Pathological significance of tRNA-derived small RNAs in neurological disorders
2020-09-18ChuanQinPeiPeiXuXinZhangChaoZhangChangBinLiuDeGangYangFengGaoMingLiangYangLiangJieDuJianJunLi
Chuan Qin , Pei-Pei Xu , Xin Zhang , Chao Zhang , Chang-Bin Liu , De-Gang Yang , Feng Gao ,Ming-Liang Yang , Liang-Jie Du , Jian-Jun Li ,
1 School of Rehabilitation Medicine, Capital Medical University, Beijing, China 2 China Rehabilitation Science Institute, Beijing, China 3 Center of Neural Injury and Repair, Beijing Institute for Brain Disorders, Beijing, China 4 Department of Spinal and Neural Functional Reconstruction, China Rehabilitation Research Center, Beijing, China 5 Beijing Key Laboratory of Neural Injury and Rehabilitation, Beijing, China
Abstract
Key Words: epigenetics; molecular biology; neurological disorders; review; sequencing; stress; tRNA; tRNA-derived fragments; tRNA-derived small RNAs; tRNA-derived stress-induced RNA
Introduction
Non-coding RNAs (ncRNAs) are RNAs that are not translated into proteins (Xu et al., 2019). The most common type of small ncRNAs, transfer RNAs (tRNAs), are universal nucleic acid entities; they are the second most abundant type of RNAs in cells, accounting for at least 4% of all cellular RNA species (Green et al., 2010; Kirchner and Ignatova, 2015).Traditionally, tRNAs translate nucleic acid code into amino acid sequences by decoding messenger RNA during protein translation (Li, 2007). Apart from this canonical function,tRNAs are also involved in other pathophysiological processes, such as the regulation of apoptosis (Mei et al., 2010), cell wall biosynthesis (Dare and Ibba, 2012), and other biological functions (Raina and Ibba, 2014).
In terms of tRNA biogenesis, the formation of mature tRNA starts with the transcription of its precursor tRNA(pre-tRNA), which requires RNA polymerase III. After this, the 5′ and 3′ U trailer sequences are cleaved by the endonucleases RNase P and RNase Z (Frank and Pace, 1998;Ceballos and Vioque, 2007). After the removal of intronic sequences by the tRNA-splicing endonuclease, tRNA nucleotidyltransferase performs 3′ CCA tailing, and other post-transcriptional modifications are completed (Keam and Hutvagner, 2015; Fröhlich et al., 2018). Mature tRNAs have a clover-leaf secondary structure and are typically 73-90 nucleotides (nts) in length, and include a T-loop (also known as a TC loop), a D-loop, a variable loop, an acceptor stem,and an anticodon loop. The tertiary structure of tRNA forms an L-shape, which is maintained by hydrogen bonds (Kirchner and Ignatova, 2015).
Numerous studies have demonstrated that tRNAs can be cleaved into a heterogeneous population of small ncRNAs with lengths of 18-40 nts, known as tRNA-derived small RNAs (tsRNAs) (Phizicky and Hopper, 2010; Kumar et al., 2014; Zhou et al., 2017). These RNA species were first considered to be the byproducts of random tRNA cleavage.However, mounting evidence has demonstrated their critical functions as regulatory factors in the pathophysiological processes of various diseases, with roles including tumor proliferation in ovarian cancer cells (Zhou et al., 2017), genome stability maintenance (Sharma et al., 2016), potential biomarkers in prostate cancer (Olvedy et al., 2016), ribosome biogenesis (Kim et al., 2017), breast cancer (Zhou et al., 2019), the stress response (Emara et al., 2010), maternal overnutrition (Sarker et al., 2019) and cell cycle regulation in non-small-cell lung cancer (Shao et al., 2017).
Although some tsRNA functions have been revealed in various physiological and pathological conditions, the underlying mechanisms by which tsRNAs affect specific cellular processes are largely unknown. The present study summarizes the current knowledge around tRNAs and discusses tRNA biogenesis, biological functions, and potential mechanisms,focusing on their associations with neurological diseases.
To electronically search the Medline database for literature describing tsRNA from 1946 to 2018, the following search criteria was used: tsRNA and tRNA-derived fragments(tRFs) and tRNA-derived stress-induced RNA (tiRNA). In this study we further screened the results by headings and abstracts and included all articles. We focused on articles relating to neurological diseases.
Search Strategy
The articles used in this review were retrieved by replicating the search terms. An electronic search of the Medline database for literature describing tsRNA from 1970 to 2019 was performed using the following conditions: tRNA-derived small RNAs OR tRNA-derived fragments OR tRNA-derived stress-induced RNA. The results were further screened by title and abstract.
In addition, we also completed an electronic search of the Medline database for methods of inducing human trials. This search included publications prior to March 2019, and used the following search criteria: tRNA-derived small RNAs OR tRNA-derived fragments OR tRNA-derived stress-induced RNA. Subsequent searches were completed that were specifically relevant to each method discussed with the following term: neurological disorders.
Discovery, Classification, and Biogenesis of rRNA-Derived Small RNA
Discovery
In 1977, Borek et al. first determined that tsRNAs were able to “turnover” during the degradation of tRNAs in tumor tissues. Two years later, Speer et al. (1979) suggested that one or more of these tRNA breakdown products might act as a potential clinical cancer biomarker (Ahmed et al., 2017).With the rapid development of high-throughput sequencing technology, a variety of tsRNAs have subsequently been found in viruses (Yeung et al., 2009; Reese et al., 2010), bacteria (Kumar et al., 2014), archaea (Heyer et al., 2014), plants(Thompson et al., 2008; Chen et al., 2011), protozoa (Couvillion et al., 2010), mice (Li et al., 2012), chickens (Liao et al., 2010), and humans (Lee et al., 2009; Kumar et al., 2015).Therefore, tsRNAs are found throughout the evolutionary tree. Additionally, biological links have been identified between tsRNAs and numerous organisms.
