无论是干季还是湿季,Qd都随PAR增加呈现线性增长趋势,Qd都随VPD的增加呈现出对数增长趋势(图4)。拟合曲线的斜率反映了蒸腾速率对气象因子响应的敏感性,湿季各树种的Qd对VPD响应曲线的斜率高于干季,说明湿季各树种对VPD的响应比较迅速(图4: B, D)。在干季,池杉和落羽杉的Qd对PAR和VPD响应曲线的斜率显著低于枫香树和乐昌含笑的(图4: A, B), 从图4: C,D可见,池杉和落羽杉的Qd在湿季对PAR和VPD响应曲线的斜率高于枫香树和乐昌含笑。湿季池杉和落羽杉对PAR和VPD响应曲线的斜率非常接近,说明这2种树在湿季对PAR和VPD的响应趋同; 但在干季时落羽杉对PAR和VPD响应曲线的斜率明显高于池杉,说明落羽杉在干季时的储水能力可能要高于池杉。无论是干季还是湿季,枫香树对PAR和VPD响应曲线的斜率都要高于乐昌含笑。
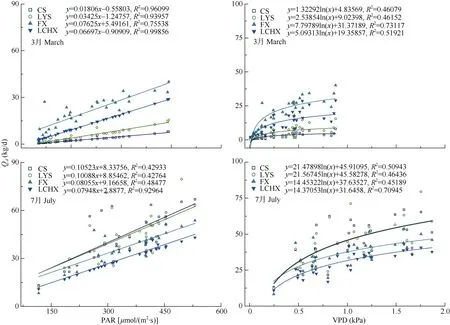
图4 干湿季植物整树日均耗水量(Qd)与光合有效辐射(PAR)、空气水汽压亏缺(VPD)的关系Fig. 4 Relationship between photosynthetically active radiation (PAR), vapor pressure deficit (VPD) and daily water consumption (Qd) of plants in dry and wet seasons
3 结论和讨论
我国裸子植物多集中在山地[21],物种多样性总体呈南高北低趋势,裸子植物与被子植物的比例受能量和水分的显著影响,当能量降低和水分减少时,裸子植物所占的比例会显著升高[22]。系统发育显著影响裸子植物的木材性状,裸子植物的基本密度、气干密度、径向收缩系数、切向收缩系数、体积收缩系数、抗弯强度均显著低于被子植物,但横截面硬度却显著高于被子植物[23]。金鹰等[24]的研究表明,我国3种温带地区裸子植物7月的叶导水率显著低于同地区的6种被子植物。虽然裸子植物和被子植物的水力结构不同,但并不代表裸子植物对水的运输能力比被子植物弱。虽然裸子植物的传输速率比被子植物慢,但有些裸子植物的整体运输量还是惊人的,加州红杉(Larix potaninii)从叶片到根的运输距离可达110 m[25]。
整树水力导度指的是植物将水分从根部附近的土壤运送至冠层的效率,树木会形成不同的形态结构和水分输送策略来适应生存环境,改变整树水力导度能调节植物在生长发育过程中对水分的需求[26]。整树水力导度是树木水分传输效率在树木水力结构上的间接反映[27]。裸子植物与被子植物在树木水力结构上有着极大的差异,大量的研究表明裸子植物的管胞结构在运输能力上要比被子植物的导管结构弱[3,25,28]。华南地区属于亚热带季风性湿润气候,湿季降雨充足且气温较高,适合多种植物生长,周娟等[26]的研究表明,华南地区的被子植物荷木(Schima superba)在湿季的水力导度比干季下降,但并不显著,这与本研究中2种被子植物样树的结果一致。程静等[29]的研究表明,华南鼎湖山地区的裸子植物马尾松干湿季的液流密度与被子植物锥栗和荷木相近,且干湿季液流密度差异显著,这也与本研究结果一致。
Caitlyn等[30]在美国得克萨斯州的研究表明,裸子植物火炬松(Pinus taeda)的蒸腾速率显著高于2种栎属被子植物(Quercus stellata和Q. marilandica),火炬松的液流密度比这2种栎属植物高约36%,这与本研究中湿季的结果一致。在华南地区,裸子植物池杉和落羽杉的整树蒸腾和整树水力导度在湿季(7月)均显著高于被子植物枫香树和乐昌含笑。总体来说,4种树的整树水力导度从干季到湿季都有显著增长,其中,池杉和落羽杉的涨幅最大,这可能与池杉与落羽杉的生长习性相关,池杉和落羽杉是落叶植物,在干季整树叶面积急剧降低,整树水力导度也会随之下降。池杉和落羽杉都是华南地区常见水生树种,池杉和落羽杉都有较强的耐水性,但耐旱性较弱[31-32],华南地区湿季水热条件良好,这2种裸子植物的生长环境几乎不会出现干旱缺水的情况,可能由此导致两者整树水力导度的剧增。
PAR主要调节叶片气孔开度,VPD则是蒸腾作用的驱动力,所以这2个因子是控制蒸腾最主要的气象因子。很多研究表明环境因子PAR和VPD对树木蒸腾耗水有一定的影响,不同季节的PAR和VPD显著影响荷木的整树蒸腾,但影响程度有所不同[13],推测可能是由荷木的内在调节造成的;柠檬桉(Eucalyptus citriodora)和金合欢(Acacia auriculaeformis)的液流密度与PAR和VPD都呈显著正相关[33]; 有研究表明,柚木(Tectona grandis)的液流密度在干湿季都与VPD呈显著相关,但与PAR的相关程度较弱[34]。在本研究中,4种样树干湿季的日均耗水量和整树蒸腾速率都与PAR和VPD有显著相关关系,但在不同季节的相关程度不同,这与倪广艳等[19]的研究结果一致。罗紫东等[35]报道,枫香树的叶子在干季的衰老过程中还能进行光合作用,也就意味着蒸腾和呼吸作用还在持续,这与本研究中干季枫香树树干仍有液流的结果相符合。
研究表明,温带森林中的裸子植物具有很高的耐受适应性,往往分布在相对寒冷贫瘠的地方[36]。生存环境对裸子植物的水分运输能力的影响很大,在华南温暖地区,裸子植物池杉和落羽杉的根部对缺氧的水下环境具有较强的耐受适应性,使自身长期处于一种不受水分限制影响的环境中,这可能是植物对环境的一种适应策略,这种策略使这两种裸子植物在华南地区湿季可以达到比被子植物更高的水分运输效率。从本研究结果来看,池杉和落羽杉的整树蒸腾速率受环境因子的影响,在湿季有显著提高, 且高于枫香树和乐昌含笑。由此, 我们推测,裸子植物的管胞输水效率在适宜的环境条件下高于被子植物的导管。裸子植物在地球上出现的时间远比被子植物早,本研究结果能从水分运输方面说明裸子植物对环境有更强的适应能力, 也从侧面反映了裸子植物能够在地球上存活较长时间的原因。
华南地区水热充沛、河网密布,池杉和落羽杉能很好地适应水岸边的生活环境,本研究对华南地区森林修复,人工林以及园林造景的树种选择具有重要的参考价值。从微观角度对池杉、落羽杉、枫香树和乐昌含笑的水力结构对比还有待进一步研究。目前,对于从整树尺度上对比裸子植物和被子植物水分运输差异的研究还比较少,日后还应继续深入研究其他多种裸子植物与被子植物的水分运输差异。
