急性低氧胁迫和复氧对鲫鱼氧化应激的影响
2020-09-23黄进强权金强吴深基王晓谰潘玉财
张 倩,黄进强,权金强,吴深基,王晓谰,潘玉财
( 甘肃农业大学 动物科学技术学院,甘肃 兰州 730070 )
影响鱼类生长的因素大致分为两大类,一类是鱼类本身的内源因子,另一类是鱼类生存的外部环境因子。内源因子与鱼类自身基因的调控、亲本的遗传特性等相关,以此决定鱼类的生长发育;外源因子(包括温度、溶解氧、光照等其他环境因子)通过刺激机体代谢水平,直接或间接影响鱼类的生长发育,其中溶解氧是鱼类生长和活动的主要限制因子[1-2]。目前低氧胁迫已经成为鱼类生理学的研究热点之一,研究表明,适宜的溶解氧水平可促进鱼类的孵化成活、生长发育及摄食等,而在过饱和或低溶解氧水体中的鱼类,很可能产生强烈的应激反应而影响其正常生活[3]。特别是低氧环境下,易使鱼类正常的呼吸和生理代谢发生紊乱,导致鱼类摄食量下降、生长缓慢,甚至影响鱼类的行为、形态学特征及生存策略[4-6]。研究发现,在低氧环境中西伯利亚鲟(Acipenserbaerii)血清中丙二醛、总蛋白含量均呈先升后降的趋势;河川沙塘鳢(Odontobutispotamophila)心脏、肝脏、鳃中过氧化氢酶活力显著上升[7-8];Lushchak等[9]发现,金鱼(Carassiusauratus)在经过8 h低氧处理后肝脏中谷胱甘肽过氧化物酶活性显著增加79%,复氧14 h后,大脑中谷胱甘肽过氧化物酶活性仍然较高;葛氏鲈塘鳢(Perccottusglenii)在低氧处理2、6 h和10 h下肝脏的超氧化物歧化酶活性显著上调[10]。
血清生理生化指标是反映动物环境应激时体内物质代谢和组织器官机能状态变化的一个重要特征,研究发现,溶解氧水平明显影响养殖鱼类的血液指标,而鳃作为鱼类的呼吸器官,最先感知溶解氧的变化,为机体正常代谢供给氧气发挥着重要作用[11-12]。研究表明,低氧条件下,鱼体肌肉组织中氧化应激现象明显[10,13]。因此,通过研究低氧条件下鱼类血清、鳃和肌肉组织生理水平的变化及其应激效应,对水产养殖具有重大指导意义,亦可为探讨鱼类适应低氧环境的策略提供参考资料。
