镉胁迫下能源甜菜BvGST基因在酵母中的功能分析
2020-10-02杨舒涵王锦霞郭萌萌马龙彪刘大丽
杨舒涵,王锦霞,郭萌萌,马龙彪,刘大丽
(1.黑龙江省普通高等学校甜菜遗传育种重点实验室/黑龙江大学现代农业与生态环境学院,哈尔滨 150080;2.中国农业科学院北方糖料作物资源与利用重点实验室/中国农业科学院甜菜研究所,哈尔滨 150080;3.黑龙江大学生命科学学院,哈尔滨 150080)
0 引言
重金属(As、Cr、Pb、Cd等)毒害导致活性氧物质(ROS)的形成和积累,并干扰细胞的氧化还原平衡,进而严重地影响植物的生长发育[1]。作为固生生物的植物在长期的适应过程中,也相应地进化出了一套调控防御系统,并激活逆境应答机制来应对有害环境的毒害[2]。植物体内拥有高效的酶促和非酶促抗氧化防御体系,以作用于一系列不可控的氧化反应,保护植物细胞最大程度地免受毒害。在植物的抗氧化机制中,用于清除ROS 的酶类[3-4],如过氧化氢酶(CAT)、超氧化物歧化酶(SOD)、过氧化物酶(POD)等,以及解毒酶(Glu‐tathione S-transferases,GSTs),在直接或间接地保护细胞免受逆境伤害中发挥着重要作用。
Glutathione S-transferases(GSTs,EC 2.5.1.18)作为多功能蛋白之一,由一类高度多样性的古老的基因家族编码,并广泛参与到植物生长发育、细胞内代谢、逆境耐受、外源毒素和内生化合物的解毒过程中。它通过与谷胱甘肽(Glutathione,GSH)共价结合形成多种多样的三肽疏水亲电底物[5],从而络合活性氧物质,调控细胞氧化还原状态,生物合成、结合和转运次生代谢产物及激素等[6-7]。通过对包括拟南芥、杨树和水稻在内的植物基因组研究发现,植物中的GSTs拥有一个超过55个成员的庞大的基因家族[8-10]。根据氨基酸一致性、基因结构和底物特异性,GSTs被分为八大类:Tau、Phi、Lambda、Theta、Zeta、Dehydroascorbate reductase(DHAR)、Elongation factor 1 gamma(EF1Bγ)以及Tetrachlorohydroquinone dehalogenase(TCHQD)[8]。其中,Tau和phi是液泡植物中最丰富的两种类型的GSTs,并拥有广泛的底物特异性[11]。由于它们属于酶活性蛋白,大量的研究集中在结合基因组学结构,基因表达以及酶促分析等阐述重复基因的保留和功能分化的功能机制方面[12]。
除此之外,研究还发现,植物GSTs参与众多细胞过程,作用于过氧化氢的解毒、花青苷的转运、除草剂解毒、生长素代谢平衡、酪氨酸代谢以及程序性细胞凋亡的调控等由于生物及非生物逆境带来的响应[13],从而保护植物免遭包括重金属逆境在内的各种非生物逆境的伤害[14-15]。研究表明,拟南芥AtGSTF2 基因与防御相关化合物的结合和转运调控有关[16],过量表达AtGSTF2 基因可以提高植物对苯酚逆境的耐受性[17]。重金属As 逆境会导致由于GST 和GPX 基因的诱导表达而带来的GSH 含量的增加,从而大量合成植物螯合素以解毒活性氧物质[18]。除此之外,研究表明,拟南芥Phi 家族的GSTs 可以被Cu 和Al 诱导表达[19];Trichoderma virens GST(TvGST)可以提高真菌对Cd的耐受性[20]。
