云杉碳氮化学计量比对土壤水分和氮有效性的响应
2020-09-17谢路路胡雪凤刘庆华王丽霞尹春英
谢路路, 胡雪凤, 刘庆华, 王丽霞, 尹春英,*
1 中国科学院成都生物研究所,中国科学院山地生态恢复与生物资源利用重点实验室, 生态恢复与生物多样性保育四川省重点实验室 成都 610041 2 中国科学院大学, 北京 100049
生态化学计量比已经广泛应用于陆地森林生态系统中,不仅用于研究森林生态系统的演替与衰退、评估土壤养分的有效性等[1-2],还用于探究森林生态系统C、N、P等营养元素的吸收、利用以及分配过程等,对于揭示森林生态系统物质循环过程的影响因素及其作用机制具有重要意义[3-4]。植物碳氮化学计量比是植物在对外界环境适应过程中所形成的自我调控策略,反映出植物的生长速率和植物对氮素的利用效率[5-6]。在特定生长环境条件下, 植物可通过权衡不同器官之间以及器官内的碳、氮分配来提高植物生态适合度,从而优化植物整体的生长发育过程[7]。目前,植物碳氮化学计量比对外界环境变化的响应成为生态学领域的研究热点[8-9]。研究表明,植物碳氮比与土壤含水量有着显著的正相关关系,不同水分条件下,植物对于碳、氮的利用效率不同,影响了植物体内的碳积累过程和氮的分配格局,从而导致植物碳氮化学计量比变化[10]。马百兵等[11]以藏北高寒草地植物群落为研究对象,发现植物碳氮比随物种丰富度及降水、温度、干旱等外界环境因素而变化。近些年来,有关全球变化下森林生态系统碳氮化学计量比的研究已经涉及到碳氮循环[12-13]、养分固定[14]、森林更新与发育[15]等方面。
大气氮沉降和降水格局不均是全球气候变化的主要特征[16],对植物碳氮比有着直接或者间接的影响:首先,氮素随着降水或者降雪落到土壤中,改变了土壤有效氮含量[17];其次,降落到土壤中的氮素改变了土壤的理化性质如pH、酶活性等,影响了植物对氮素的吸收,从而改变植物的碳氮比[18]。此外,水分和氮素作为植物光合作用过程中必不可缺的物质基础,通过影响植物的光合作用和养分利用效率改变植物碳氮比[19]。洪江涛等[20]发现降水增加会提高土壤养分有效性和植物的净光合速率, 改变植物体内碳、氮的再分配, 从而影响植物碳氮比。因此,探究植物的养分吸收过程与植物的光合作用对大气氮沉降和降水的响应是了解植物碳氮比变化的重要生态生理过程。无机氮是植物根系吸收的主要氮源,但植物根系对土壤无机氮的吸收在不同土壤水分和氮有效性条件下呈现什么规律,如何影响植物的碳氮比,这些问题都未见报道。