鲫鱼(Carassiusauratus),属鲤形目、鲤科、鲫属,由于其营养价值高、肉质较嫩、味道鲜美,且生长速度快、适应性强、食性杂等众多优势,在我国淡水养殖业中占据重要地位[14-16]。鲫鱼生活于中下层水域,相对其他常见淡水养殖鱼类对低氧有极强的耐受性,是一个研究低氧胁迫的良好模型[11,17]。然而,目前关于低氧胁迫及复氧对鲫鱼氧化应激影响的研究未见报道。笔者通过对养殖水环境中溶解氧水平的调控,分析在急性低氧和常氧恢复后的不同时间点鲫鱼血清、肌肉及鳃组织中抗氧化酶的变化,探究急性低氧胁迫对鲫鱼氧化应激的影响,以期为鲫鱼的健康养殖提供科学的理论依据。
1 材料与方法
1.1 试验动物
试验用鲫鱼购自兰州市水产品交易市场,将试验鱼运输至实验室循环流水室内养殖槽中暂养14 d,暂养期间连续充气保持溶解氧>6.7 mg/L,温度(25±1) ℃,试验用水均为曝气后的自来水,日换水量约为暂养水体积的1/3。
1.2 试验设计
暂养结束后,挑选40尾规格相近[体质量(250±10) g,体长(24±0.5) cm]、体质健壮、体表无伤的鲫鱼用于试验。通过迅速往养殖槽中注入氮气来降低溶解氧水平,使溶解氧在30 min内降至(1.0±0.1) mg/L,达到所需含量后开始计时,利用溶氧仪来实时监测溶解氧水平的变化,试验过程中通过调节注入空气和氮气的量来维持所需要的溶解氧水平。试验分为低氧胁迫(0、3、6、12、24 h)和常氧恢复(3、6 h)两个阶段,常氧恢复试验在低氧胁迫24 h取样结束后立即进行,恢复期间溶解氧水平为(6.7±0.1) mg/L。
1.3 样品采集与处理
降低溶解氧水平前随机取5尾鲫鱼作为对照组,在低氧胁迫(3、6、12、24 h)和常氧恢复(3、6 h)的各时间点进行尾静脉采血约2 mL,每个时间点取5尾作为生物学重复,血液置于冰箱4 ℃静置2 h后,3500 r/min离心10 min,取上层血清于-80 ℃保存用于测定血清生化指标。采血结束后剖取鲫鱼肌肉和鳃组织(采样时间点及样品数量同血清),置于液氮中速冻后保存于冰箱-80 ℃中用于后续组织中生化指标的测定。
1.4 样品测定
测定血清、肌肉和鳃组织中抗氧化指标,主要包括超氧化物歧化酶(A001-1)、过氧化氢酶(A007-1)、谷胱甘肽过氧化物酶(A005)、总抗氧化能力(A015)、丙二醛(A003-1)和总蛋白(A045-3),试剂盒均购自南京建成生物工程有限公司,吸光度利用紫外可见分光光度计(UNICO UV-3802)测定。
1.5 试验数据分析