Classification and biogenesis
Genes for tRNA are transcribed into pre-tRNAs by RNA polymerase III, and additional bases are included at their 5′ and 3′ ends. These additional bases form the leader and trailer sequences, respectively (Phizicky and Hopper, 2010).After this, RNase P and RNase Z-like enzymes remove the leader and trailer sequences of pre-tRNA, which has a length of 50 nts. Next, a CCA terminal nucleotide is added to the 3′ end of the trailer-free tRNA by the enzyme tRNA nucleotidyltransferase, for tRNA maturation (Maraia and Lamichhane, 2011). Finally, mature tRNAs are transported to the cytoplasm by exportin-T, which requires the mature 5′ and 3′ends of the tRNA as well as an added, non-templated single CCA sequence (Xiong and Steitz, 2006). Several enzymes are critical for the generation of tsRNAs in vertebrates, namely RNase Z, Dicer, and angiogenin (Cole et al., 2009; Fu et al.,2009; Haussecker et al., 2010). Generally, tsRNAs are divided into two main types, tRFs and tiRNAs, based on their length and cleavage sites. There is currently no consensus regarding consistent nomenclature for tsRNAs. As a result, the present review uses a comprehensive description based on several new publications (Keam and Hutvagner, 2015; Kumar et al., 2016; Kim et al., 2017; Soares and Santos, 2017; Li et al.,2018b; Park and Kim, 2018; Shen et al., 2018).
tRNA-derived fragments
tRFs are a type of small RNA with a length of 14-30 nts that can be produced from precursor and mature tRNA transcripts (Keam et al., 2014). Notably, they have a similar size to miRNA, with a 5′ phosphate and a 3′ hydroxyl group(Couvillion et al., 2010). To date, various tRFs have been identified following the development of high-throughput sequencing. Based on their canonicity, tRFs are broadly divided into two groups: typical and atypical tRFs.
There are three major subclasses of tRFs based on their different mapped positions: tRF-5, tRF-3, and tRF-1 (Figure 1). tRF-5 are 14-30 nts in length and are generated by cleavage in the D-loop or stem region of tRNA by Dicer, which is usually aimed at the D-loop and anticodon loop of tRNAs(Kumar et al., 2014). The three groups of tRF-5s are classified by their lengths: tRF-5a (14-16 nts), tRF-5b (22-24 nts), and tRF-5c (28-30 nts). The lengths of these tRF-5 series follow a normal distribution. tRF-3s are generated from cleavage sites in the T-loop by Dicer, angiogenin, or other members of the RNase A superfamily. They are further divided into tRF-3a or tRF-3b subgroups, and lengths of 13-22 nts. In contrast to tRF-5 and tRF-3, the abundance of tRF-1 is usually several orders of magnitude lower than the highest abundance fragments (Olvedy et al., 2016). tRF-1 are derived from cleavage of the 3′ ends of pre-tRNAs. tRF-1001, the first type of tRF-1 to be identified, is derived from the 3′ end of a Ser-TGA pre-tRNA transcript that is not retained in the mature tRNA.tRF-1001 has relatively high expression in a variety of cancer cells and is closely related to cell proliferation (Lee et al.,2009).
In addition to typical tRFs, there are other tRNA fragments, known as atypical tRFs, which can be identified from sequencing data despite their low abundance. For instance,novel unclassified tRFs that were first identified in breast cancer cell lines are known as tRF-2. These are derived from tRNAGlu, tRNAAsp, tRNAGly, and tRNATyr; they comprise anticodon stem loop sequences and fail to meet the conditions to be considered tRFs using the general classification scheme (Schaffer et al., 2014; Goodarzi et al., 2015). One publication has stated that tRF-2 is broadly generated under hypoxic stress conditions (Kumar et al., 2016). However,the details of the RNase process for the production of tRF-2 remain unclear (Schorn et al., 2017). In addition, a novel subclass of tRFs that are mostly derived from mature tRNA internal structural domains with variable sizes are classified here as i-tRFs. These are reported to be strongly associated with disease states in their lengths and expression levels(Hanada et al., 2013; Goodarzi et al., 2015). tRFs derived from intron-containing tRNAs are also detected in cleavage and polyadenylation factor I subunit 1 (CLP1) knockout mice (Hanada et al., 2013). CLP1 is required for the proper functioning of the tRNA splicing endonuclease, and its absence is strongly associated with the accumulation of unspliced pre-tRNAs and 5′ leader exon Tyr-tRFs, which accumulate in response to oxidative stress.
tRNA-derived stress-induced RNA
tiRNA, or tRNA-halves, are another type of tsRNAs, and are characterized by a specific cleavage in the anticodon loops of mature tRNAs with a length of 31-40 nts. They are generally generated under stress conditions, including starvation, oxidative damage, heat shock, hypoxia, and radiation (Lee and Collins, 2005; Jöchl et al., 2008; Li et al., 2008; Thompson et al., 2008, 2009; Hsieh et al., 2009). In some species, for example, tRNA can be cleaved under stress conditions into tiRNAs; this process is regulated by angiogenin (Fu et al., 2009;Yamasaki et al., 2009). However, some tiRNAs are reportedly induced under non-stress conditions (Thompson and Parker,2009). Two subclasses of tiRNAs have been identified based on whether they have a 5′ or 3′ sequence at the anticodon cleavage site, and are known as 5′-tiRNAs and 3′-tiRNAs(Li and Hu, 2012; Saikia and Hatzoglou, 2015), as shown in Figure 1. Furthermore, 5′ tiRNAs range from the 5′ end of mature tRNAs to the end of the anticodon loop, while 3′ tiRNAs start from the anti-codon loop and end at the 3′ end of mature tRNAs. In terms of intracellular localization, the majority of tiRNAs are located in the cytoplasm, although some are located in the nucleus and mitochondria (Dhahbi et al.,2013).
