单侧后牙缺失对大鼠髁突软骨中蛋白激酶C-δ和caspase-12表达的影响
2020-10-09谢丽丽
程 洁 张 栋 谢丽丽 魏 强 康 林
颞颌关节紊乱综合征是口腔科临床的常见病、多发病,其致病机理尚未完全清楚。颞颌关节紊乱综合征的病因复杂多样,其中单侧后牙缺失、牙体牙髓病等导致的偏侧咀嚼是临床常见引起颞颌关节紊乱病的原因之一。咬合异常导致的双侧髁突受力不均,外界压力变化使得髁突软骨细胞发生氧化应激反应,引起细胞凋亡程序的启动,凋亡信号的转导途径具有多样性,使凋亡过程得到精确的调控。近年来对外界压力刺激导致的髁突软骨细胞的凋亡机制的研究多关注于细胞的基本转导通路[1~3],而对非专一性转导通路的研究较少。本研究通过造模单侧后牙缺失大鼠,检测凋亡蛋白caspase-12和蛋白激酶C-δ的变化,观察蛋白质激酶C-δ(protein kinase C-δ,PKC-δ)在髁突软骨细胞凋亡非专一性转导通路中的作用及其与基本转导通路的关系。
资料和方法
1.动物分组与取材:3周龄雄性SD大鼠(河北医科大学动物中心提供)96只,体重160~190g,适应性饲养一周后,随机等量分为8组,实验组及对照组各4组,每组12只。实验组大鼠经10%水合氯醛4ml/kg腹腔注射麻醉后,拔除左侧上、下颌所有磨牙,拔牙后第 1d、7d、14d、28d,每组随机选取 6 只,完整取出双侧髁突关节软骨,于多聚甲醛中固定。另外6只,麻醉后直接取髁突软骨冻存于-80℃冰箱中保存。对照组大鼠不做任何处理,与对应实验组同期处死并取材。
2.苏木精-伊红染色(HE染色):组织块固定后,常规石蜡包埋,4μm切片。常规脱水,透明,封片。
3.实时定量PCR(RT-PCR)检测:髁突软骨于-80℃保存,用Trizol一步提取法提取总RNA。取5μl RNA用1%琼脂糖凝胶进行电泳,以检测RNA的完整性。用TIANScript RT KIT进行反转录为cDNA,实验操作按产品说明书进行。设计引物序列进行扩增,β-actin用作内定参考。PCR反应程序:95℃15min,95℃10s,58℃30s,72℃30s,循环 40 次。分别用目的基因引物和内参基因引物进行扩增。同时在60~95℃进行溶解曲线分析。
用荧光定量PCR仪,采用2-△△CT法进行数据的相对定量分析。根据RealTimePCR原始检测结果,按照 2-△△ct相对定量计算公式,即:△△Ct=[Ct目的基因(实验组)- CtBeta基因(实验组)]-[Ct目的基因(对照组)-CtBeta基因(对照组)]。计算出各样品的目的基因相对定量结果,即其他各个样品相对于对照样品目的基因mRNA转录水平的差异。
4.统计方法:采用SPSS 19.0统计软件对实验数据进行单因素方差分析,P<0.05为差异有统计学意义。比较不同时间点实验组左、右侧及对照组caspase-12和PKC-δ在髁突软骨中的表达情况,以及各组内同侧髁突软骨不同时间点的变化。

图1 HE染色( ×200)

图2 HE染色( ×200)
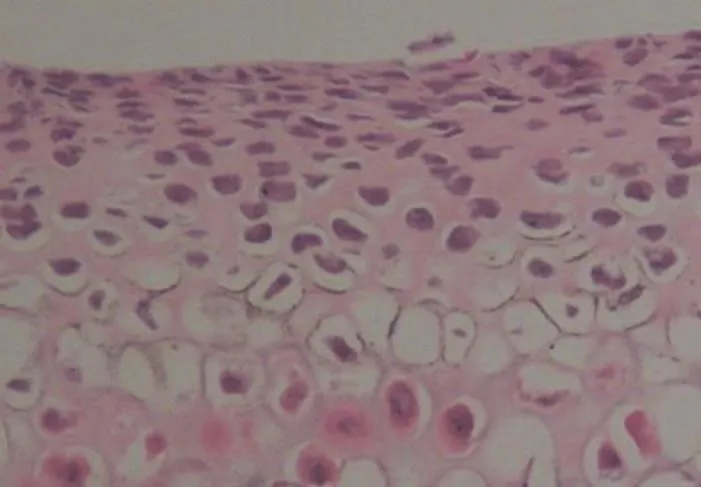
图3 HE染色( ×200)
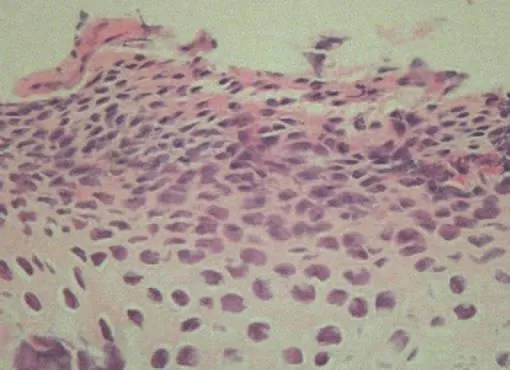
图4 HE染色( ×200)
结 果
1.HE染色:大鼠髁突软骨由表及里依次是纤维层,增殖层,肥大层及钙化软骨层。对照组中各层细胞界限清晰,排列整齐,未见退行性改变(图1)。实验组左侧7天时出现细胞排列紊乱,增殖层与肥大层界限开始模糊(图2)。实验组左侧14天时肥大层与软骨层细胞排列稀疏,可见胞核固缩(图3)。实验组左侧28天时出现细胞退行性变化,表层纤维断裂松解,软骨细胞可见核内空泡性变(图4)。