试验数据用平均值±标准差表示,利用SPSS 19.0统计软件进行单因素方差分析以及Duncan′s多重比较进行数据差异性分析,P<0.05为有显著统计学差异。
2 结果与分析
2.1 急性低氧胁迫和复氧对鲫鱼血清中氧化应激的影响
低氧胁迫后,鲫鱼血清中丙二醛浓度显著升高(P<0.05),低氧胁迫6 h,丙二醛浓度达到最高,较对照组升高了62%,随着胁迫时间的延长,在胁迫12、24 h时,丙二醛浓度显著下降(P<0.05);恢复3 h时,鲫鱼血清中丙二醛浓度较胁迫24 h显著上升23%(P<0.05),恢复6 h,丙二醛浓度开始下降,与对照组相比差异不显著(P>0.05)(图1)。超氧化物歧化酶活力在低氧胁迫初期显著升高24%(P<0.05),随后在胁迫6 h略有下降,胁迫12、24 h时,鲫鱼血清中超氧化物歧化酶活力较对照组显著升高40%、38%(P<0.05);恢复后,鲫鱼血清中超氧化物歧化酶活力较胁迫24 h显著下降12%、27%(P<0.05),恢复6 h时,鲫鱼血清中超氧化物歧化酶活力略高于对照组,差异不显著(P>0.05)(图2)。过氧化氢酶活力总体呈升高趋势,低氧6、12、24 h时,较对照组显著升高了55%、67%、72%(P<0.05);恢复3 h与6 h时,过氧化氢酶活力较低氧24 h时显著下降(P<0.05),与对照组相比,血清中过氧化氢酶活力略升高,差异不显著(P>0.05)(图3)。谷胱甘肽过氧化物酶活力迅速升高,低氧胁迫12 h时酶活力最高,较对照组显著升高42%(P<0.05),随着胁迫时间的延长,血清中谷胱甘肽过氧化物酶活力开始下降,胁迫24 h时酶活力较12 h时显著下降30%(P<0.05);恢复3 h时,鲫鱼血清中谷胱甘肽过氧化物酶活力较胁迫24 h时显著升高21%(P<0.05),随后逐渐恢复到初始水平(图4)。急性低氧胁迫后,鲫鱼血清中总抗氧化能力活力显著上升,胁迫24 h时酶活力达到最高,低氧3、6、12、24 h时较对照组分别显著升高16%、47%、41%和52%(P<0.05);恢复溶氧3、6 h时,血清中酶活力较胁迫24 h时显著下降24%、39%(P<0.05),恢复6 h酶活力略高于对照组,差异不显著(P>0.05)(图5)。
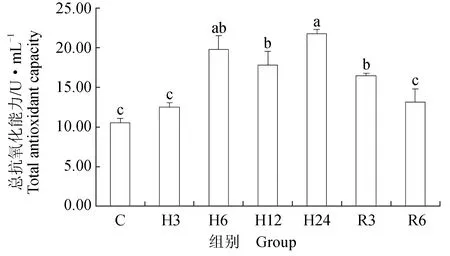
图5 低氧胁迫及复氧对鲫鱼血清中总抗氧化能力的影响
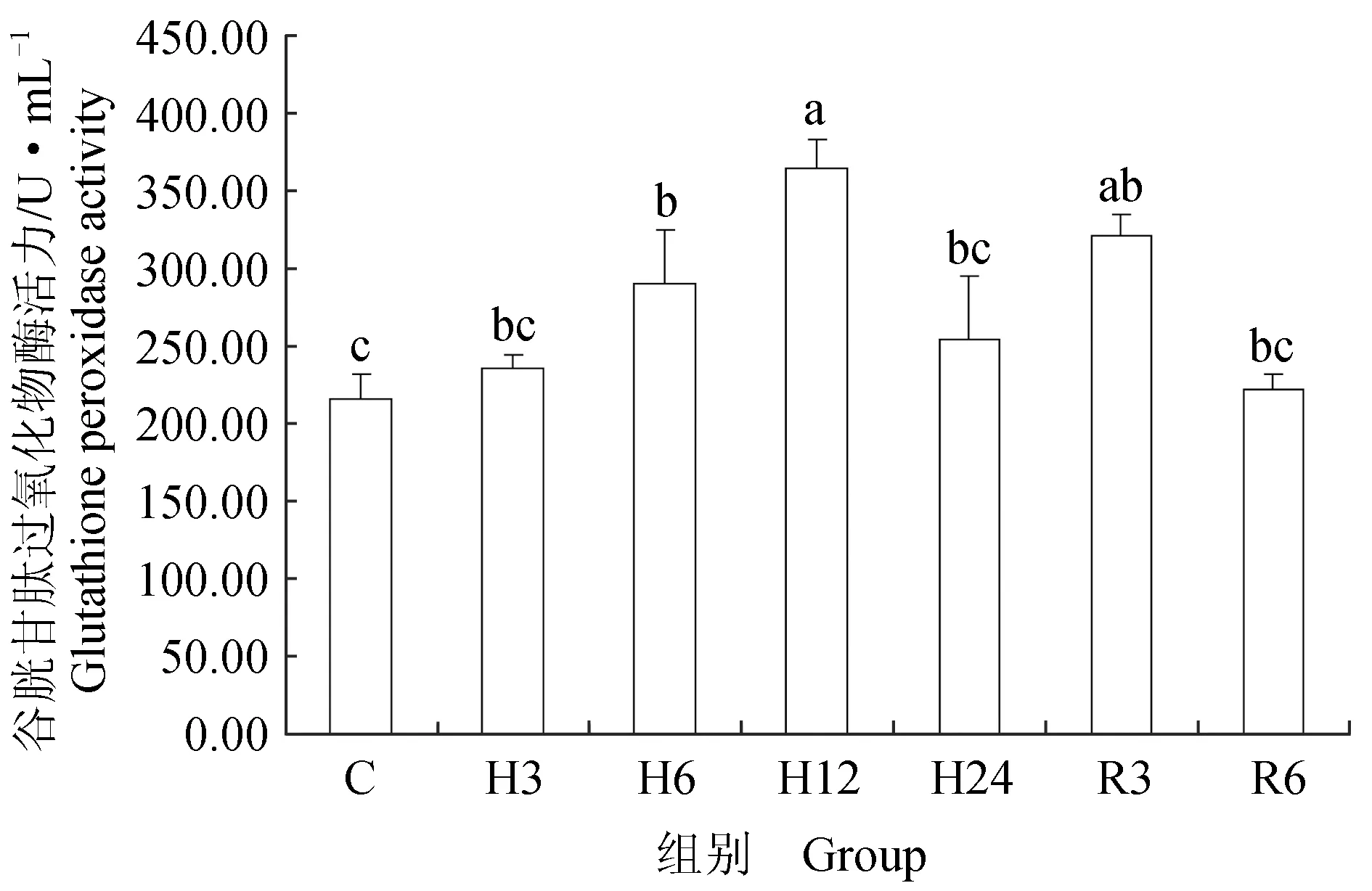
