沙柳实际光化学效率与光合有效辐射日变化动态及其对环境因子的响应
2020-09-27梁丽壮李晓燕胡雪
梁丽壮 李晓燕 胡雪
(北京林丰源生态环境规划设计院有限公司 北京海淀100083)
干旱半干旱区约占地球陆地面积的40%,同时提供了40%的陆地净初级生产力[1-2]。在全球气候不断恶化的背景下[3],诸如水分亏缺、极端温度、高太阳辐射等极端天气在该区出现的频率和程度持续增加[4-6]。在多种环境胁迫下,该地区荒漠植被经常表现出明显的受胁特征[7-9],并且脆弱的生态系统也面临着急剧退化的风险[10]。因此,研究自然条件下荒漠植物对环境胁迫的适应规律和调控机理至关重要[11],探寻各种途径和方法去捕捉并衡量荒漠植物对环境胁迫的响应也是必要的,进而可精确地捕捉并预测荒漠植物对目前和未来可能的极端气候变化的响应[12]。
光合作用是植物细胞中最重要的吸能反应,不同强度和不同持续时间的多重胁迫会抑制整个光合过程[13],光系统II(PSII)光化学被认为是光合作用中对胁迫最敏感的部分[14],光合过程中PSII 所吸收的光能会有约5%以叶绿素荧光的形式被重新释放出来[15]。叶绿素荧光与光合有紧密的联系,因此叶绿素荧光技术也被视为是研究植物光合作用与环境关系的内在探针[16]。通过监测植物叶片叶绿素荧光的变化,我们可以快速、无损的测定光合过程中光能的吸收、传递、耗散、分配等,以此了解受PSII 胁迫程度大小以及光合器官是否被破坏等信息,如通过暗适应后的叶片由光照引起的荧光变化来评估植物的光合生理状态[17-18]。近年来,叶绿素荧光技术监测植被对环境因素的响应已逐渐在环境胁迫对光合的影响方面得到应用[19-20],有关该技术的研究也越来越受到人们的关注。脉冲振幅调制荧光测量,是指通过饱和脉冲使叶绿素荧光快速地处于某些特定状态,并瞬时进行测量的技术[21],其已成为研究植物光合作用最有力工具之一,并逐渐地应用于野外现场原位监测[8,22-23]。实际光化学效率(ФPSII)和最大光化学效率(Fv/Fm)都是通过脉冲振幅调制荧光测量计算得到的参数[21],ФPSII反映了光合作用中PSII的电子捕获能力,Fv/Fm表示植物的潜在最大光合能力,两者都是反映植物光合性能和健康状况的重要参数[16-17]。计算Fv/Fm需要最小荧光(Fo)和最大荧光(Fm),通过暗适应后PSII 反应中心完全开放测得Fo,后作用于饱和脉冲光下,此时所有电子通道关闭,光合作用被暂时抑制,叶绿素荧光值达到最大,测得Fm[18]。然而,这种对黑暗的苛刻要求是在白天实地测量很难实现的,有关研究提出ФPSII对光合有效辐射(PAR)(ФPSII-PAR)的日变化关系可以很好地捕捉植物对环境变化的响应,ФPSII-PAR回归斜率和截距(ФPSII在PAR= 0 时的拟合值)可以作为评估植物的光合生理状态的有效指标[24]。