A variety of studies have identified a close relationship between tiRNAs and stress conditions (Hartmann et al., 1999;Jöchl et al., 2008; Li et al., 2012). Furthermore, tiRNAs induced by oxidative stress, heat shock, or UV irradiation have been reported to repress translation in a phospho-eIF2α-independent manner (Yamasaki et al., 2009). In mammalian cells, tiRNAs play an important role in the inhibition of protein synthesis and the intrinsic pathway of apoptosis (Saikia and Hatzoglou, 2015). A group of special tiRNAs known as sex-hormone-dependent tRNA-derived RNAs, which are produced with participation by angiogenin, are notably not produced under stress conditions. Instead, they are highly expressed in estrogen-receptor-positive breast cancer and androgen-receptor-positive prostate cancer cell lines, thus indicating their hormone- and receptor-dependent features(Honda et al., 2015).
Recently, in addition to the traditional identification of tiRNAs based on sequences and structure, computational approaches using functional information have provided new identification options (Pagès et al., 2018). For example,researchers have developed SeRPeNT, which uses reproducibility across relevant tools to process small ncRNAs by the means of harnessing the power of read profiles (Pagès et al.,2018).
Key enzymes in tsRNA production
Three key enzymes are involved in the generation and maturation of tRFs and tiRNAs, namely RNase Z or ElaC domain protein 2 (ELAC2), Dicer, and angiogenin (Fu et al., 2009);each has a different role. RNase Z is an endonuclease that is involved in the maturation of the 3′ end of tRNAs (Xie et al.,2011). A recent study showed that RNase Z is required for tRNA processing and the balanced maintenance of tRFs (Siira et al., 2018). Furthermore, it has been reported that RNase Z cleaves tRNAs into tRF-1001, the first tRF of its series to be identified (Lee et al., 2009). Recent studies have indicated that the processing of miRNAs and some tRFs is Dicer-dependent in humans (Maute et al., 2013), mice (Babiarz et al., 2008), and zebrafish (Soares et al., 2015). For example,decreasing Dicer expression can result in downregulation of tRFs derived from tRNAGln, suggesting Dicer involvement in this process (Cole et al., 2009). Similar results were observed in human mature B cells, which experienced Dicer1-dependent biogenesis (Maute et al., 2013). However,another study reported a contradictory result, demonstrating that the production of numerous 5′ and 3′ tRFs with lengths of 22-30 nts was not Dicer-dependent in a range of human cell lines and in experiments in Drosophila or Schizosaccharomyces pombe (Li et al., 2012). Thus, whether Dicer is critical to the biogenesis of tRFs may depend on cell type, tissue,and species. In addition, some studies have reported that angiogenin is strongly associated with the generation of these smaller 5′ and 3′ tRFs and tiRNAs. For example, the generation of 5′ and 3′ tRFs and tiRNAs with lengths of 22-30 nts increased after incubation with angiogenin in HEK293 cell lines (Li et al., 2012). Angiogenin secretion is known to increase under hypoxic conditions in cancer cells and is a critical factor in the stress response (Hartmann et al., 1999).Nevertheless, in vivo studies are still needed to validate the involvement of angiogenin in the process of tsRNAs.tRNA modification systems affect tsRNA biogenesis
tRNAs are critical for every cellular process. Thus, it is very important to ensure that tRNAs have normal structure, function, and stability, and this occurs via various post-transcriptional modifications (Torres et al., 2014). Indeed, most of these modifications are located in the anticodon loop, with methylation of tRNA functioning in interactions with proteins and decoding fidelity (Yacoubi et al., 2012). The generation of tsRNAs has also been reported to be influenced by these tRNA modifications. For instance, tRNA-methyltransferase Trm9 catalyzes 5-methylcarboxylmethyl modification of uridine, resulting in resistance to tRNA cleavage in yeast(Klassen et al., 2008). In another example, Dnmt2-mediated 5-methylcytosine modifications repressed the specific cleavage of the tRNA anticodon loop regulated by angiogenin,which decreased tiRNA levels (Schaefer et al., 2010). In terms of the neurodegenerative process, genetic mutations in various tRNA modification systems are frequently related to stress conditions such as irradiation, toxicity, ischemia, and acute injury, and stress conditions including ischemia and oxidative stress are the most common events following neurodegeneration (Cherbuin et al., 2019). These stress conditions change the tertiary structures of mature tRNAs, causing the production of specific tsRNAs, thus affecting neurological disorders (Mishima et al., 2014). Furthermore, emerging evidence has demonstrated that these modifications play an important role in the pathophysiology of human diseases,such as those of the muscle and nervous system, as reviewed in references. For example, the abundance, modification,and aminoacylation levels of tRNAs lead to mRNA decoding in ways that reflect the cell type and its environment in neurodegeneration. Moreover, tRNAs also interact with various proteins that are not involved in translation, and this could coordinate translation activity in nerve cells (Torres et al.,2014; Pan, 2018).