图4 低氧胁迫及复氧对鲫鱼血清中谷胱甘肽过氧化物酶活力的影响
2.2 急性低氧胁迫和复氧对鲫鱼肌肉和鳃组织中氧化应激的影响
肌肉组织中丙二醛含量在急性低氧胁迫3、6、12 h后显著升高(P<0.05),在24 h后又恢复到初始水平;恢复3 h后,丙二醛含量又显著高于对照组(P<0.05),恢复6 h后,显著下降,但仍高于对照组(P<0.05)(图6)。低氧胁迫3、6、12、24 h与复氧3h后超氧化物歧化酶活力显著升高(P<0.05),恢复6 h后与对照组差异不显著(P>0.05)(图7)。过氧化氢酶与谷胱甘肽过氧化物酶活力在低氧胁迫后显著上升(P<0.05)(图8~9),酶活力在低氧6 h时,达到最高(P<0.05),低氧12 h时,酶活力略有下降;恢复6 h后过氧化氢酶活力显著高于对照组(P<0.05),谷胱甘肽过氧化物酶活力恢复到初始值。总抗氧化能力在低氧胁迫3 h后持续升高,保持在较高水平,低氧24 h与恢复3 h时显著升高(P<0.05),24 h时酶活力达到峰值,恢复6 h后,与对照组差异不显著(图10)。总蛋白质量浓度在低氧胁迫后缓慢上升,胁迫12 h时质量浓度显著升高(P<0.05),24 h时质量浓度略有下降;恢复3 h时,质量浓度显著下降,恢复6 h后,恢复到初始值(图11)。
急性低氧胁迫后,鲫鱼鳃组织中丙二醛含量在胁迫3、12、24 h,恢复3 h时显著升高(P<0.05),在低氧24 h时含量最高,恢复6 h后,略高于对照组,与对照组差异不显著(P>0.05)(图6)。低氧6 h时,超氧化物歧化酶活力较对照组显著升高41%(P<0.05),达到峰值,其余时间点超氧化物歧化酶活力较初始值略有变化,差异不显著(P>0.05)(图7)。过氧化氢酶活力在胁迫6、24 h时显著增长48%、40%(P<0.05),其他时间点与对照组差异不显著(P>0.05),恢复6 h后,酶活力略高于初始值,差异不显著(P>0.05)(图8)。低氧胁迫后谷胱甘肽过氧化物酶活力显著升高,6 h时活力最高,比对照组显著增长了52%(P<0.05),恢复6 h后与对照组差异不显著(P>0.05)(图9)。总抗氧化能力在胁迫3、24 h与恢复3 h时酶活力显著升高,胁迫3 h时达到最大,相比对照组显著升高48%(P<0.05),其余时间点与对照组相比差异不显著(P>0.05)(图10)。总蛋白质量浓度在胁迫3 h时显著升高19%(P<0.05),其余各时间点质量浓度高于对照组,但差异不显著(P>0.05)(图11)。