表1 拔牙后PKC-δ含量变化

表2 拔牙后caspase-12含量变化
2.RT-PCR结果:对照组PKC-δ和caspase-12的含量在不同时间点的左右侧对比均无统计学差异(P>0.05),因此将对照组不同时间点的左、右侧合并为一组。对照组各时间点的PKC-δ和caspase-12含量均无显著性差异(P>0.05)。实验组左侧PKC-δ的含量在1d、7d、14d、28d时均高于对照组,且差异有统计学意义(P<0.05),实验组右侧PKC-δ含量各时间点均高于对照组,1d、7d组有显著性差异(P<0.05),实验组左侧PKC-δ各时间点均高于右侧,7d、14d、28d组有显著性差异(P<0.05)。随时间延长,实验组左侧PKC-δ含量逐渐升高,且各时间点较前一时间点增加有显著性差异(P<0.05);实验组右侧PKC-δ逐渐降低,7d、14d组较前一时间点降低有显著性差异(P<0.05),14d、28d组与对照组无统计学差异(P>0.05)(表1)。实验组左侧caspase-12含量各时间点均高于对照组,且差异有统计学意义(P<0.05),实验组右侧caspase-12含量各时间点均高于对照组,1d,7d,14d组有显著性差异(P<0.05)。随时间延长,实验组左侧caspase-12含量逐渐升高,14d,28d时升高有统计学差异(P<0.05),而实验组右侧caspase-12含量逐渐降低,在7d,28d时降低有统计学差异(P<0.05),28d时与对照组无统计学差异(P>0.05)(表2)。
讨 论
髁突软骨细胞的凋亡途径中的凋亡蛋白酶caspases家族是重要的调控因子。caspase-12是细胞基本凋亡通路中内质网介导的凋亡通路的重要调控因子[2,4]。蛋白激酶 C(PKC)由一条单肽链组成,静止细胞中PKC主要存在于胞浆中,细胞受到刺激后,PKC通过Ca2+依赖的形式从胞浆转移到胞膜上,这一过程称为PKC的转位激活[5]。PKC是非专一性转导通路中磷脂酶C-蛋白质激酶C途径的标志蛋白。非专一性转导通路和基本转导通路实质是上下游的关系,因为前者往往需要通过激活后者才能诱导凋亡。PKC各亚型的作用也不同,如PKC-α、PKC-ε等与抗细胞凋亡关联更为密切,而PKC-δ、PKC-μ、PKC-θ 与促凋亡相关[6]。PKC-δ属于新型PKC亚家族,一般在胞浆中处于无活性状态,当受到外界凋亡信号刺激后,PKC-δ被二酰甘油(DAG)/佛波醇酯类激活,具有促进细胞凋亡的作用。PKC-δ的活化处于凋亡途径的早期,经过进一步的级联反应,激活caspase-12,最终激活caspase-3,实现细胞凋亡。
本研究中实验组大鼠拔除左侧后牙后,左侧颞颌关节髁突软骨细胞中的caspase-12和PKC-δ含量均较对照组增加,且随时间延长呈上升趋势,说明左侧髁突软骨细胞受到拔牙后的炎症刺激及正常咀嚼压力的缺失产生了氧化应激反应。当细胞传导胞外信号时,G蛋白偶联受体接受细胞外第一信使,受体被活化,进而激活G蛋白,G蛋白可进一步激活磷脂酶C,进而产生DAG,DAG与PKC-δ结合,使PKC-δ发生构象变化,产生转位激活,激活的PKC-δ转移至胞膜等亚细胞结构,磷酸化相应的底物,进一步激发细胞凋亡途径的下游事件基本转导通路,caspase-12含量的升高说明内质网介导的凋亡通路激活,而内质网介导的凋亡反应是三种基本转导通路之一,caspase-12进一步激活caspase-9,最终激活caspase-3执行细胞凋亡。因此PKC-δ和caspase-12的含量升高相辅相成。实验组大鼠右侧caspase-12和PKC-δ含量高于对照组,但随时间延长逐渐下降,可能由于拔牙初期,大鼠的咀嚼压力全部转移至右侧,使得右侧髁突软骨受到突然增大的外界压力发生细胞凋亡反应。细胞凋亡不是病理状态下的自体损伤,而是细胞应对外界刺激主动产生的自我调控反应。随着时间延长,右侧颞颌关节逐渐适应新的咬合状态,右侧髁突软骨细胞中的caspase-12和PKC-δ含量开始下降,至28天组时与对照组无显著性差异。PKC-δ在细胞凋亡中的作用机制在肿瘤研究中较为深入[7],但其对不同类型的应答反应机制尚未完全清晰,仍需进一步研究。PKC-δ的激活与caspase家族的水解切割密切相关,凋亡初期,由PKC-δ的构象变化引起转位激活,但其激活后与各凋亡蛋白间的详细传导通路尚不完全清楚。PKC-δ促凋亡的特性在许多肿瘤药物的研究中发挥了重要作用[8~10],但在髁突软骨细胞代谢途径的研究较少,进一步明确PKC-δ的相互作用蛋白,明确其促凋亡的作用机制,对于解释单侧咀嚼导致的颞颌关节紊乱病的疾病发生发展的进程具有意义。