在这种基于日变化的关系中,ФPSII-PAR回归斜率反映了光合吸收光能的变化,过剩光能以热的形式消散[15],斜率绝对值的增大表明植物对光能的耗散能力增加,因此,斜率可以用来反映植物PSII 在胁迫条件下的热耗散敏感性。另外,ФPSII-PAR回归截距可以很好的代替Fv/Fm(作为当天Fv/Fm的估值)[25-26]。由于ФPSII-PAR日变化关系所包含的信息量远远超出单次测量的荧光参数,所以其数值方面波动也更敏感,反映结果也更趋近于真实[24]。目前关于ФPSII-PAR日变化关系的研究较少,并且这种日变化关系能否有效地揭示干旱地区环境胁迫对荒漠植物光合的影响我们并不明确,因此还需要进一步探究了解。
以荒漠灌木沙柳为对象,采用叶绿素荧光原位连续监测技术进行了整个生长季的观测,并同步观测了环境因子,分析比较了ФPSII、Fv/Fm和ФPSII-PAR日变化关系的动态及其和环境因子的关系,验证了ФPSII-PAR日变化关系的回归斜率和截距可作为反映荒漠植物PSII 对环境波动响应的适合指标,进而为区域尺度研究和荒漠生态系统过程建模提供重要的光合生理生态参数,为环境胁迫如何影响荒漠灌木PSII 并对其造成损伤提供理论依据。
1 材料与方法
1.1 研究区概况
研究选在宁夏盐池荒漠生态系统国家定位观测研究站点(37°53′83″N,107°25′46″E,海拔1550 m)进行,盐池县地处宁夏东部,北接毛乌素沙地,东临黄土高原,是半干旱区向干旱区过渡的典型生态交错带。该区抗干扰能力差,易受外界条件影响,是沙漠化敏感地带,属中温带大陆性季风气候,年平均气温8.1℃,年内降雨不均,蒸发强烈,以风沙土为主,植被主要包括沙柳、杨柴(Hedysarum mongolicum)、油蒿(Artemisia ordosica)和花棒(Hedysarum scoparium)等荒漠灌木。
1.2 叶绿素荧光的观测
于2013 年5-10 月使用多通道连续叶绿素荧光监测仪(MONI-PAM,Walz,Germany)对试验站样地内多年生天然沙柳群落进行自然状态下的长期原位连续监测。在长势良好的植株上选择粗细适中枝条的健康成熟叶片,用固定荧光探头(MONI-head)叶夹夹取2 至3 片固定,调整探头位置使叶片保持充分受光,定期检查并适时更换。监测参数包括实时荧光值(Ft)和光下最大荧光值(Fm′),测定间隔为30min。
1.3 环境因子的观测