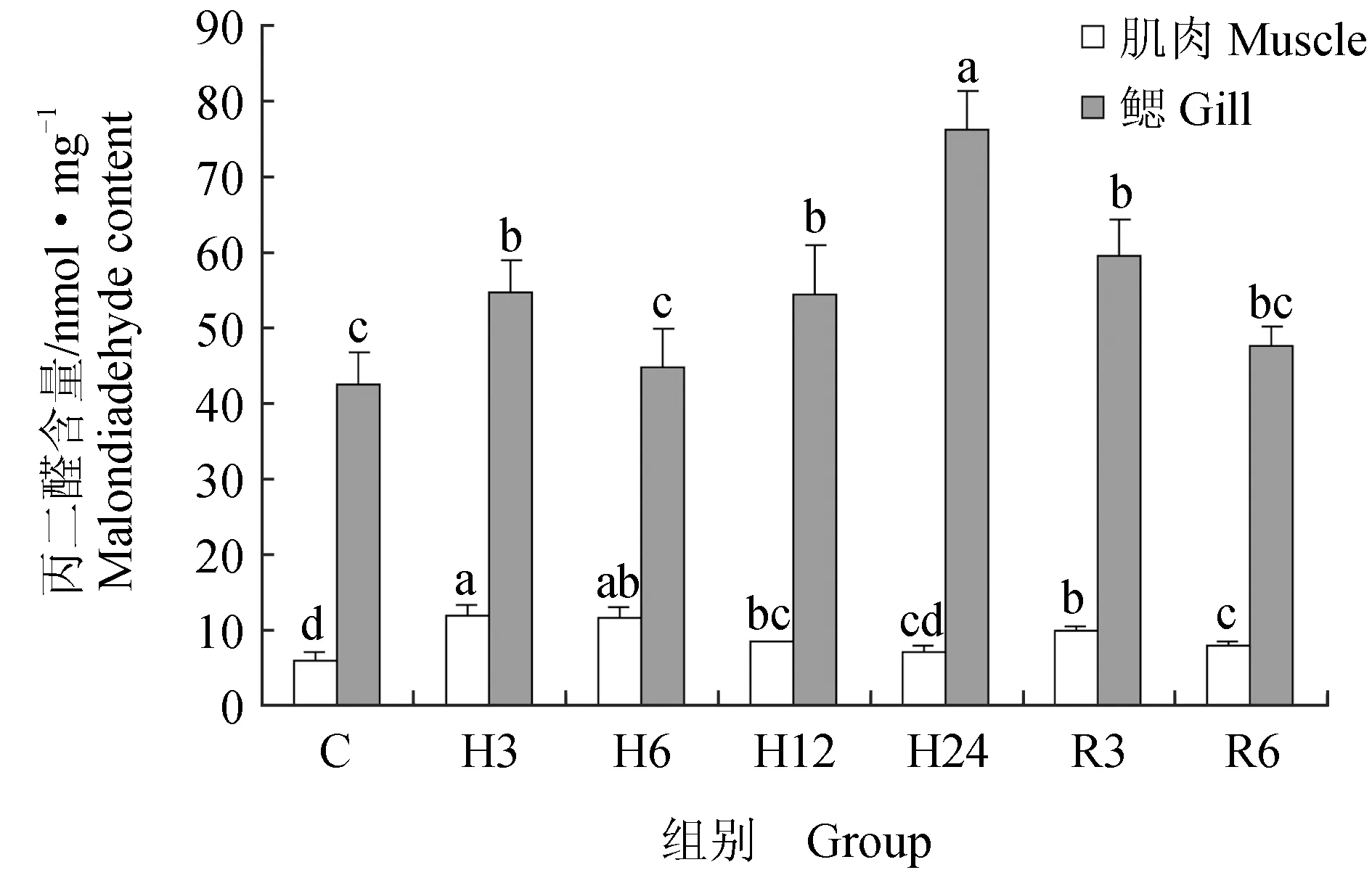
图6 低氧胁迫及复氧对鲫鱼肌肉和鳃组织中丙二醛含量的影响
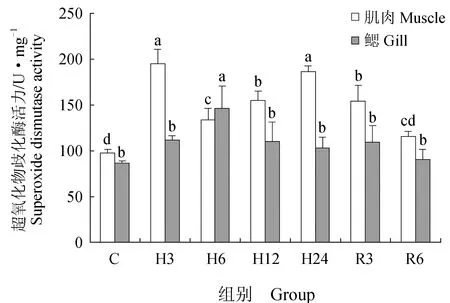
图7 低氧胁迫及复氧对鲫鱼肌肉和鳃组织中超氧化物歧化酶活力的影响
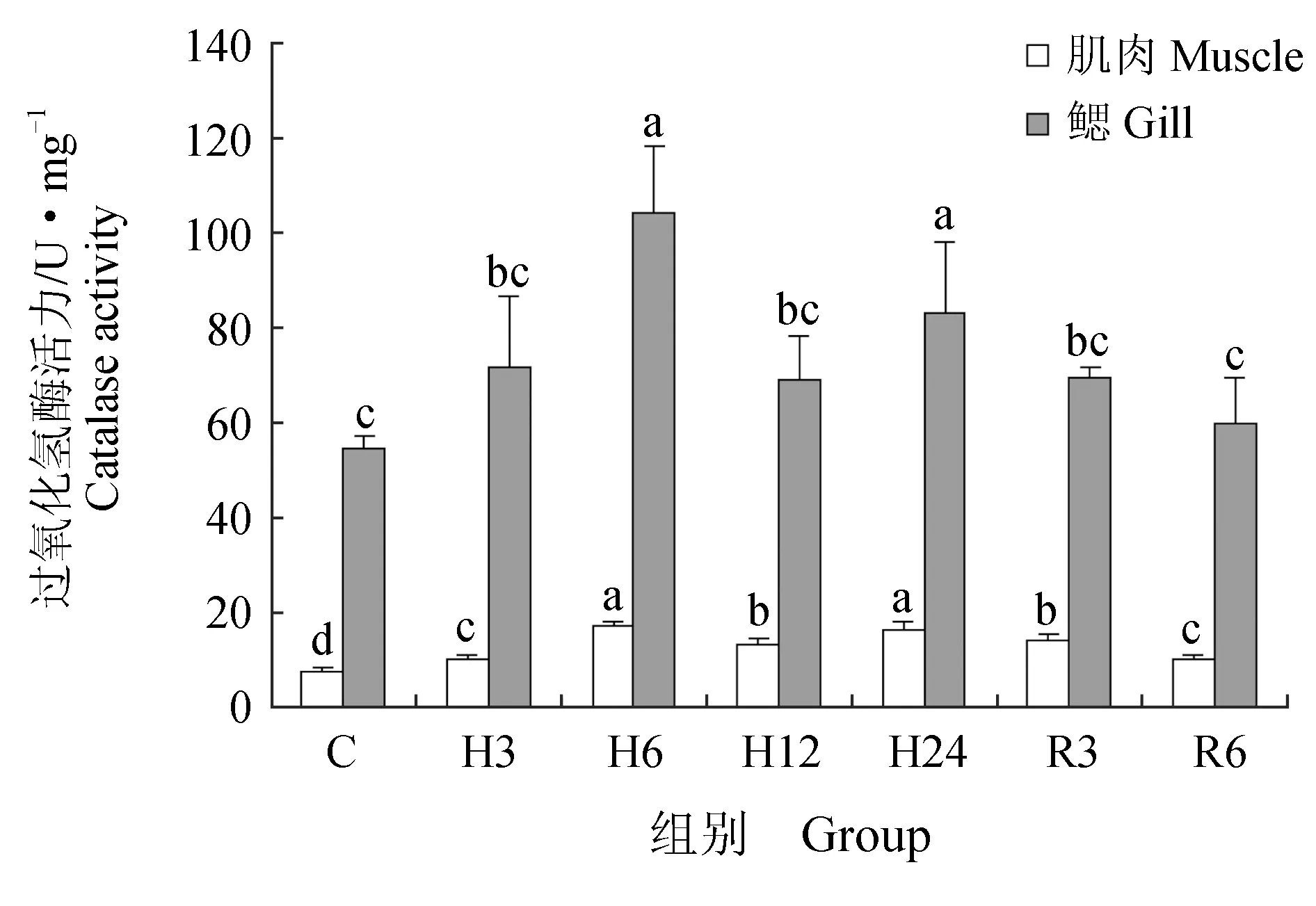
图8 低氧胁迫及复氧对鲫鱼肌肉和鳃组织中过氧化氢酶活力的影响
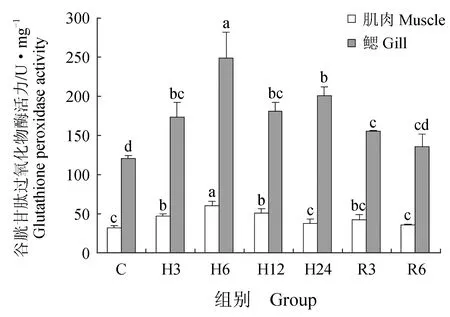
图9 低氧胁迫及复氧对鲫鱼肌肉和鳃组织中谷胱甘肽过氧化物酶活力的影响
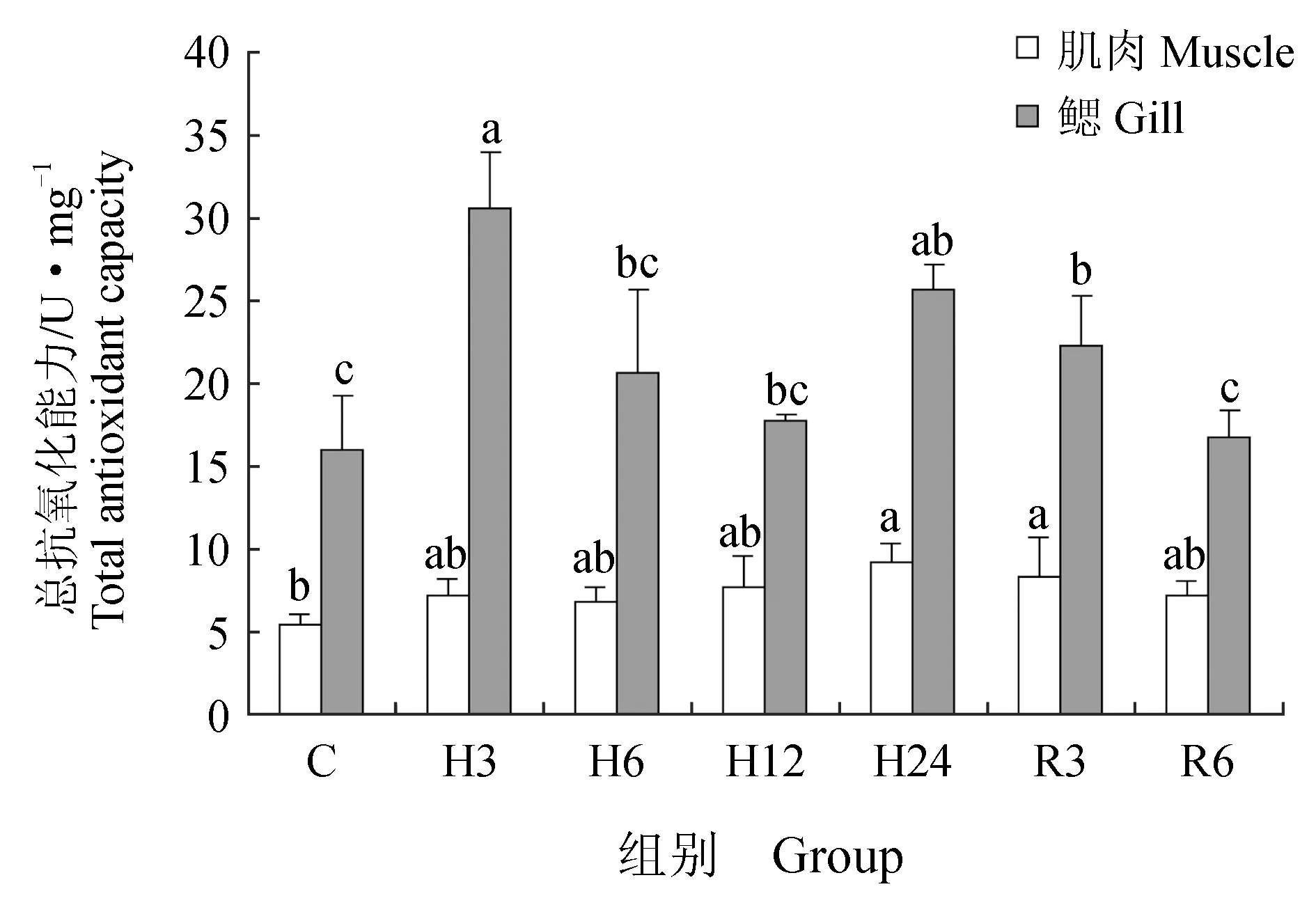
图10 低氧胁迫及复氧对鲫鱼肌肉和鳃组织中总抗氧化能力的影响
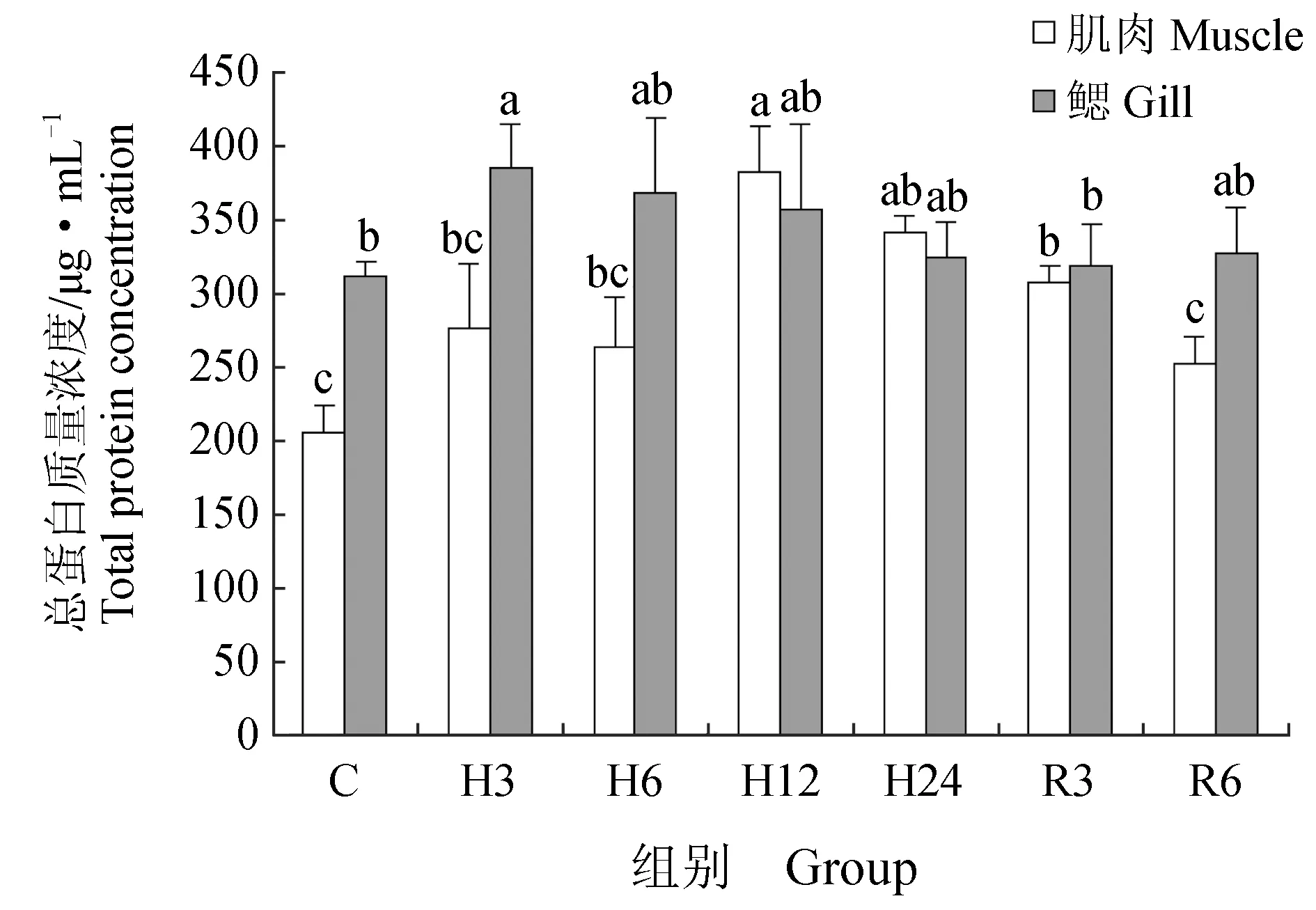
图11 低氧胁迫及复氧对鲫鱼肌肉和鳃组织中总蛋白质量浓度的影响
3 讨 论
鱼类在新陈代谢的过程中,细胞内氧化与抗氧化之间一直处于动态平衡[18],当细胞内产生大量的活性氧自由基超过机体的承载范围时,细胞内过多的自由基会攻击各种生物膜中所含的不饱和脂肪酸,发生连锁的脂质过氧化反应,并因此产生丙二醛等脂质过氧化产物,对机体造成损伤[19-22]。为避免这种情况,机体在长期的进化过程中,形成了一套完整的保护体系,即抗氧化防御系统来清除体内多余的氧自由基,保护细胞免受氧化损伤[23-24]。超氧化物歧化酶、过氧化氢酶和谷胱甘肽过氧化物酶等作为抗氧化防御系统中的重要组成部分,在机体的氧化与抗氧化平衡中起着重要作用,对机体细胞损伤后的氧化过程具有很强的防御功能[21]。研究认为,超氧化物歧化酶在清除体内活性氧自由基的过程中最早发挥作用,它促使超氧阴离子(O2-·)歧化为H2O2和H2O,随后过氧化氢酶和谷胱甘肽过氧化物酶再将H2O2催化为H2O和O2,因此超氧化物歧化酶、谷胱甘肽过氧化物酶和过氧化氢酶等具有清除体内多余的活性氧自由基和保护机体免受氧化损伤的作用[21]。
3.1 急性低氧胁迫和复氧对鲫鱼血清中氧化应激的影响