Biological Functions of tsRNA
Despite an increasing amount of evidence for the production of tsRNAs in various cell lines (Li et al., 2012; Torres et al.,2014; Pan, 2018; Dou et al., 2019), controversy remains over whether tsRNAs might be the products of aberrant degradation by specific endonucleases. However, an increasing number of tRFs and tiRNAs have been reported to act as functional regulatory factors in physiological processes and in cellular metabolisms, including in neurological disorders,indicating their non-random processing (Figure 2).
miRNA-like actions in gene expression regulation
As a type of small non-coding RNA, there is mounting evidence that some tsRNAs regulate mRNA expression levels by directly binding to the target mRNA and acting as miRNAs. For example, in several colorectal cancer cell lines, tRNALeu-derived fragments with lengths of 17 nts were found to support the function of cancer stem-like cells in colorectal cancer progression. They directly targeted JAG2 as miRNAs through 3′ UTR luciferase assays, which is a canonical mechanism (Huang et al., 2017). However, in H1299 cells,which are a model of non-small-cell lung cancer, tRF-Leu-CAG knockdown caused the inhibition of aurora kinase A(AURKA), leading to suppressed cell proliferation and an impeded cell cycle. This suggests uncanonical miRNA- or siRNA-like activity (Shao et al., 2017). Moreover, in terms of neurological disorders, little evidence of miRNA-like function for tsRNA has been found. Thus, our team has recently focused on miRNA-like functions of tsRNA in neurological diseases using sequencing screening and functional verification to select specific tsRNAs and their target genes, such as BDNF. Experimental results were satisfactory and the paper is currently under review. Because tRFs are evolutionarily ancient and are present in various species, it is plausible that some tRFs inhibit the translation of target mRNA and other translational factors through complementary base pairing, including in cancers, neurological diseases, and other disorders. Notably, these tsRNAs are physically related to Argonaute (Ago) proteins (Ago1-4) (Maute et al., 2013),and any association between Ago and tsRNAs has biological significance. Recently, an increasing amount of evidence has supported the function of tsRNAs in Ago complexes.For example, in both embryonic stem cells and embryos, a decrease in the gene expression of tRF-5 derived from tRNA-GlyGCC was related to the Ago1 complex (Sharma et al., 2016). Another study identified pre-tRNA-IleUAU as a miRNA precursor that is transported into the cytoplasm for Dicer processing and Ago loading (Hasler et al., 2016). We know that miRNA preferentially binds to Ago2, which is associated with post-transcriptional RNA silencing (Matsui et al., 2016). In addition, a meta-analysis stated that, in human HEK293 cells, tsRNAs are strongly associated with Ago1,Ago3, and Ago4, which all play a major role in RNA regulation, in contrast to Ago2 (Kumar et al., 2014). Nevertheless,tRF preferences change from Ago1 to Ago2 in young and old flies, respectively (Karaiskos et al., 2015). Thus, to determine the endogenous mRNA targets of tsRNAs, more studies should investigate the relationships between tsRNAs and Ago complexes.
Regulation of protein translation
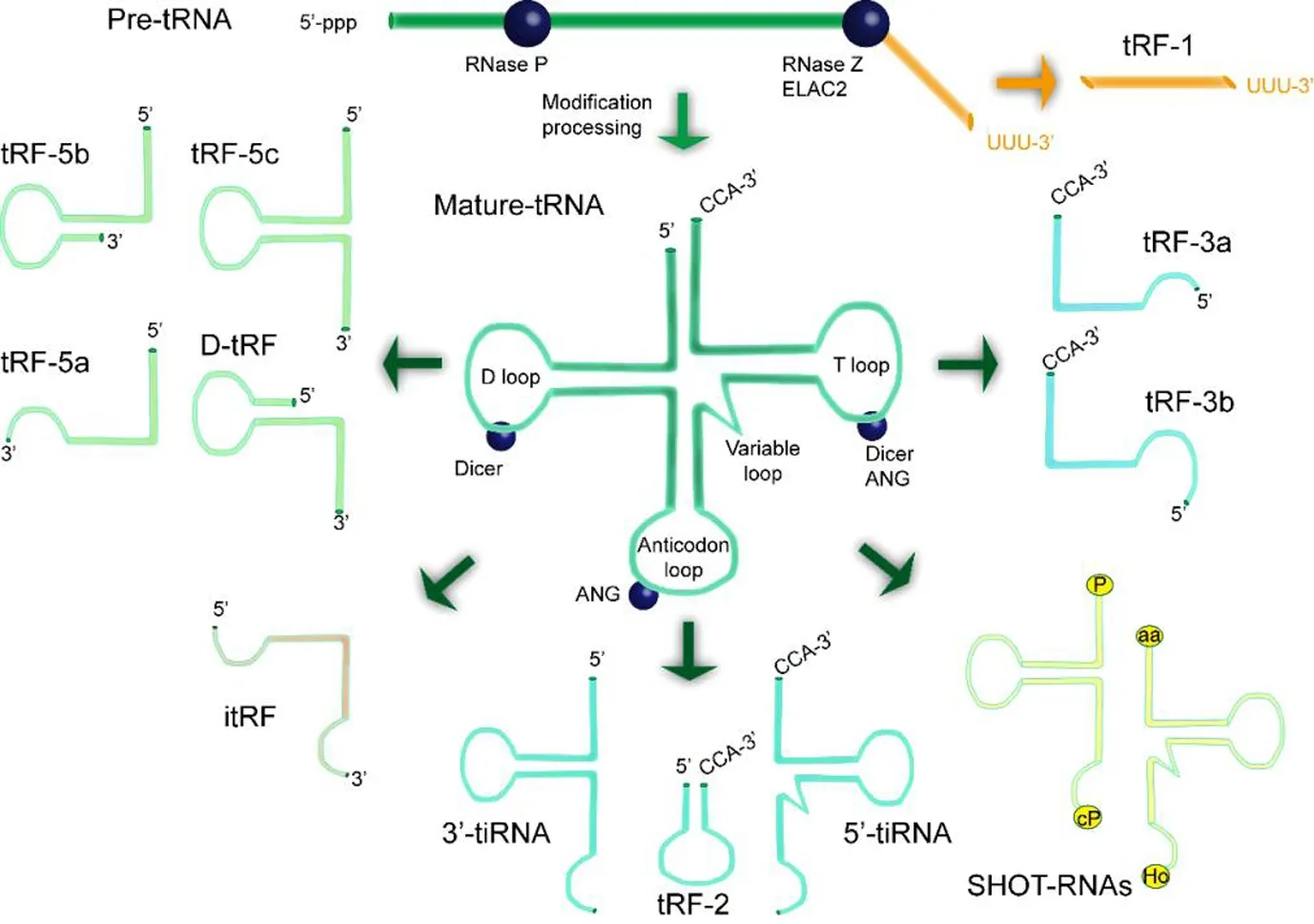
Figure 1 Biogenesis and main classifications of tsRNAs, which are generated during pre-tRNA processing or from mature tRNAs,including tRF-1, tRF-2, tRF-3, tRF-5, D-tRF,itRF, 3′-tiRNA, 5′-tRNA, and SHOT-RNAs.
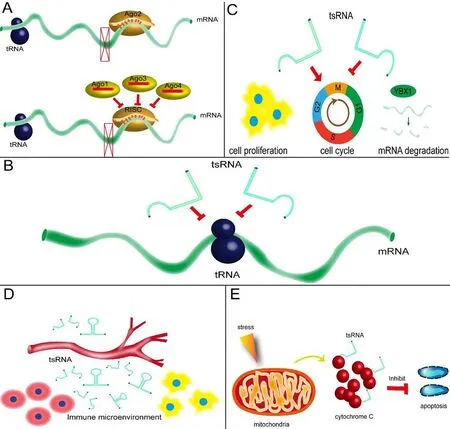
Figure 2 Diverse functions of tsRNAs.