环境因子PAR由荧光监测仪实时同步测量,空气温度(Ta)和相对湿度(RH)由距荧光监测仪200 m、高度为6 m 通量塔装配有的温湿度传感器(HMP155A, Vaisala, Vantaa,Finland)测量,饱和水汽压差(VPD)由Ta和RH计算求得(VPD= 0.611e17.27Ta/(Ta+237.3)(1-RH/100))。土壤含水量(SWC)通过在荧光监测仪旁深度为30 cm 的ECH2O-5TE 土壤温湿度传感器测定(Decagon Devices, Pullman,WA,USA)。降雨量(PPT)由距荧光监测仪400m 处空地内高1.8 m 的TE525W 翻斗式雨量筒(Campbell Scientific Inc.,Logan,UT,USA)测定,环境因子观测间隔为30min。
1.4 数据处理与分析
使用数据采集器的WinControl-3 软件对原始荧光数据进行批量处理,获得荧光参数的时间序列。用极限检查筛选法,将低于100 的Ft和Fm′的异常值从数据集中删除。ΦPSII通过公式计算求得(ΦPSII=(Fm′-Ft)/Fm′),将夜晚Ft和Fm′分别看作Fo和Fm,计算最大光化学效率(Fv/Fm=(Fm-Fo)/Fm)。将整个生长季白天ΦPSII和PAR数据按PAR间隔100μmol·m-2·s-1分段平均,将每天白天的ΦPSII和PAR用直线拟合求得当天ΦPSII-PAR日变化回归斜率和截距,用直线拟合ФPSII与PAR和回归截距与Fv/Fm。通过皮尔逊相关性分析检验环境因子对Fv/Fm和ФPSII-PAR日变化回归斜率和截距的影响,同时运用直线、指数或多项式拟合分析了上述参数对环境波动的响应。
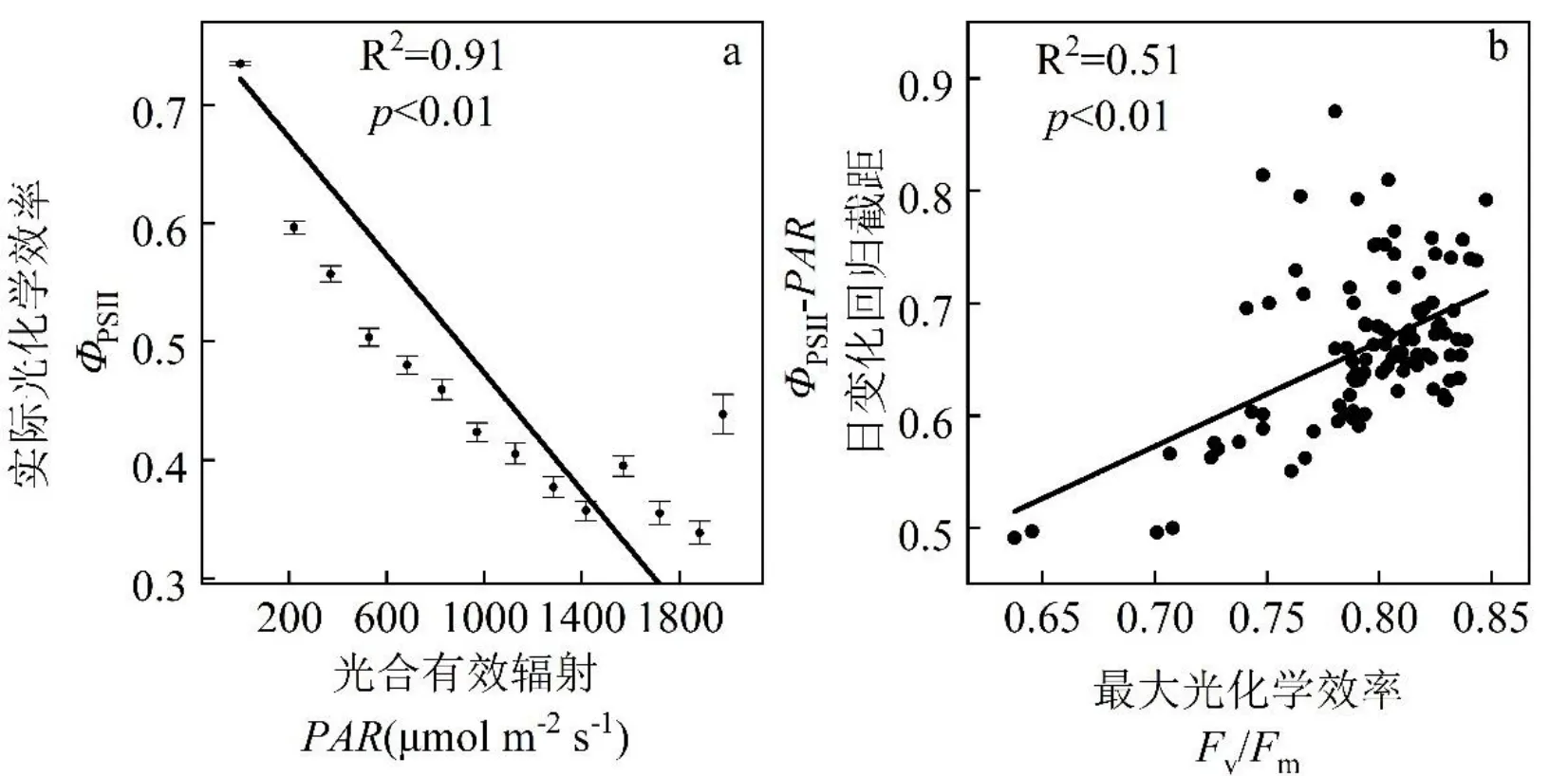
图1 实际光化学效率对光合有效辐射(ФPSII-PAR)的响应
2 结果与分析
2.1 实际光化学效率对光合有效辐射(Ф PSII-PAR)响应及日变化
由于夜晚没有光照,因此只对白天的数据进行线性拟合,观测期白天ΦPSII与实时PAR关系呈极显著线性负相关,ΦPSII随PAR增加而降低,ΦPSII-PAR直线拟合的方程为:Y=0.722-2.48×10-4X(p<0.01,R2=0.87)(图1a)。观测期ΦPSII-PAR日变化回归截距与Fv/Fm呈显著正相关,截距随Fv/Fm增加而增加,线性拟合的方程为:y=-0.08+0.92x(p<0.01,R2=0.51)(图1b)。
2.2 环境因子的动态
观测期内,PAR天平均值最高为708.5μ mol·m-2·s-1(DOY162),最低72.4μmol·m-2·s-1(DOY170);PAR天最大值(PARmax)最高1262 μmol·m-2·s-1(DOY173),最低345μmol·m-2·s-1(DOY171),PAR和PARmax在生长季内呈逐渐下降趋势(图2a)。