丙二醛的含量可反映机体内脂质过氧化的程度,从而间接反映出细胞损伤的程度[25]。本试验中,随着溶解氧水平的下降,鲫鱼血清、肌肉和鳃组织中丙二醛含量初期快速升高,这表明低氧迫使鲫鱼处于应激状态,由于机体内产生大量的活性氧自由基,使得机体内脂质过氧化程度升高,产生大量的脂质过氧化产物,随着胁迫时间的延长,丙二醛含量显著下降,机体通过调节抗氧化酶的活性来抵御缺氧造成的氧化压力,保护机体免受氧化损伤;恢复溶氧后丙二醛含量再次升高,恢复后期丙二醛含量逐渐下降至初始水平,说明此时恢复溶氧对鲫鱼而言也是一种应激[26],机体会再次激活体内抗氧化防御系统。这与Filho等[3,13]限制溶解氧供应对鱼类影响的研究结果一致。
张国松[27]在瓦氏黄颡鱼(Pelteobagrusvachelli)应对低氧胁迫的耐受机制的研究中发现,低氧胁迫促使瓦氏黄颡鱼大脑中超氧化物歧化酶活力持续上调,常氧恢复后超氧化物歧化酶活力与对照组相近,本试验结果与之相似。本试验中,急性低氧胁迫后,鲫鱼血清中超氧化物歧化酶活力在短期内迅速升高,表明当机体受到低氧刺激时,超氧化物歧化酶首先发挥作用,促使超氧阴离子(O2-·)歧化为H2O2和H2O,随着胁迫时间的延长,超氧化物歧化酶活力略有下降,随后显著升高,这是由于低氧胁迫下鲫鱼通过无氧呼吸途径[28],产生大量活性氧自由基,使得抗氧化酶活力下降,但随着时间的延长,鲫鱼机体内出现了代偿应激导致超氧化物歧化酶活力迅速上升,从而抵御氧自由基对机体的损伤。常氧恢复后期酶活力略高于对照组,推测此时常氧恢复促使机体仍处于应激状态,超氧化物歧化酶持续为清除体内活性氧自由基发挥作用。
本试验中过氧化氢酶和谷胱甘肽过氧化物酶活力在急性低氧胁迫后显著升高,常氧恢复后与对照组相比差异不显著,表明了机体内抗氧化酶类的协同性。杨凯[29]的研究结果表明,低氧胁迫后黄颡鱼(P.fulvidraco)血清中过氧化氢酶活力显著增大,但随着时间延长活性逐渐下降,这与本试验结果类似,说明血清中过氧化氢酶和谷胱甘肽过氧化物酶在鲫鱼面临氧化应激时出现协同调节作用,避免机体受到氧化损伤的危害。陈世喜等[30]研究发现,急性低氧胁迫下,卵形鲳鲹(Trachinotusovatus)幼鱼肝脏中过氧化氢酶显著升高后维持在一个较高水平,而谷胱甘肽过氧化物酶活力在持续上升后逐渐下降,慢性低氧胁迫后谷胱甘肽过氧化物酶活力显著升高,与本试验结果类似。