西南亚高山地处青藏高原东部和长江上游,是典型的气候敏感区,同时也是IPCC报告中预测大气氮沉降的重要区域[21]。粗枝云杉(Piceaasperata)是该区针叶林的主要树种,在维持川西森林生态系统的养分循环和生态系统平衡与稳定方面有着不可忽视的作用[22]。目前有关川西亚高山森林生态系统碳氮化学计量比的研究多集中在单个环境因素变化下植株叶片、根系或者土壤等方面[23-25],对土壤水分和氮添加如何响应尚不清楚。研究该区云杉碳氮化学计量比对气候变化的响应对于川西森林生态系统的科学管理有着重要的作用。为此,本研究通过土壤水分和氮添加的两因素控制实验,以粗枝云杉幼苗为材料,研究植株及器官碳氮比对土壤水分和氮有效性的响应,并结合植物主要的碳、氮过程——光合作用及对无机氮的吸收,试图分析碳氮比变化的主要生理生态机制。
1 材料和方法
1.1 试验样地
试验在中国科学院成都生物研究所茂县山地生态系统定位研究站(31.70°N, 103.90°E, 海拔1826 m)进行。该站属典型的暖温带亚高山季风气候,其年平均气温、年平均降水量和年平均蒸发量分别为9.3℃、919.5 mm和795.8 mm。土壤pH处于5.8—6.0范围内,酸性棕壤土和褐土,云杉是该区针叶林的主要树种[26]。
1.2 试验材料及设计
试验于2013年10月布置:取该区域典型针叶林的林下土壤,过筛后混匀,装入高45 cm、内径42 cm的塑料花盆中。土壤pH 6.75,有机质和全氮含量分别为 27.5、2.2g/kg,硝态氮和铵态氮含量分别是19.1 mg/kg和3.1 mg/kg。从当地苗圃选取生长一致、生长良好、无病虫害的3年生云杉幼苗,移栽装好土的花盆中,每盆1株。待云杉幼苗生长稳定,于2014年5月开始进行土壤水分和氮添加的控制试验,水分控制和氮添加为每年的生长季(5—9月),具体过程参见等Yin[27]的方法。
试验采用两因素(水分×氮素)随机区组设计。根据当地测定的旱季极端含水量和前期的预试验结果(低于40%土壤田间持水量几乎不能维持云杉的成活和生长),试验设置5个水分梯度:分别是土壤田间持水量的40%(W1)、50%(W2)、60%(W3)、80%(W4)和100%(W5),对应的土壤重量含水量分别为11.99%、14.99%、17.99%、23.98%、29.98%,其中W5为对照,另外的土壤水分梯度代表常见的土壤水分或不同程度的干旱处理。根据国际上关于氮沉降的研究,控制实验的氮沉降最高水平是34 gN m-2a-1[28],并结合本地区的相关研究[29],设置3个施氮浓度:0(N0)、20(N1)和40(N2)gN m-2a-1,其中N0为对照,N1、N2代表不同浓度的氮沉降。共15个处理,每处理15盆。