Some tsRNAs, as well as being able to act as miRNAs in gene expression regulation, have been shown to interfere with protein translation by decreasing translational efficiency.First, one study found that tRF-5 decreased the expression levels of reporter genes without binding the complementary sites of target mRNA, which implies that tRF-5 acts via a different mechanism than that of miRNAs (Sobala and Hutvagner, 2014); several other studies have reached similar conclusions. For example, in Haloferax volcanii, tRFs, also known as Val-tRFs, with a length of 26 residues, directly bind to small ribosomal subunits both in vitro and in vivo, thus inhibiting protein synthesis by decreasing peptidyl transferase activity (Gebetsberger et al., 2012). In another study,several specific tsRNAs were shown to promote the activity of many eukaryotic initiation factors from both capped and uncapped mRNA, leading to translational inhibition (Ivanov et al., 2011). Among these tsRNAs, tsRNA-AlaUGCand tsRNA-CysGCAwere dominant in regulating protein translation,which likely occurs via the binding of translational repressor proteins (vanov et al., 2014). Furthermore, Yamasaki et al.(2009) could decrease translation levels by transfecting naturally occurring 5′ tiRNAs into the human osteogenic cell line U2OS. The authors also found that several structures called terminal oligoguanine motifs, located at the ends of 5′ tiRNAs, served as key factors in the repression of protein translation (Yamasaki et al., 2009). Finally, in terms of neurological disorders, it is reported that specific tsRNAs directly target the RNA binding protein, YBX1, and prevent its binding to mRNA targets, which may have a role in neurological diseases, such as increasing inflammation (Goodarzi et al.,2015). Taken together, these results suggest that tsRNAs may plausibly inhibit protein biosynthesis by targeting translation machinery. However, more studies are needed that focus on the details of these mechanisms.
Regulation of various cellular activities
To date, a large number of studies have demonstrated that some tsRNAs can regulate various cellular activities. For instance, tRF-1001 was reported to regulate cell proliferation and the cell cycle, and was shown to reduce cell viability in the HCT116 cell line (Lee et al., 2009). Another tRF-5,known as CU1276, showed a similar proliferative inhibition function in B cell lymphoma (Maute et al., 2013). One publication has reported the identification of a set of 5′-tsRNAs that is upregulated in differentiating mouse embryonic stem cells (Krishna et al., 2019). In terms of cell growth, the Tetrahymena thermophila Ago/Piwi protein Twi12 has been shown to be required for growth in the nucleus. Twi12-bound small RNAs, a type of tRF-3, contain modified bases and lessen base pairing to targets. One study highlighted the role of an Ago/Piwi protein and tRF-3 in nuclear RNA metabolism and cell growth (Couvillion et al., 2012). RNA stability is another important regulator of cellular activity. In PC-12 cells, a type of rat neuronal cell culture widely used as a model of nerve injury, a large upregulation of tiRNA generation by oxygen-glucose deprivation was identified, and some specific tsRNAs were found to regulate apoptosis and cell viability after cell damage, suggesting that these tsRNAs could be used as a potential marker of cell damage or an indicator of treatment effect in this type of injury (Elkordy et al., 2019). In addition, in breast cancer cells, tRFs derived from tRNAGlu, tRNAAsp, tRNAGly, and tRNATyrwere found to repress the stability of multiple oncogenic transcripts by displacing their 3′ untranslated regions from the RNA-binding protein YBX1 upon induction, thus reducing their stability and promoting degradation (Goodarzi et al., 2015).