李根瑞[31]研究发现,仿刺参(Apostichopusjaponicus)在低氧环境中肌肉、呼吸树和消化道中总抗氧化能力显著升高,复氧初期总抗氧化能力仍显著高于对照组,随后慢慢恢复到初始水平。管越强等[32]在研究低氧胁迫对日本沼虾(Macrobrachiumnipponense)的影响时发现相似结果,低氧胁迫后日本沼虾肝胰腺和肌肉中总抗氧化能力有显著影响。本试验中,鲫鱼体内总抗氧化能力在低氧胁迫后迅速显著上升,表明鲫鱼能快速对低氧胁迫产生响应,有效清除体内氧自由基,常氧恢复后总抗氧化能力较低氧胁迫后期显著下降,恢复后期总抗氧化能力略高于对照组,推测是常氧恢复后机体在短期内对总抗氧化能力的代谢不完全造成的。
3.2 急性低氧胁迫和复氧对鲫鱼肌肉和鳃组织中氧化应激的影响
机体处于应激状态时,当机体内大量产生的活性氧因不能及时清除而累积在体内时,就会导致生物膜上的脂质物质遭受过氧化,生成丙二醛等有害物质,造成机体多种酶失活,导致细胞发生变性甚至死亡,使机体的免疫能力下降[33-34]。本试验中,急性低氧胁迫3 h后,鲫鱼肌肉和鳃组织中丙二醛含量显著升高,肌肉组织中丙二醛含量达到峰值,鳃组织中丙二醛含量在胁迫24 h达到峰值。恢复溶氧后,丙二醛含量在组织中先升后降,与对照组差异不显著。表明机体在低氧刺激后,由于体内大量产生活性氧,导致机体脂质过氧化程度升高,产生大量丙二醛,丙二醛含量在体内的变化在一定程度上反映了机体保护自身的反应机制。常氧恢复后组织中丙二醛含量在短暂升高后恢复至初始值,说明此时复氧对鲫鱼造成了二次应激,体内抗氧化系统再次调动保护机体免受损伤。郭志雄等[35]在急性低氧胁迫对军曹鱼肝脏氧化应激的影响中发现同样结果,胁迫后军曹鱼体内丙二醛含量显著上升。
当低氧胁迫导致体内活性氧自由基增多时,超氧化物歧化酶活力也会随之发生变化。马粒雅等[36]在溶解氧变化对中华乌塘鳢(Bostrychussinensis)酶活的影响中发现,随着溶解氧含量的变化,中华乌塘鳢肝脏和肠内超氧化物酶活力在溶解氧水平下降初期变化不大,在溶解氧水平降至1.04 mg/L时,酶活力显著上升,达到峰值后开始逐渐下降,随着溶解氧水平降到0 mg/L,酶活力又开始增大。本试验中,急性低氧胁迫后,鲫鱼肌肉和鳃组织中超氧化物歧化酶出现了波动性的变化,表明超氧化物歧化酶能通过酶活性的变化来调节体内自由基的平衡,维持机体内环境的稳定。
