白木通的光合特性
2015-09-10吴玲利熊利柯镔峰等
吴玲利 熊利 柯镔峰等

摘要: 为了研究白木通光合效率及不同年龄叶片之间光合特性的差异,利用Li-6400xt便携式光合仪(Li-COR,USA)对其光合特性日变化、光合-光响应曲线及CO2响应曲线进行测定。结果表明:白木通二年生盆栽苗2013年生老叶及2014年生新叶的净光合速率、气孔导度的日变化均呈双峰曲线,老叶的蒸腾速率呈单峰曲线,而新叶的蒸腾速率呈双峰曲线;老叶和新叶的胞间CO2浓度在07:00—11:00缓慢下降,到11:00时最低,分别为244 16、309 28 μmol/mol,下午又缓慢回升;水分利用效率的日变化呈不规则的U形变化;影响白木通叶片净光合速率主要生态生理指标的因素从大到小依次为气孔导度>胞间CO2浓度>蒸腾速率>光合有效辐射。白木通老叶和新叶的光补偿点分别为3111、28 54 μmol/mol,光饱和点分别为854 17、817 32 μmol/mol,对光的适应范围较小;老叶和新叶的CO2补偿点分别为106 54、111 63 μmol/mol,CO2饱和点分别为 1 870 61、1 382 37 μmol/mol,白木通光合碳同化途径具有C3植物的特性。
关键词: 白木通;叶龄;光合特性;蒸腾速率;羧化效率;日变化
中图分类号: Q945 11 文献标志码: A
文章编号:1002-1302(2015)08-0250-04
白木通(Akebia trifoliate var australis)为木通科木通属的落叶或半常绿藤本植物,别称八月瓜藤,气微弱,味苦而涩,性微寒,具有清心泻火、疏肝益肾、通经散瘀、除烦利尿之功效,主要分布于江苏、浙江、江西、广西、湖南、湖北、山西、四川等地 [1-4]。目前,关于木通属植物的研究报道主要集中在油的理化特性、资源分布、外观性质的鉴定、化学成分分析及药理学研究等方面,而对木通属植物人工栽培的研究还处于起步阶段,关于光合特性与环境因子关系等方面的研究鲜有报道。光合作用是植物体内重要的生理过程及其他代谢活动的生理基础,是植物药材产量和品质形成的基础,也是评价药用植物生产能力的重要指标 [5-7]。由于其在植物进化过程中的稳定性而成为植物生理生态研究的重要内容,光合作用的日变化特征反映了植物的遗传特性和对环境中水肥气热的适应能力,可以作为判断植物生长和抗逆性的指标 [8]。因此,本研究对白木通的二年生盆栽苗2013年生老叶和2014年生新叶的光合日变化及光合特性进行观测和分析,旨在揭示不同叶龄之间光合日变化的差异、对光的利用能力及光适应范围,为白木通立地条件选择及栽培利用提供理论基础。
1 材料与方法
1 1 供试材料
供试材料白木通由江西省林业科学院提供,为二年生盆栽苗,试验在中南林业科技大学校内进行。
1 2 光合日变化的测定