Ta最高为32.1°C(DOY217),最低为0.6°C(DOY268),一天内最大温差17.9°C(DOY183),Ta在春季(DOY106-152)和秋季(DOY245-287)波动较大,在夏季(DOY153-244)较稳定,呈缓慢增加后降低趋势(图2b)。VPD最高为3.67 kPa(DOY179),一天内最大差值为3.41kPa(DOY217)(图2c)。观测期SWC平均为0.083 m-3·m-3,最高为0.166m-3·m-3(DOY191),最低为0.062 m-3·m-3(DOY169),降雨量总计242mm,SWC只在降雨量超过20mm 的两次降雨(DOY184,DOY190)后有相对明显的增加,存在时间滞后(图2d)。
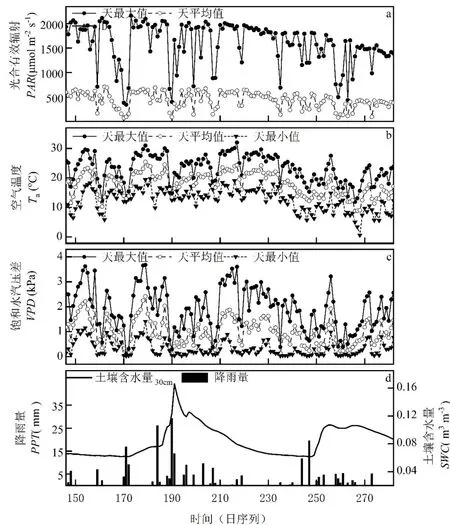
图2 环境因子的动态变化
2.3 叶绿素荧光参数和ФPSII-PAR 日变化关系的动态
观测期ΦPSII日均值平均为0.62,最大日均值为0.78(DOY265),最小为0.41(DOY147),ΦPSII在整个生长季波动较大(图3a)。Fv/Fm平均值为0.79,最高为0.84(DOY259),最低为0.63(DOY282),Fv/Fm变化趋势与ΦPSII日均值相似,在生长季前期和末期各有一次较为明显的下降(图3b)。ФPSII-PAR日变化回归斜率均值(绝对值)为2.04×10-4,最高为4.37×10-4(DOY263),最低为0.91×10-4(DOY227),在生长季末期波动明显增加(图3c)。ФPSII-PAR日变化回归截距均值为0.66,最高为0.87(DOY192),最低为0.49(DOY282),变化趋势与Fv/Fm相似(图3d)。
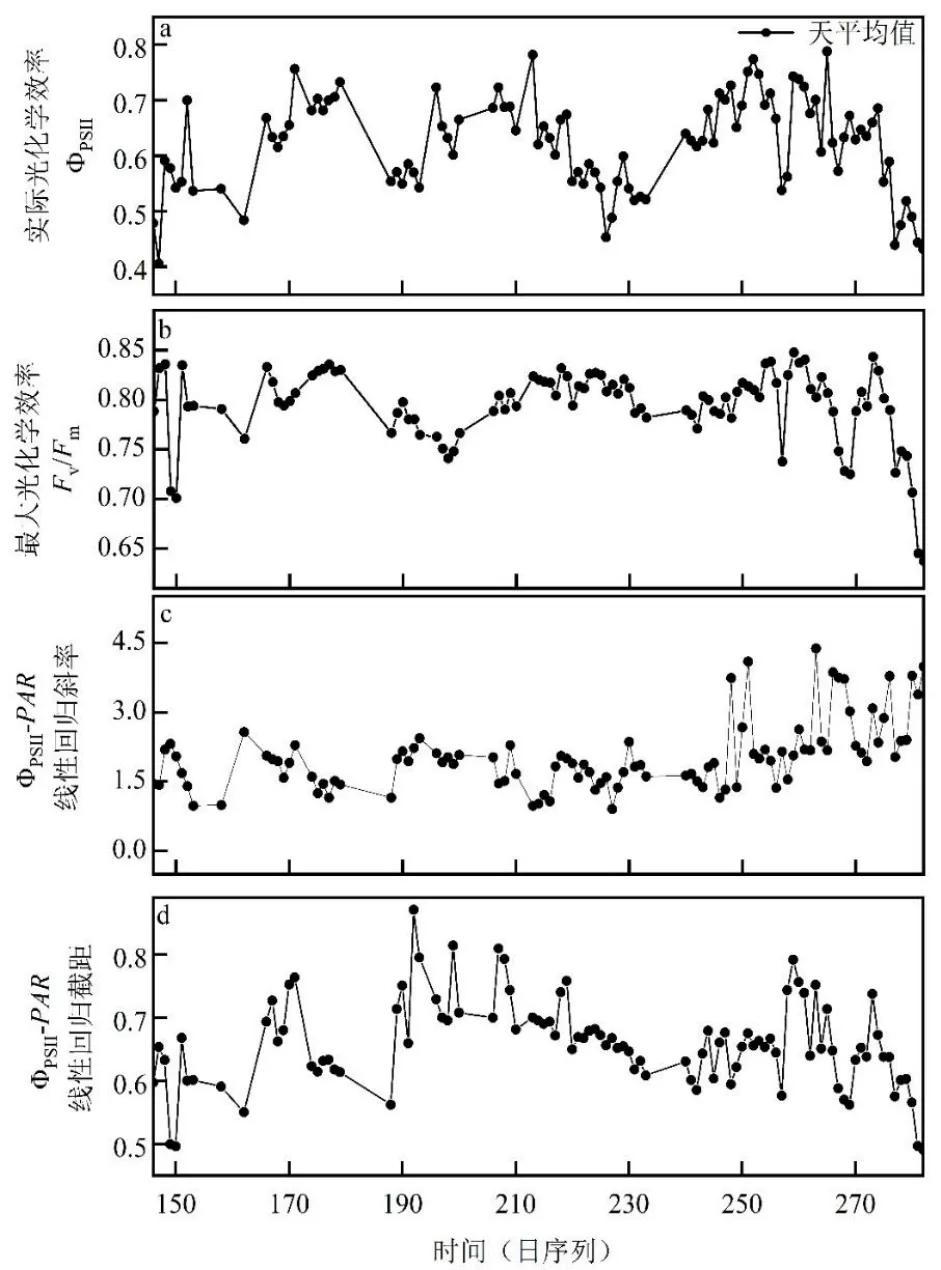
图3 叶绿素荧光参数和ФPSII-PAR 日变化关系的动态
2.4 最大光化学效率(Fv/Fm)和ФPSII-PAR 日变化关系对环境动态的响应
为探究沙柳PSII 对环境因子波动的响应,用皮尔逊相关性分析法检验了环境因子PARmax、Ta、VPD和SWC与Fv/Fm、ФPSII-PAR日变化斜率和截距的关系,同时进行了指数或线性拟合(比较R2选取最优模型)。其中,参数Fv/Fm只与PARmax和Ta显著相关(p<0.05),且无具有统计学意义的拟合模型适用。ФPSII-PAR日变化斜率和截距均与所有环境因子呈极显著相关关系(p<0.01)(表1)。ФPSII-PAR日变化斜率拟合方程为:Y=6.31-3.9×10-1Ta+7.72×10-3Ta2(p<0.01,R2=0.48),Y=2.49-2.8×10-1VPD+1.6×10-1VPD2(p<0.01,R2=0.38)。ФPSII-PAR日变化截距拟合方程为:Y=0.79-8.85×10-5PARmax(p<0.01,R2=0.31),Y=0.65×VPD-0.07(8p<0.01,R2=0.29),Y=0.59+1.18×SWC(p<0.01,R2=0.37)(图4)。