Immune mediation
Recently, studies have shown that several tsRNAs can exert a critical influence on the immune response under certain stress conditions. For example, tRFs have been found not only in hematopoietic and lymphoid tissues, but also in blood circulation. During the acute inflammatory stage,tRFs are significantly upregulated in blood circulation, suggesting a potential function in immune regulation (Dhahbi et al., 2013; Zhang et al., 2014). In another study, 5-tRF that was derived from tRNAGlu, which is abundantly expressed in human monocytes, significantly inhibited the transcription of CD1a by forming a complex (Zhang et al., 2016). Another study identified that tsRNAs can directly interact with Tolllike receptors to activate immune responses in Th1 and toxic T lymphocytes (Wang et al., 2006). However, in part because limited numbers of studies have investigated these phenomena, the underlying mechanisms remain unclear.
Responses to stress conditions
A number of recent studies have shown that tiRNA expression is dysregulated under stress conditions (Emara et al., 2010; Goodarzi et al., 2015 Weitzer et al., 2015). Under hyperosmotic stress conditions, specific tiRNAs (including both 5′ tiRNAs and 3′ tiRNAs) can combine with mitochondria-released cytochrome C to form a ribonucleoprotein complex that suppresses apoptosis, including apoptosome formation and activity. Thus, these tiRNAs can be considered a type of novel anti-apoptotic regulator. Angiogenin is also a secreted RNase that cleaves tRNA to initiate a stress-response program in mammalian cells. The arseniteand pateamine A-induced assembly of stress granules can be promoted, and protein synthesis can also be initiated with the help of angiogenin, which is related to cell apoptosis and virus infections. In addition, in contrast to natural 3′ tiRNAs,5′ tiRNAs are capped with a 5′ monophosphate, which is required for the optimal assembly of stress granules, indicating that the assembly of stress granules is a component of the angiogenin- and tiRNA-induced stress-response program(Emara et al., 2010). In another study, the generation of a valine tRNA-derived fragment was reported in the halophilic archaeon H. volcanii under certain stress conditions. The function of this specific tRF is to bind to the small ribosomal subunit. As a result, global translation can be weakened in vivo and in vitro (Gebetsberger et al., 2017). Furthermore,when transfected into cells, angiogenin-dependent tsRNAs increased motor neuron survival upon exposure to stress,suggesting that tsRNAS can confer neuroprotection (Ivanov et al., 2014). Several studies have also linked cellular stress(Goodarzi et al., 2015; Elkordy et al., 2018), tRNA modification networks, and angiogenin-mediated production of tsRNAs to neurodevelopmental disorders.
Functional Roles of tRNA-Derived Small RNA in Neurological Diseases
It is well known that neurodegenerative diseases are caused by both genetic and environmental factors. As an increasing number of tRF and tiRNA have been discovered, the abnormal expression of tsRNA has been shown to be strongly associated with the development of various diseases, such as tumors, the cardiovascular system, epigenetics, and neurological diseases, all of which have increasingly concerned scientists in recent years. Developmental studies of tsRNA in neurological disorders have also been undertaken, the results of which are shown in Table 1.
Links to neurodegeneration
Well-established causes of neurodegeneration include abnormalities in tRNA metabolic pathways and tRNA processing enzymes. A series of studies revealed the founding mutation in CLP1 in four independent pedigrees (Hanada et al., 2013;Karaca et al., 2014; Schaffer et al., 2014; Weitzer et al., 2015).Specifically, this mutation is considered to include an important component of the tRNA splicing endonuclease complex, which is reported to be strongly associated with disorders of lower spinal motor neurons. Notably, homozygous missense mutations of CLP1 have been identified in patients with severe motor-sensory deficits, cortical dysgenesis, and microcephaly, which is also associated with the dysregulation of tRNA splicing. These defects in tRNA processing that lead to tRNA fragment accumulation may trigger the development of neurodegenerative diseases.