过氧化氢酶和谷胱甘肽过氧化物酶在机体抗氧化系统中的主要作用是清除H2O2,起到保护细胞膜结构和功能完整性的作用[29]。杨明等[37]在研究日本沼虾在低氧胁迫和复氧下体内抗氧化酶活力的变化中发现,低氧胁迫后日本沼虾肌肉、鳃组织中过氧化氢酶、谷胱甘肽过氧化物酶活力在胁迫12 h时酶活力显著高于对照组,复氧后酶活力出现波动性变化。本试验中,鲫鱼在急性低氧胁迫后,肌肉组织及鳃组织中过氧化氢酶及谷胱甘肽过氧化物酶活力显著升高,在胁迫6 h时达到峰值,胁迫12 h二者酶活力显著下降,这表明机体内过氧化氢酶和谷胱甘肽过氧化物酶具有一定的协同作用,二者共同为清除体内脂质过氧化产物发生作用,而在12 h酶活力显著下降,其原因可能是超氧化物歧化酶通过歧化反应产生过量的H2O2,超过了机体的承受范围,导致酶活力受到抑制。复氧后,机体内抗氧化防御系统清除了过量的有害物质,酶活力逐渐恢复至初始值。
总抗氧化能力是用于衡量机体抗氧化系统功能状况的综合性指标,代表和反映了机体抗氧化防御系统对外来刺激的代偿能力以及机体自由基代谢的状态[19]。本试验中,鲫鱼肌肉组织中总抗氧化能力在胁迫后持续升高,胁迫24 h时最大,鳃组织中总抗氧化能力在胁迫3 h达到峰值后开始下降,胁迫24 h显著升高,表明机体在应激状态下,体内抗氧化防御系统能在应对不良刺激时做出响应,保护机体。复氧后肌肉及鳃组织中总抗氧化能力开始下降,恢复至初始值。这与李根瑞[31]研究结果相似。
总蛋白具有维持血液正常胶体渗透压和pH恒定的作用,并与脂肪酸、固醇等物质运输及免疫功能相关[33]。本试验中,急性低氧胁迫下鲫鱼组织中总蛋白含量随胁迫时间的延长而逐渐升高,常氧恢复后总蛋白含量与对照组差异不显著。这同陈静怡等[7]研究发现20、28 ℃时低氧胁迫导致西伯利亚鲟血清中总蛋白含量随胁迫时间的延长而升高,在达到峰值后逐渐降低,最终与对照组差异不显著的结果相似。
4 结 论
急性低氧胁迫会使鱼类产生氧化应激反应,而应激后的快速恢复溶氧,也会使鱼类产生应激,调动体内的抗氧化防御系统。低氧胁迫下,鲫鱼血清、肌肉和鳃组织中抗氧化酶活力升高,共同清除体内因缺氧产生的大量活性氧自由基,常氧恢复后,机体又会产生大量活性氧自由基,同时再次调动抗氧化系统开启防御机制,抵御氧化应激对机体造成的损伤,这是机体为了适应低氧胁迫和胁迫后复氧所采取的抗氧化策略。本试验结果为鱼类低氧胁迫生理学研究提供了基础资料,也为鲫鱼养殖业的健康发展提供理论依据。