采用Li-6400xt便携式光合仪(Li-COR,USA)对光合作用日变化进行测定,测定时选取长势及颜色基本一致且无病虫害的2013年生老叶和2014年生新叶,保持叶片自然生长角度,共测定9株,测定老叶和新叶各3张/株,重复3次。测定时间选在2014年7月5—7日晴朗无风光照充足的3 d,07:00—19:00每隔2 h测定1次。测定的光合指标包括光合有效辐射(PAR)、气温(T)、净光合速率(Pn)、气孔导度(Gs)、蒸腾速率(Tr)、胞间CO2浓度(Ci)等参数。水分利用效率(WUE)=净光合速率(Pn)/蒸腾速率(Tr)。
1 3 光合-光响应曲线及CO2响应曲线的测定
光合指标测定在2014年7月8—10日08:00—11:00,选取叶龄不同、长势一致的叶片,用Li-6400xt便携式光合仪(Li-COR,USA)测定光合-光响应曲线及CO2响应曲线;光 合作用光响应曲线用6400-LED红蓝光源自动light-curve测定;CO2浓度由小钢瓶提供,浓度设定为400 μmol/mol, 光合有效辐射梯度设定为2 100、1 800、1 500、1 200、900、600、300、200、150、100、75、50、25、0 μmol/(m2·s)。光饱和点(LSP)、 光补偿点(LCP)等光合指标根据直角双曲线修正模型进行拟合计算 [9]。光合作用CO2响应曲线测定选择A-CI Curve曲线测定,叶室CO2梯度设定为400、300、200、150、100、50、400、600、800、1 000、1 200、1 500 μmol/mol, 设定照度为 1 000 μmol/(m2·s),CO2饱和点、补偿点等参数的拟合方法同上。
1 4 数据统计与分析
用Excel 2007对数据进行处理作图,用SPSS 17 0软件进行相关性及其差异显著性分析。
2 结果与分析
2 1 白木通光合特性的日变化
2 1 1 净光合速率(Pn)的日变化
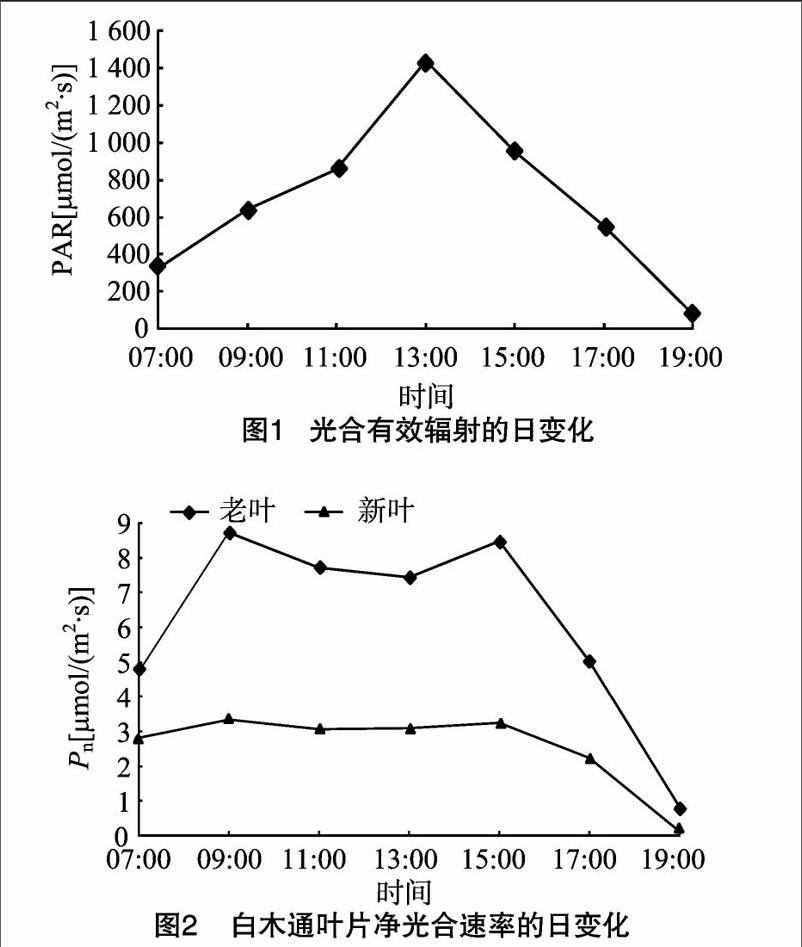
照度是影响植物光合作用的主要因素 [10]。 由图1可知,光合有效辐射(PAR)在 13:00 时达到最高,为1 438 μmol/(m2·s),之后逐渐降低。净光合速率(Pn)是植物光合作用储存碳水化合物的直接体现,它的大小直接决定着植物光合能力的强弱 [11]。由图2可知,白木通新叶和老叶Pn的日变化均呈双峰曲线,出现“光合午休”现象,两者相比,老叶的“光合午休”现象更明显。老叶和新叶的第1峰均出现在09:00,其值分别为8 73、334 μmol/(m2·s),第2峰出现在15:00,其值分别为8 48、3 22 μmol/(m2·s),17:00以后Pn迅速降低,19:00以后Pn接近于0。白木通老叶和新叶的Pn日变化平均值分别为615、2 55 μmol/(m2·s),其中老叶的Pn远高于新叶的Pn。