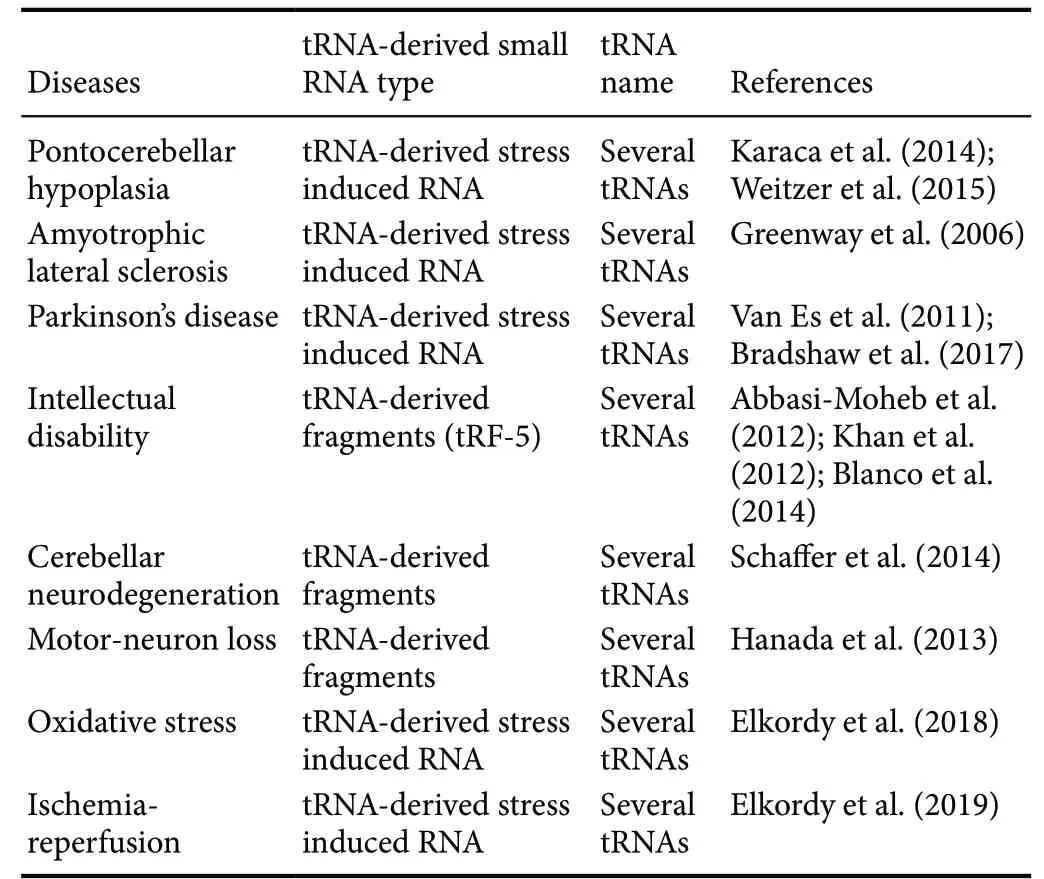
Table 1 Experimental evidence regarding tsRNA in neurological disorders
Over the past 20 years, more than 40 angiogenin mutants have been identified in neurodegenerative diseases, including amyotrophic lateral sclerosis and Parkinson’s disease.These mutations mostly affect the ribonucleolytic activity of angiogenin (Greenway et al., 2004). Significantly, ribonucleolytic activity regulates the production of tiRNAs. As a result,it is reasonable to speculate that certain tiRNAs are involved in the pathophysiological processes of these diseases. For example, tiRNAAlaand tiRNACysand their DNA analogs with G4 motifs may promote neuronal survival under stress conditions and provide new treatments for neurodegenerative disorders in the future (Li et al., 2018a).
The transfection of a particular tRF-5 into patient-derived neurons resulted in a reduction in cell survival under oxidative stress in vitro (Schaffer et al., 2014). In another study,microcephaly and other neurological abnormalities found in mice and patients were shown to likely be caused by mutations in cytosine-5 RNA methyltransferase NSun2. In NSun2-deficient mice, the accumulation of a certain tRF-5 caused by the loss of cytosine-5 RNA methylation decreased protein translation rates and activated stress pathways in cortical, hippocampal, and striatal neurons. Thus, the authors concluded that the aberrant methylation of NSun2-mediated tRNA linked cellular stress to neurodevelopmental disorders via stress-induced RNA cleavage (Blanco et al., 2014). In addition, other studies demonstrated that mutations in NSun2 cause a Dubowitz-like syndrome and a syndromic form of intellectual disability in humans (Abbasi-Moheb et al., 2012;Khan et al., 2012; Martinez et al., 2012). Other neurological diseases such as Parkinson’s disease (Van Es et al., 2011;Bradshaw et al., 2017) and amyotrophic lateral sclerosis(Greenway et al., 2006) have also been reported to have a close relationship with abnormal tRNAs and tsRNAs.
These results show that abnormal tRNA metabolism resulting in the dysregulation of tsRNAs is possibly associated with neurodegeneration. New therapies targeting these mechanisms may be developed in the future.
Links to neurotrauma
Aside from neurodegenerative diseases, neurotrauma is another subgroup of neurological diseases (Jiang et al., 2019;Wofford et al., 2019). However, the volume of research into this area is limited. Elkordy et al. (2018, 2019) reported that in PC-12 cells, tRNA cleavage and tiRNA generation were initiated under stress conditions induced by arsenite and hydrogen peroxide or ischemic reperfusion. Notably, the authors stated that the generation of tiRNAs occurred before severe cell damage, indicating their potential role as novel biomarkers of cell damage. In addition, tiRNA generation was angiogenin-mediated in these experiments. However,before these findings can be validated, studies focusing on the regulatory mechanisms of tsRNAs in neurotrauma need to be conducted both in vivo and in vitro.