虽然GSTs 被证实与重金属逆境耐受相关,然而对于在重金属尤其是镉逆境下GSTs 精确的生理作用和分子机制还不清楚。甜菜是新兴的能源作物,在重金属污染土壤的生物修复中表现出了极大的潜力[21],因此,了解其重金属耐受作用机理十分重要。结合前期的相关研究[21-23],我们在对能源甜菜重金属镉逆境的转录组表达谱的研究中,发现BvGST基因的差异表达十分显著,并通过qRT-PCR 进一步确定了该基因与Cd的耐受应答相关。本研究克隆了BvGST 基因,并利用单细胞酵母重组构建及表达该基因,通过比较分析该基因的表达真核酵母细胞中的作用,从而为进一步了解BvGST 在能源甜菜体内的功能及其在重金属离子代谢平衡中的分子机制奠定基础。
1 材料与方法
1.1 目的基因的克隆
根据甜菜Beta vulgaris glutathione S-transferase(BvGST,LOC104898671)基因的碱基序列以及酵母表达载体pYES2 的多克隆位点设计引物:[Forward primer (5′-3′):CGGGA TCC(BamHI) ATG GGA ATC AAG ATT CAT GGA AT;Reverse primer(5′-3′):CCGCTC GAG(XhoI)TTA AGC TTG CTT CAA CAG AGC CAC]。以Trizol法提取的能源甜菜总RNA反转录的cDNA(First strand cDNA synthesis kit,TOYOBO)为模板,利用KOD-Plus-Neo 高保真酶(TOYOBO)扩增获得目的基因全长。将PCR 产物于25 ℃连接到克隆载体pEASY-T1(Trans‐Gen)上,并转化到Trans1-T1 感受态细胞中。提取KanaR阳性克隆质粒DNA,并利用BamHI和XhoI对重组质粒进行限制性内切酶酶切鉴定。所获得的阳性重组质粒进一步送往上海生工测序,确定其碱基序列的正确性和完整性。
1.2 酵母表达载体的构建
利用BamHI 和XhoI分别双酶切pEASY-T1-BvGST和pYES2质粒。经过琼脂糖凝胶电泳以及胶回收,利用T4 ligase(NEB)将回收载体和目的片段按照1∶7 的摩尔比于16 ℃连接过夜。连接产物转化到Trans1-T1大肠杆菌感受态细胞中,提取AmpR克隆质粒DNA,利用BamH I 和Xho I鉴定pYES2-BvGST的正确性。重组酵母表达载体转化到酵母InVSc1中备用。
1.3 酵母RNA的提取
将含有pYES2-BvGST的酵母菌培养于YPGDR 培养基中,从OD600=0.5初始诱导培养0 h、4 h、8 h、12 h和24 h,收集菌液。加入等体积的Tris 饱和酚以及Lysis buffer(10 mmol/L Tris-HCl pH7.4,10 mmol/L EDTA,0.5%SDS)充分重悬细胞,65 ℃孵育45 min。经过酚、氯仿反复抽提上层清液,并利用3 mol/L NaAc(pH5.2)以及无水乙醇沉淀获得酵母RNA。利用紫外分光光度计NavoVue plus在OD260和OD280下检测RNA质量。
1.4 半定量RT-PCR
以定量后的酵母RNA 反转录的cDNA 为模板,进行PCR 扩增。其中,内参Saccharomyces cerevisiae ACT1基因(ACT1,YFL039C)的扩增引物为:Forward primer (5′-3′):TCATGGTCGGTATGGGTCAA;Reverse primer(5′-3′):TCAGCAGTGGTGGAGAAAGA。Tm=58 ℃;扩增片段长度为487 bp。取20µL PCR 产物,于1.0%的琼脂糖凝胶上进行电泳检测。