2 1 2 蒸腾速率的日变化 蒸腾作用是植物生命活动的重要组成部分,它是植物吸收水分和运输矿质营养的主要动力,调节叶片温度,维持植物体内生理活动的有序进行 [12]。如图3所示,白木通老叶Tr的日变化呈单峰曲线,07:00—09:00迅速上升,然后缓慢上升,到15:00达到最大,为 3 49 mmol/(m2·s),15:00以后随着PAR的降低迅速降低。新叶的Tr呈双峰曲线,第1峰出现在09:00,之后有所降低,在11:00出现低谷,在15:00时达到第2峰,其值为 2 93 mmol/(m2·s)。15:00以后白木通新叶和老叶的Tr随PAR与T的降低而降低,到19:00时 Tr的值趋近于0。
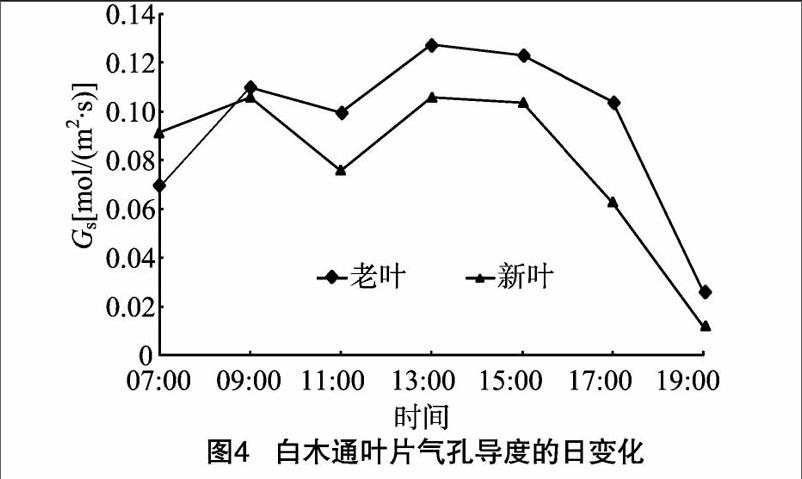
2 1 3 气孔导度的日变化 白木通老叶和新叶的Gs与Pn的日变化呈正相关关系,两者Gs的日变化也呈双峰曲线,07:00 老叶和新叶的Gs已经处于很高的水平,且新叶的气孔导度高于老叶的气孔导度,其值分别为0 07、0 09 mol/(m2·s), 说明弱光对新叶气孔的诱导更明显,弱 光更有利于新叶的光合作用,这也是新叶逐渐适应外界环境的一种直接体现。07:00—09:00老叶的Gs迅速升高,09:00以后Gs随着照度的增加有所降低。老叶和新叶Gs第2峰出现在13:00,分别为013、0 11 mol/(m2·s),老叶的Gs在 17:00 后迅速降低,而新叶的Gs在15:00后迅速降低(图4)。
2 1 4 胞间CO2浓度的日变化 Ci是外界CO2气体进入叶肉细胞过程中所受各种驱动力和阻力以及叶片内部光合作用和呼吸作用最终平衡的结果 [13-14]。由图5可以看出,白木通老叶和新叶的Ci在07:00时较高,其值分别为301 01、361 20 μmol/mol,随着净光合速率及体内酶活性等的增大,进入叶片内的CO2被羧化为碳水化合物,Ci在11:00时达到最低,之后又缓慢上升;15:00以后,随着Pn的降低及环境CO2浓度的升高,Ci缓慢升高。
2 1 5 水分利用效率的日变化
水分利用效率表明植物固定单位数量CO2的耗水量,是植物利用水分的能力和抗旱性的一个重要指标,也是植物对干旱适应的直接体现 [15-16]。由图6可知,白木通老叶的WUE远高于新叶的,老叶和新叶的WUE在07:00时最高,分别为5 98、2 60 μmol/mmol,之后随着PAR的升高及空气相对湿度的降低逐渐减小;老叶的WUE在17:00后迅速回升,而新叶的WUE在15:00后缓慢回升。由此可知,不同叶龄叶片对水分利用的能力不同,白木通老叶对干旱的适应性强于新叶对干旱的适应性。
2 1 6 白木通新叶和老叶的Pn与其Tr、Gs、Ci、WUE、PAR的[CM(25]相关性分析
由表1可知,白木通2013年生老叶和2014
年生新叶Pn的日变化与其Tr的日变化呈显著正相关,与Gs的日变化呈极显著正相关,2014年生新叶的相关系数高于2013年生的老叶。白木通2013年生老叶和2014年生新叶Pn的日变化与PAR的相关系数分别为0 746、0685,说明老叶对光的利用能力更强。2013年生老叶和2014年生新叶Pn的日变化与Ci、WUE呈负相关关系,其中2013年生老叶的Pn与Ci呈极显著负相关关系(P<0 01)。相关分析结果表明,影响白木通老叶、新叶Pn主要生态生理指标的因素从大到小分别为Gs>Ci>Tr>PAR、Gs>Tr>PAR>Ci。