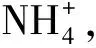
1.3 试验方法
水、氮控制试验开展5年后,于2018年8月从每处理随机选择5株云杉进行样品的采集、分析测试和收获,具体方法如下。
1.3.1光合速率的测定
在生长季中期,选择晴朗无云的天气,进行云杉叶片光合速率的测定,连续测定三天。在每处理中,随机选择5株云杉,每株选取相同部位、当年生完全展开的3—5簇针叶,用LI-6400XT便携式光合仪(Model LI-6400,Li-Cor.,Lincoln,NE,USA)进行测定。测量具体时间在2018年8月中旬9:00—11:00和13:30—16:30,选用针叶叶室(Li-Cor 6400-05)和LED红蓝光源于控制环境条件下完成测定:叶室温度是24—26 ℃,叶室内相对湿度是36%—55%,设置光强为1200 μmol m-2s-1,CO2浓度是400 μmol mol-1,具体过程参考Yin等[27]方法。测量结束后将测量的云杉叶片收集并带回实验室测定叶面积(WinRHIZO扫描分析仪,Régent instruments, Quebec, QC, Canada),得到投影面积之后带入到LI-6400XT便携式光合仪系统自带的计算公式中,得到单位叶面积的光合速率(Pn)[32]。
1.3.2根系离子流速测定

1.3.3碳和氮含量测定
将上述收获的根、茎、叶样品,于110 ℃杀青15 min,70℃烘干至恒重。采集的土壤(过2 mm筛),带回实验室于4℃冰箱保存。将烘干的根、茎、叶利用研钵粉碎、混匀,过100目筛,置于50 ℃烘箱中处理2 h后准确称取40 mg样品,用锡箔纸包好,用元素分析仪(PE-2400 II)测定总碳和总氮含量。土壤有效氮含量采用AutoAnalyserIII流动分析仪(SEAL Analystical, 德国)进行测定。准确称取10 g鲜土,置于50 mL 2 mol/L的KCL浸提液中,混合均匀,于25 ℃室温下,震荡器震荡30 min(震荡频率180±20 r/min)后上机测定。
1.4 统计分析
2 结果与分析
2.1 土壤水分和氮添加对云杉不同器官碳和氮含量的影响
相同土壤水分和氮添加下,根的碳含量((373±45)g/kg)显著低于叶片((496±9)g/kg)(P<0.05)和茎的碳含量((491±5)g/kg)(P<0.05),茎和叶之间的碳含量相近(P>0.05)(图1)。土壤水分对根和叶的碳含量有显著影响(P<0.05),随土壤水分有效性的降低,根和叶的碳含量逐渐升高;茎的碳含量未受到土壤水分的显著影响(P>0.05)(图1)。各器官的碳含量均受到氮添加的显著影响(P<0.05)。氮添加显著提高了根的碳含量,氮添加浓度越高,提高幅度越大;随氮添加浓度的增加,茎和叶的碳含量逐渐降低。土壤水分和氮添加对于器官碳含量的影响均不存在互作效应(图1)。