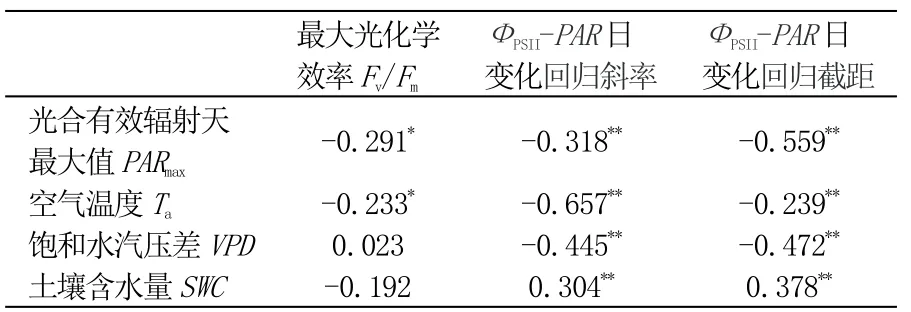
表1 叶绿素荧光参数与环境因子的相关性
3 讨论
3.1 叶绿素荧光相关参数的季节动态
光被植物叶片吸收后并不能全部用于光化学反应,一部分会以热能和叶绿素荧光的形式耗散掉,保护光合器官免受灼伤[27],因此通过叶绿素荧光可以反映光合作用的活性和行为。叶绿素荧光参数ФPSII表示了光化学反应的实际转化效率,能够确定环境变化条件下PSII反应中心的作用效率[16],研究地沙柳ФPSII波动于整个时期,在生长季初期和末期处于相对较低水平,同时在SWC低(SWC<0.08 m-3·m-3)时期也有不同程度的下降(图2d;图3a)。这与沙柳物候期和生理特性基本相符,5 月末和10 月初沙柳处于展叶期和叶片枯萎期,此时叶片未完全展开或开始枯萎,叶绿素含量与活性较低[28],光能捕获能力下降,影响了光合作用过程中电子传输速率和能量的供应[29],最终导致了ФPSII的低值。另外,高等植物叶绿体内除叶绿素外还有大量的类胡萝卜素[30],如叶黄素。其可以直接充当补光色素以保证光合作用的正常进行,同时也在防御光合器官损伤中发挥重要作用[31]。ФPSII的降低,也可能是由于该物候时期叶黄素等类胡萝卜素的缺少,影响了叶绿体类囊体膜上脱氧化和环氧化相互转化的叶黄素循环等胡萝卜素调控机制[23]。SWC低时,沙柳促使其气孔关闭抑制蒸腾作用避免水分的过度流失[32-33],但气孔导度的下降也限制了光合气体交换过程,细胞间的二氧化碳浓度无法满足光合机构的大量需求,二氧化碳代谢无法与吸收的光能同步[33]。过剩光能同时也影响了Rubisco 酶的羧化活性,电子传递受阻最终导致光抑制现象产生[34],光合效率降低达到饱和,表现为ФPSII的降低。
Fv/Fm可作为植物是否受到环境胁迫的重要指标[24],其大小基本不受物种和环境条件的影响,植物在非胁迫状态下的Fv/Fm约为0.75-0.85[24],研究认为Fv/Fm=0.44 可以作为PSII 是否受损的临界值[17,34]。沙柳Fv/Fm在观测期内波动范围在0.60-0.85 之间,远高于0.44。Fv/Fm在生长季初期和末期处于较低水平,伴随连续阴天(DOY195)以及两次气温骤降(DOY257,DOY267),Fv/Fm也出现了短期低值的情况(图2a,b;图3b)。结果表明沙柳在展叶期和叶片枯萎期,由于叶片发育不成熟或叶片老化PSII容易受到胁迫[28,35-36],环境因子PARmax的增加和SWC的降低均对沙柳PSII 造成抑制并表现出不同程度的受胁迫程度,从Fv/Fm的波动来看,沙柳基本处于相对适宜状态(Fv/Fm达到0.8 以上),可以认为高辐射低温等环境胁迫对沙柳PSII 造成的损伤具有可恢复性[24],沙柳能通过自身调节恢复到适宜状态。