2 2 白木通不同叶龄对光合-光响应曲线的影响
由图7可知,随着光合有效辐射的增加,白木通老叶和新叶的Pn先升高后降低。通过直角双曲线修正模型拟合可知,白木通老叶及新叶的光饱和点分别为854 17、817 32 μmol/mol,此时,老叶和新叶的最大净光合速率分别为8 82、 6 43 μmol/(m2·s)。光补偿点表示植物对弱光的利用能力,白木通老叶和新叶光补偿点分别为31 11、28 54 μmol/mol, 说明白木通新叶更适合在弱光下进行光合作用。表观量子效率(AQY)是植物叶片光能利用效率的一个重要指标,它反映叶片对弱光的利用能力,AQY越高,表明植物吸收和转换光能的色素蛋白复合体较多 [17]。白木通老叶和新叶的AQY分别为0 070、0 060,说明白木通对对弱光的利用能力较强,是较耐阴的植物。白木通新叶和老叶的暗呼吸速率分别为1 89、1 65 μmol/(m2·s),说明白木通老叶的暗呼吸速率高于新叶(表2)。
2 3 白木通不同叶龄光合-CO2响应曲线的比较
羧化效率(CE)反映植物在给定条件下对CO2的同化能力,它体现了植物在低浓度CO2下的光合能力 [18]。由表3可知,白木通老叶和新叶的羧化效率较低,说明白木通在低浓度CO2条件下的光合能力较弱。CO2补偿点是光合作用评价植物CO2同化能力的重要指标,白木通老叶和新叶的CO2补偿点均较高,分别为 106 54、111 63 μmol/mol,CO2饱和点分别为 1 870 61、1 382 37 μmol/mol,说明白木通老叶更适合在高浓度CO2下进行光合作用。由此可知,白木通具有典型C3植物的光合特性。
3 结论与讨论
白木通2013年生老叶和2014年生新叶Pn日变化的平均值分别为 6 15、2 55 μmol/(m2·s),老叶及新叶的净光合速率、气孔导度的日变化均呈双峰曲线,中午有明显的“午休”现象,主要是由中午PAR较强使得光合生理活性降低的非气孔因素引起的 [19]。而白木通老叶的蒸腾速率呈单峰曲线,新叶的蒸腾速率呈双峰曲线。Ci和WUE的日变化呈不规则的U形变化;白木通老叶和新叶的光补偿点分别为3111、28 54 μmol/mol,光饱和点分别为 854 17、817 32 μmol/mol。植物光饱和点与光补偿点直接体现它对光能的利用能力,可用来衡量植物的需光程度,光补偿点低、光饱和点高的植物对光环境的适应性较强 [20]。本研究结果表明,白木通具有较低的光饱和点及光补偿点,说明白木通对光的适应范围较小,更适合在照度较弱的阴坡生长。本研究通过对白木通老叶和新叶光合特性的比较发现,老叶的光饱和点高于新叶,这与老叶光合生理机构更健全、光适应范围更广有关。白木通叶片的羧化效率较低,新叶和老叶的羧化效率都在0 03以下,说明白木通不利于在低浓度的环境下进行光合作用。除此之外,白木通具有较高的CO2饱和点和CO2补偿点,说明白木通具有C3植物的光合特性。