Challenges and Perspectives
There are several questions about tsRNAs that have not yet been answered. First, the potential functions of tsRNAs and their underlying mechanisms, as well as the biogenesis of most tsRNAs in neurological disorders, should be clarified.Although several functions of specific tsRNAs have been identified in neurological diseases, they are still far from being able to be used as novel diagnostic and therapeutic targets in the clinic. Second, there is an urgent need to be able to correctly quantify tsRNAs. Currently, high-throughput sequencing and microarrays are commonly used for screening,while quantitative reverse transcription-polymerase chain reaction, SYBR staining, and immuno-northern blotting are used to detect certain tsRNAs. However, a combination of methods will likely need to be developed to specifically and sensitively quantify tsRNAs. Third, the exact relationship between tsRNAs and miRNAs needs to be deduced. Because of their similar lengths, non-coding features, and ability to form a combination with AGO proteins, they may have a similar function in regulating gene expression by inhibiting target mRNAs. However, their preferences for AGO proteins are different, suggesting that tsRNAs and miRNAs interact with each other. For example, a miRNA and a tsRNA have both been identified to inhibit one specific target mRNA. However, it is unknown whether these two types of RNAs act alone,mutually compete, or mutually promote one another.
Although there is a significant amount of knowledge still to be discovered in the field of tsRNAs, mounting evidence has shown that the abundance and composition of tRFs are related to disease types. A majority of tsRNAs are produced under stress conditions, and this pathophysiological environment is found in neurodegeneration and neurotrauma.As a result, it is reasonable to speculate that various tsRNAs may be involved in neurological disorders.
Concerning the diagnostic and therapeutic potential of targeting tsRNAs in neurological disorders, we must take the following aspects into consideration. First, as a diagnostic and therapeutic marker, tsRNAs should be easily acquired in the clinic. As an example, several tsRNAs are found more often than miRNAs in exosomes within body fluid (Vojtech et al., 2014). Thus, they have a strong potential to be invasive biomarkers of neurological disorders in the near future. Second, it is necessary to be able to clearly identify the roles of tsRNAs in neurological disorders. Screening and selecting a target tsRNA after neurological disease is quite important.For example, in an in vitro trial, after oxygen-glucose deprivation damage imitating stroke, significant changes were identified in several tsRNAs, indicating a potential role in cell damage (Elkordy et al., 2018). In addition, our recent in vitro study showed differential expression of tsRNAs in neurotrauma. These results strongly suggest that tsRNAs have functional roles in neurological disorders. Finally, the mechanisms for the therapeutic effects of tsRNAs in neurological disorders need to be understood before their clinical application. Although the underlying mechanisms remain to be clarified, we have reviewed many molecular experiments of tsRNAs in neurological disorders in the present paper, and we are therefore confident that this challenge will be resolved in the near future.
In terms of therapeutic approaches of tsRNAs in neurological diseases, no tsRNAs have yet been identified to be developed for clinical treatment. In our opinion, we should be prudent and conduct more studies in vivo and in vitro before clinical trials are performed. Generally speaking, in vivo,overexpression, or knockout of target tsRNA techniques could be used to verify their functions in disease processes.Next, in vitro, CRISPR, or genome editing techniques could be used to observe the beneficial effects and unwanted side effects of target tsRNA treatment in animal models. Finally,in a clinical trial, AAV or other advanced viral vectors that are harmless to humans could be adapted for treatment.However, this approach will likely require multiple attempts.
Identification of the functional versatility of tsRNAs will provide valuable insights into the physiology and pathophysiology of neurological disorders, thus offering new targets for future clinical treatments.
Acknowledgments:authors would like to thank Hao Feng and Yi Liu from China Rehabilitation Research Center for their cooperation in the professional consultation.
Author contributions:Original draft preparation: CQ; manuscript review and editing: CQ and PPX; figure design: CBL and CZ; table design:DGY and XZ; supervision: FG; project administration: MLY and LJD;study concept: JJL. All authors approved the final version of the paper.
Conflicts of interest:The authors declare that there are no conflicts of interest associated with this manuscript.
Financial support:This work was supported by the National Natural Science Foundation of China, No. 81870979 (to JJL), No. 81271366 (to MLY); the National Key R & D Program of China, No. 2018YFF0301104(to JJL); the Special Fund for Basic Scientific Research of Central Public Research Institutes of China, No. 2018CZ-1 (to JJL); the Basic Scientific Research Foundation of China Rehabilitation Research Center, No.2018ZX-30 (to FG); the Scientific Research Foundation of CRRC, No.2012C-1 (to JJL); the Major Science and Technology Project of Beijing of China, No. D161100002816004 (to JJL); the Special Capital Health Research and Development of China, No. 2018-1-6011 (to JJL).
Copyright license agreement:The Copyright License Agreement has been signed by all authors before publication.
Plagiarism check:Checked twice by iThenticate.
Peer review:Externally peer reviewed.
Open access statement:This is an open access journal, and articles are distributed under the terms of the Creative Commons Attribution-Non-Commercial-ShareAlike 4.0 License, which allows others to remix, tweak,and build upon the work non-commercially, as long as appropriate credit is given and the new creations are licensed under the identical terms.
Open peer reviewer:Ahmed Majeed Al-Shammari, Mustansiriyah University, Iraq.
Additional file:Open peer review report 1.
杂志排行

中国神经再生研究(英文版)的其它文章
- Ethanol extract from Gynostemma pentaphyllum ameliorates dopaminergic neuronal cell death in transgenic mice expressing mutant A53T human alpha-synuclein
- Peripheral nerve injury induced changes in the spinal cord and strategies to counteract/enhance the changes to promote nerve regeneration
- Genetic targeting of astrocytes to combat neurodegenerative disease
- Applications of advanced signal processing and machine learning in the neonatal hypoxic-ischemic electroencephalography
- Protective effect of hydrogen sulfide on oxidative stress-induced neurodegenerative diseases
- Current status and future prospects of stem cell therapy in Alzheimer's disease