3.2 ФPSII-PAR 日变化关系及其对环境波动的响应
测定Fv/Fm需要黑暗环境以保证PSII 反应中心的完全开放或关闭,这在野外实地监测中存在很大难度[18,24]。目前有研究学者在清晨黎明前测定Fv/Fm[23],因为此时PAR较低,满足测定的条件,但植物经过了一晚上的恢复,测定的Fv/Fm对植物实际胁迫的表达不可避免的存在不足之处[24]。将夜晚中连续测定的Ft和Fm′分别作为Fo和Fm,计算求得单次Fv/Fm,再将夜晚所有Fv/Fm平均后再作为当日Fv/Fm,同样存在表达不足的问题,因此导致Fv/Fm与环境因子的关系并不理想(表1;图4)。Howarth等发现一种海草泰莱草(Thalassia testudinum)白天ФPSII与PAR的线性回归截距与Fv/Fm对光照强度、盐分胁迫的响应一致[25],因此认为白天ФPSII-PAR日变化回归截距能够作为植物是否受到胁迫的指标。同时,沙柳ФPSII与PAR有很好的线性关系(R2=0.87)也证明了该研究方法可行(图1a)。另外,ФPSII-PAR日变化回归斜率表示植物耗散过剩光能的能力,反映了在胁迫下PSII 的光化学敏感性[24-26]。
从季节动态来看,沙柳ФPSII-PAR日变化回归斜率在生长季前中期(DOY146-210)趋势和Fv/Fm相似,同时处于较低水平,在生长季末期(DOY255-282)其趋势和Fv/Fm大致相反,ФPSII-PAR日变化回归斜率高值与Fv/Fm低值交替出现(图3b,c),ФPSII-PAR日变化回归截距比Fv/Fm偏低,但其波动趋势与Fv/Fm相近(图3b,d)。造成这种明显的季节差异的原因可能是沙柳在生长季前中期叶片发育不完全,叶片无法利用较多光能[28,37],使得一天内ФPSII都处于较低水平,因此斜率降低。由于新生叶片的抵抗能力较弱,PSII 易损伤且不能及时恢复[15,38],导致Fv/Fm也处于较低水平。而在生长季末期,成熟的沙柳叶片能够在短期内高效的利用光能,在高PAR或其他环境胁迫下通过提高热耗散来消耗过剩光能[39],ФPSII在一天之中大幅度变化导致回归斜率增加。同时,低ФPSII不利于叶片维持光能转化及暗反应的碳同化物的积累[24,38],伴随着低温、干旱和高辐射等环境胁迫的出现(图2a,b,d),加重了光抑制导致PSII 反应中心损伤[7,14,39-40],导致Fv/Fm低值出现。