气孔的开放及大小是植物对外界环境适应性的直接体现,也是植物衰老的衡量指标之一 [21-22]。气孔是CO2进入叶片的主要通道,气孔的开闭直接影响植物的净光合速率。本研究发现,气孔导度与Pn呈极显著正相关关系,由图2和图4发现,气孔导度在13:00时达到最大,随后又缓慢降低,在15:00时又达到最高,这与任建武等的研究结果 [23]一致。影响植物净光合速率主要有气孔因素和非气孔因素,气孔因素主要由环境中的气温、辐射强度、空气湿度的变化而引起植株气孔关闭,外界CO2不能通过气孔进入叶片内部,导致光合速率降低;非气孔因素主要是在短时间内气孔扩散阻力不会限制植物的光合能力,主要是植物体内光合酶活性等减弱所致,说明气孔导度的瞬时降低对光合速率的影响较小。因此,在13:00—15:00时净光合速率与气孔导度并不显著正相关,说明主要是非气孔因素限制了白木通叶片的光合效率。Tr的大小反映植物吸收和运输能力的强弱,一定程度上反映了植物调节水分损失能力及环境适应能力 [24];与植物的Pn高度相关,一般认为光合速率高,Tr也较高,因为蒸腾作用过程不仅为光合作用过程提供光合底物——水,还为光合作用过程积累物的运输提供动力 [25]。白木通老叶和新叶在15:00的Tr和Pn均达到了较高水平,呈显著正相关关系,这与前人的研究结果 [26-27]一致。关于Pn与Ci的关系前人研究得较多,本研究发现Pn与Ci呈负相关关系,主要是因为植物在高净光合速率的状态下,叶片内部的光合生理活性及相关酶的活性较强,通过气孔进入的CO2同化较快,使得Ci降低,这与李朝阳等在槲蕨中的研究结果 [28]一致。
本研究主要测定了白木通光合特性的日变化及净光合速率对光合有效辐射及CO2的响应,同时也比较了白木通不同叶龄的光合特性差异,通过研究初步了解白木通的光合特性,为白木通的人工栽培驯化提供科学依据。本研究选取的材料为白木通的盆栽苗,对大田栽培的植株及该属植物其他物种的光合特性还须进一步研究。
参考文献:
[1] 国家药典委员会 中华人民共和国药典:一部[M] 北京:化学工业出版社,2005
[2]蒋 岩,杜研学,熊 华,等 白木通籽油的理化特性及热氧化稳定性[J] 食品科学,2011,32(15):34-38
[3]高慧敏,王智民 白木通中一个新的三萜皂苷类化合物[J] 药学学报,2006,41(9):835-839
[4]柳 方 关木通与白木通的来源与鉴别[J] 新疆中医药,2004,22(2):35-37
[5]褚建民,孟 平,张劲松,等 土壤水分胁迫对欧李幼苗光合及叶绿素荧光特性的影响[J] 林业科学研究,2008,21(3):295-300
[6]李 泽,谭晓风,袁 军,等 4个油桐品种光合特性的日变化研究[J] 中国农学通报,2013,29(25):12-15
[7]刘金花,李 佳,张永清 黄芩植株光合特性初步研究[J] 中草药,2009,40(6):961-964