1.5 镉胁迫下酵母菌生长曲线的测定
将转入pYES2 或pYES2-BvGST 的InVSc1 菌株培养于SC-U(6.7 g/L yeast nitrogen base、200 mg/L-Uracil DO supplement和2%glucose)培养基直到OD600=0.5。收集菌液并培养于含有0.5 mmol/L CdCl2的YPGDR(1%yeast extract、2%peptone、1.94% galactose、0.06%glucose 和1%raffinose)液体培养基中,30 ℃,每隔一定时间间隔,于OD600测一次菌液浓度,制作生长曲线。实验分别进行3次重复。
2 结果与分析
2.1 能源甜菜BvGST基因的克隆
根据能源甜菜Beta vulgaris glutathione S-transferase(BvGST,LOC104898671)基因的碱基序列,在该基因起始密码子和终止密码子两端设计分别带有BamHI和XhoI酶切位点的特异引物,以能源甜菜总RNA反转录的cDNA 为模板扩增该目的基因。如图1-A 所示,经过RT-PCR 及琼脂糖凝胶检测,利用高保真Taq 酶扩增BvGST基因,在642 bp 左右的位置获得了目的条带。此PCR产物与目的基因在Genbank里的序列大小一致,无杂带,可以直接用于下一步的载体构建。
将BvGST基因的PCR扩增产物连接到克隆载体pEASY-T1上。如图1-B所示,经过E.coli转化、Kana抗性筛选阳性克隆、质粒提取以及BamHI、XhoI 限制性内切酶酶切鉴定,实验分别在3.928 kb 和642 bp 左右的位置上,获得了相对应于载体pEASY-T1 和目的基因片段BvGST 的条带。将阳性重组质粒测序,通过Blast碱基序列比对,最终确定了pEASY-T1-BvGST克隆的目的基因的正确性。

图1 能源甜菜BvGST基因的克隆Fig.1 Cloning of energy beet BvGST gene
2.2 酵母pYES2-BvGST表达载体的构建
提取酵母pYES2质粒,利用限制性内切酶BamHI和XhoI将pYES2质粒和pEASY-T1-BvGST质粒进行酶切,得到带有特异性连接的粘性末端的载体和目的片段(图2-A)。胶回收目的条带,通过T4 连接酶过夜连接以及大肠杆菌转化,所获得的的Amp 抗性克隆的质粒DNA 经过BamHI 以及XhoI 的双酶切鉴定,分别在5.9 kb和642 bp 左右的位置获得了相应的目的条带,从而获得正确的酵母重组质粒pYES2-BvGST(图2-B)。将阳性克隆转入酵母InVSc1中以进行下一步的检测及功能分析。

图2 pYES2-BvGST重组质粒的鉴定Fig.2 Identification of recombinant plasmid of pYES2-BvGST
2.3 BvGST基因在酵母体内的表达
为了进一步检测重组质粒pYES2-BvGST在酵母InVSc1中的表达情况,实验将重组菌培养于诱导型培养基YPGDR 中,适量诱导表达目的基因BvGST,以酵母ACT1 基因作为内参基因。Semi-quantitative PCR 研究表明,经过半乳糖的诱导,随着培养时间的增加,与酵母ACT1基因的表达几乎没有变化相比,BvGST 基因的表达量却在逐渐增加(图3),从而实验进一步证实了甜菜BvGST 基因在重组酵母菌体内可以被诱导并正确地表达。