从线性回归来看,截距与Fv/Fm之间存在线性关系(R2=0.87),ФPSII-PAR日变化回归截距随Fv/Fm的增加而增加,拟合截距接近0 且斜率接近1(图1b)。从与环境因子的相关性来看,ФPSII-PAR日变化回归斜率和截距均比Fv/Fm对环境因子有着更强的关系(表1)。从对环境波动的响应来看,ФPSII-PAR日变化回归斜率随Ta和VPD的升高而降低。可认为沙柳可在Ta和VPD较低的有利条件下提高光合速率,在高温和高VPD的不利条件下,通过调节气孔减少蒸腾[32-33],防止水的过度丧失,但不可避免地抑制了光合速率。研究发现沙柳能够通过调节体内渗透产物、抗氧化酶等来缓解因缺水导致的PSII 受损来适应干旱胁迫[8,24],这一结果也是沙柳PSII 对水分亏缺长期适应的体现。ФPSII-PAR日变化回归截距随PARmax和VPD的升高而降低,随SWC升高而增加(图4)。由此可知,PAR、VPD和SWC是影响沙柳PSII 光合生理状态的主要因素[39-41],在高辐射、高VPD和干旱等不利环境条件下,沙柳通过由质子动势推动的类囊体酸化过程的诱导等一系列热耗散调节机制耗散过剩的光能[18,34,39],保护自身PSII 反应中心免受损伤。然而随着时间和耗散能量不断增加,一些产物如活性氧的积累破坏了光保护蛋白[27,30],造成PSII 反应中心受损和失活。水作为光合作用的底物和场所,在干旱条件下沙柳对水分的获取远远达不到需求,水分亏缺抑制了光合水裂解释放电子数量[15,40-41],严重影响了光合磷酸化以及暗反应相关酶的活性[34,41],因而进一步导致PSII 光抑制甚至光损伤现象的加剧。通过季节动态、环境因子的相关性以及线性关系的多方面讨论,可见沙柳ФPSII-PAR日变化回归截距也能够很好的代替Fv/Fm,ФPSII-PAR日变化回归斜率和截距可作为反映PSII光合生理状态的适合指标。
4 结论
荒漠灌木沙柳PSII 在日尺度和季节尺度上均受环境波动影响,但基本能通过自身调节恢复到相对适宜状态(Fv/Fm约0.8,ФPSII-PAR日变化回归截距约0.7)。在日尺度上,沙柳ФPSII随PAR的升高而降低,过剩光能以热的形式耗散。在季节尺度上,受物候期影响,沙柳PSII 光合生理状态在其展叶期和叶片枯萎期均处于相对较低生理水平(Fv/Fm<0.75,回归截距<0.65)。高辐射、高VPD和干旱是限制沙柳PSII 光合生理状态的主要环境胁迫。ФPSII-PAR日变化回归斜率和截距是反映PSII 光合生理状态的适合指标,回归截距能够很好的代替Fv/Fm,在一定程度上解决了野外实地长期监测过程中测定Fv/Fm的困难,为探究干旱半干旱区荒漠植物对环境变化响应提供了重要手段。