[8]刘 娟,梁军生,王健敏,等 思茅松干季光合生理日动态及光响应特征分析[J] 林业科学研究,2009,22(5):677-682
[9]叶子飘,于 强 光合作用光响应模型的比较[J] 植物生态学报,2008,32(6):1356-1361
[10] 瞿显友,孙年喜,李隆云,等 不同种质资源黄连净光合速率的研究[J] 中药材,2011,34(3):336-339
[11]孙 磊,章 铁,李宏开,等 柿树光合速率日变化及其影响因子的研究[J] 北方果树,2006(2):4-6
[12]萧浪涛,王三根 植物生理学[M] 北京:中国农业出版社,2004
[13]原向阳,郭平毅,张丽光,等 草甘膦对大豆光合特性日变化的影响[J] 山西农业大学学报:自然科学版,2010,30(6):500-504
[14]范彩霞,刘桂华,周 敏,等 3种阔叶树幼树光合特性的比较研究[J] 安徽农业大学学报,2010,37(2):324-327
[15] 罗爱花,郭贤仕,柴守玺 水分胁迫对不同品种春小麦的水分利用率及生理指标的影响[J] 干旱区研究,2008,25(5):683-689
[16]曹生奎,冯 起,司建华,等 植物叶片水分利用效率研究综述[J] 生态学报,2009,29(7):3882-3892
[17]朱巧玲,冷佳奕,叶庆生 黑毛石斛和长距石斛的光合特性[J] 植物学报,2013,48(2):151-159
[18]叶子飘,高 峻 丹参羧化效率在其CO2补偿点附近的变化[J] 西北农林科技大学学报:自然科学版,2008,36(5):160-164
[19]王满莲,唐 辉,孔德鑫,等 红根草光合特性的初步研究[J] 中药材,2012,35(2):179-182
[20]李 超,董希斌,李芝茹 大兴安岭低质林不同皆伐改造后枯落物持水性能分析[J] 东北林业大学学报,2012,40(4):92-96
[21]高辉远,邹 琦,程炳嵩 大豆光合日变化过程中气孔限制和非气孔限制的研究[J] 西北植物学报,1993,13(2):96-102
[22]郑国生,王 焘 田间冬小麦叶片光合午休过程中的非气孔限制[J] 应用生态学报,2001,12(5):799-800
[23]任建武,王 雁,彭镇华 3种温室栽培石斛冬季光合特性研究[J] 西北林学院学报,2009,24(1):39-43
[24]高 柱,王小玲,余发新,等 干热河谷牛角瓜光合特性[J] 东北林业大学学报,2014,42(3):19-23
[25]李 勇,韩海荣,康峰峰,等 抚育间伐对油松人工林光合作用的影响[J] 东北林业大学学报,2014,42(2):1-6
[26]栾金花 干旱胁迫下三江平原湿地毛苔草光合作用日变化特性研究[J] 湿地科学,2008,6(2):223-228
[27]杨江山,种培芳,常永义 樱桃叶片的光合特性研究[J] 甘肃农业大学学报,2006,41(4):49-51
[28]李朝阳,周 羲,陈 军,等 槲蕨的光合特性与环境因子的关系[J] 中药材,2012,35(3):347-351