图3 BvGST基因在酵母中的诱导表达Fig.3 Induction and expression of BvGST gene in yeast cells
2.4 镉胁迫下甜菜BvGST基因在酵母体内的功能分析
为了分析在重金属镉逆境下,BvGST 基因在酵母菌体内的功能,实验以转化pYES2 空载体的酵母为对照,通过半乳糖以及葡萄糖的诱导抑制组合,分析目的基因的表达是否可以提高酵母对Cd的耐受性。如图4所示,在正常生长条件下,即没有重金属逆境胁迫的条件下,无论是YPD 抑制表达培养基还是在YPGDR 诱导培养基中生长的两组酵母的生长速率几乎持平,说明BvGST 基因的表达对酵母菌的生长没有明显的影响。与之相反的是,在Cd逆境胁迫下,无论是对照还是含有目的基因的重组菌,在两种培养基中的生长均受到了抑制,生长缓慢并在一定阶段伴随着下降的趋势;但在YPGDR 培养基中,适量表达BvGST基因的酵母的生长趋势要明显优于对照pYES2酵母菌。这些数据表明,能源甜菜BvGST在酵母内的表达在镉逆境胁迫下的生长和耐受性的提高中发挥着重要作用。
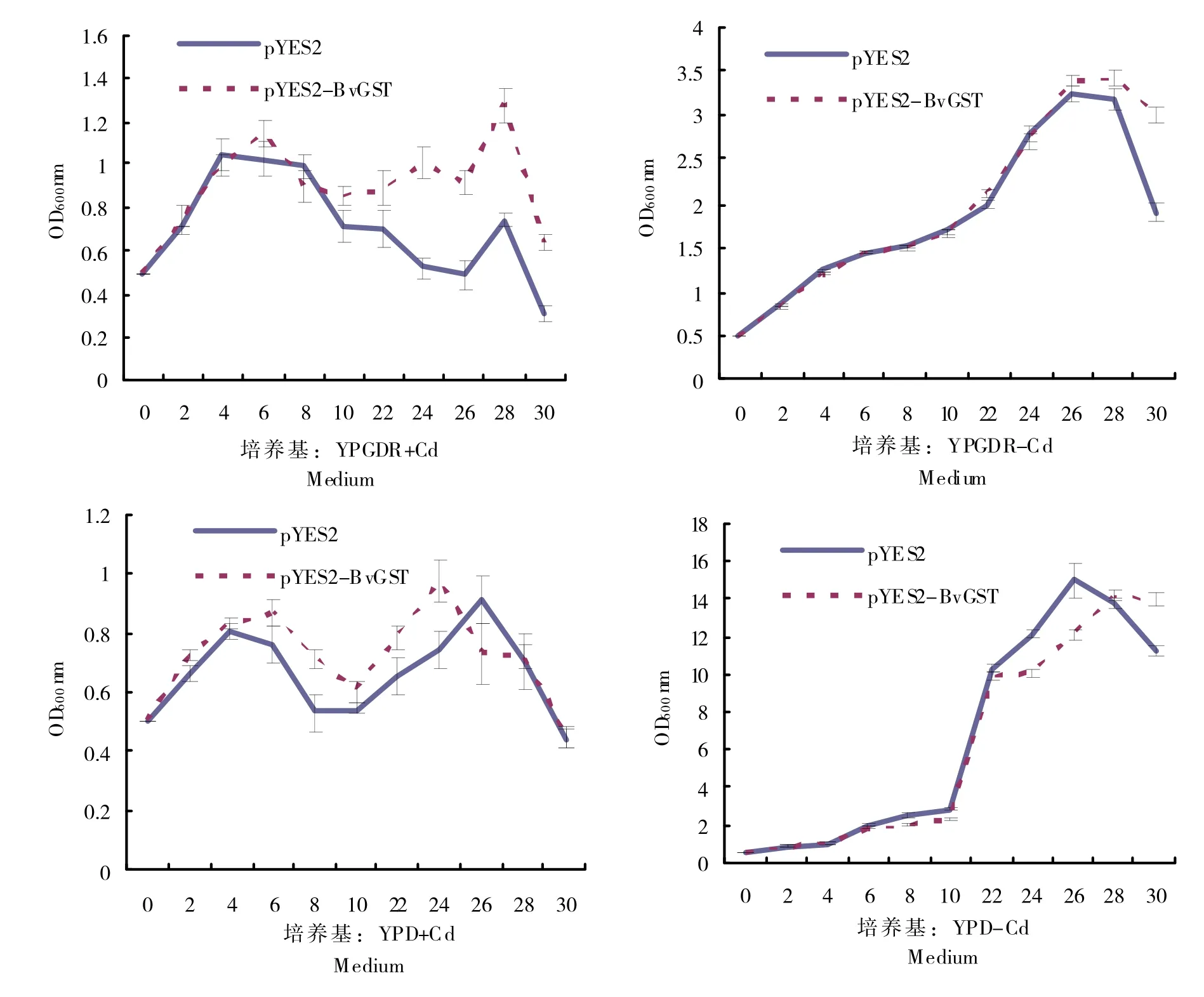
图4 镉胁迫下重组酵母菌的生长曲线Fig.4 Growth curve of recombinant yeast under cadmium stress
3 讨论
在重金属逆境胁迫以及相关的生物修复研究中,镉被认为是一类高毒性的离子,它可以通过植物根部吸收并通过输导组织被运送到其它部位的细胞、组织及器官中,引发次级逆境-氧化逆境的发生,进而直接或间接对植物产生毒害作用。而在谷胱甘肽作为二硫还原剂,介导调控由于重金属逆境所引起的基因表达中,谷胱甘肽转移酶Glutathione S-transferases(GSTs)扮演着重要的角色[24]。除此之外,植物GSTs 在对生长素的细胞应答以及植物次生产物的代谢过程发挥着重要作用。可见,了解BvGST 基因的作用机制和分子功能,对于解读能源甜菜在Cd逆境下的基因应答调控机理十分重要。
前期的研究发现,重金属逆境会对能源甜菜的生长造成一定程度的影响[21]。通过转录组学的分析发现,在众多基因的胁迫应答过程中,能源甜菜BvGST基因的差异表达极其显著。因此,本研究利用RT-PCR 技术克隆了该基因,并将其重组表达于酵母这一真核单细胞生物体内以研究其作用机制。通过半乳糖以及葡萄糖组合的适量诱导表达目的基因,发现该基因在重金属镉逆境胁迫下,可以在一定程度上提高酵母菌的生长状态和对Cd离子的耐受性。
在以往的报道中发现,水稻OsGSTU30和OsGSTU41基因的大量表达在不影响酵母在正常生长条件下的生长状态的情况下,可以通过较高的GST 活性提高菌体对重金属Cr(VI)的抗性[25]。因此,研究结果说明了BvGST 是能源甜菜体内重要的解毒酶之一,通过其在酵母体内与Cd 逆境胁迫中的生长调控机制的研究,确定其在重金属逆境功能机制中发挥着重要作用。同时,该基因的克隆也为下一步深入细致地研究其在重金属逆境胁迫下的组织表达特性和缺失突变体的获得奠定基础。
4 结论
本研究通过克隆和酵母菌体外重组表达目的基因BvGST,发现该基因可以在一定程度上提高宿主菌在重金属Cd逆境下的生存状态和生长趋势。能源甜菜BvGST基因与镉逆境胁迫之间存在着一定的应答关系,该基因可能通过直接或间接的作用降低Cd对细胞的毒害,从而提高酵母对重金属的耐受性。研究结果也为进一步对BvGST基因的功能开发和利用生物技术改造人工超累积能源甜菜奠定基础。