氮在不同器官的分配表现为:叶的氮含量((13.1±1.3)g/kg)显著高于根((10.9±0.6)g/kg)(P<0.05),茎((7.3±0.7)g/kg)最低(P<0.05)(图1)。根和茎的氮含量受土壤水分和氮添加的显著影响(P<0.05)。根和茎的氮含量随着土壤水分有效性的降低而逐渐增加。氮添加提高了根和茎的氮含量,氮添加浓度越大,氮含量越高。土壤水分和氮添加的交互作用显著影响了茎的氮含量(P<0.05)(图1)。云杉叶片氮含量仅受到氮添加的影响,氮添加提高了叶的氮含量(图1),一般而言,随着氮添加浓度的增加而逐渐增加,呈现N2>N1>N0的趋势。
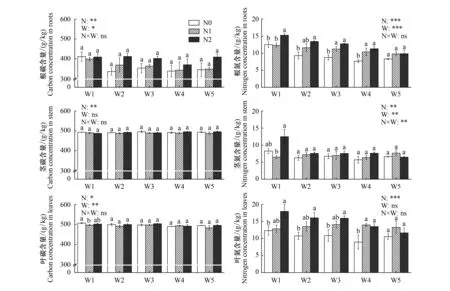
图1 云杉根碳含量、氮含量,茎碳含量、氮含量,叶碳含量、氮含量对土壤水分和氮添加的响应(平均值±标准误)
2.2 土壤水分和氮添加对云杉各器官碳氮比的影响
云杉茎的碳氮比(70.5±6.9)显著高于植株(45.3±3.6)(P< 0.05)和叶的碳氮比(40.2±4.2)(P<0.05),根碳氮比(34.9±1.8)最低(P<0.05)(图2)。植株和茎的碳氮比受土壤水分和氮添加的显著影响(P<0.05)(图2)。随土壤水分有效性的降低,植株和茎的碳氮比呈现先升后降的变化,W4处理下达到最大。氮添加降低了植株和茎的碳氮比,氮添加浓度越高,植株和茎的碳氮比越小。根的碳氮比受到土壤水分、氮添加及其交互作用的显著影响(P<0.05)(图2)。根碳氮比随土壤水分有效性的降低而减小,氮添加的效应因土壤水分有效性的不同而不同:W1、W3和W4处理下,氮添加显著降低了根碳氮比,N2处理下的根碳氮比显著低于N0和N1;其他水分处理下(W1和W5),不同氮添加浓度之间没有显著差异。叶碳氮比仅受到氮添加的显著影响(P<0.05)(图2),氮添加显著降低了叶碳氮比。随着氮添加浓度增加,叶碳氮比逐渐降低,呈N0>N1>N2的趋势。
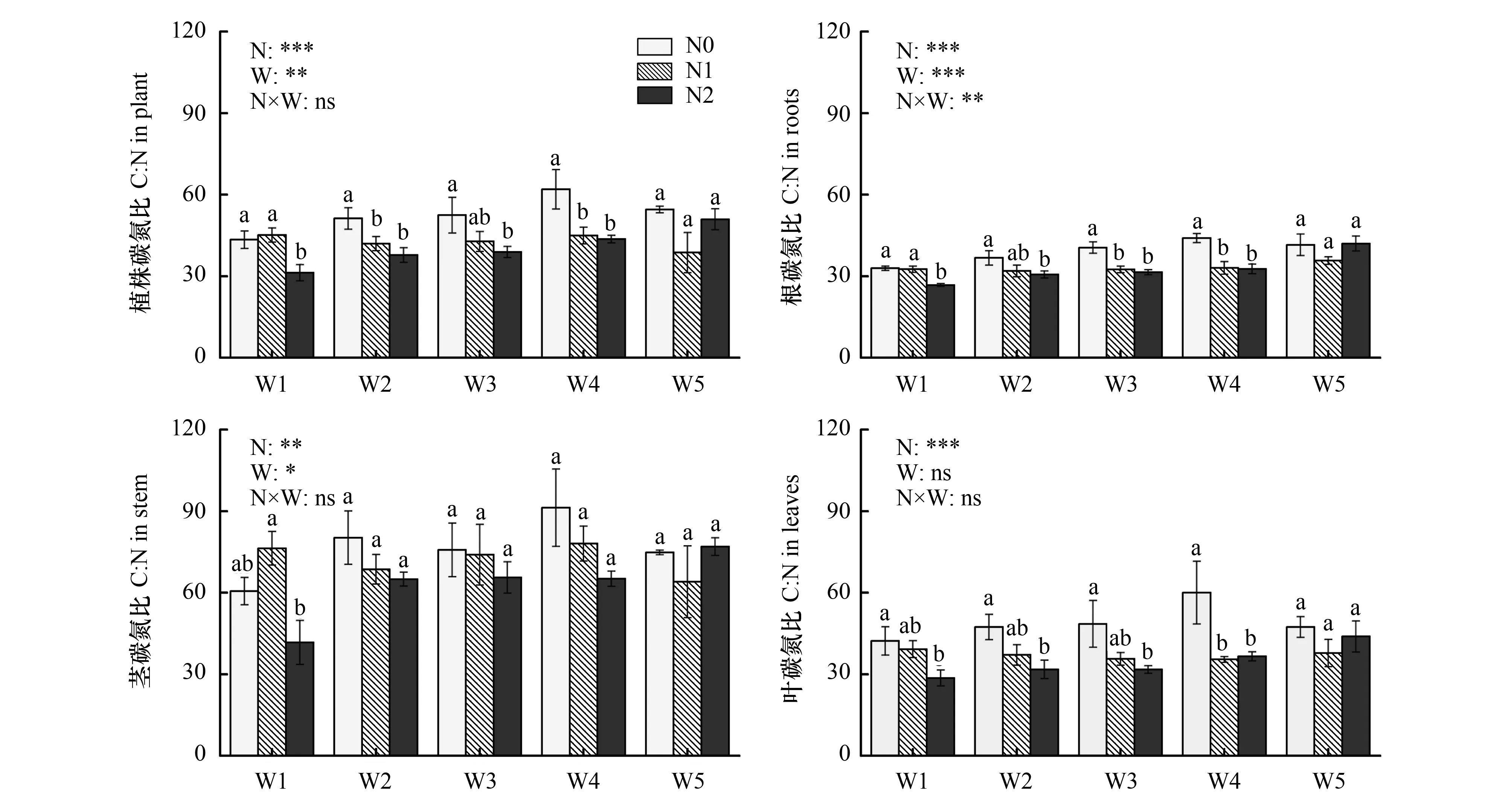
图2 云杉植株碳氮比、根碳氮比、茎碳氮比、叶碳氮比对土壤水分和氮添加的响应(平均值±标准误)
2.3 土壤水分和氮添加对云杉净光合速率(Pn)的影响
土壤水分、氮添加及其交互作用显著影响Pn(P<0.05)(图3)。随土壤水分有效性的减小,Pn先升后降,在W4时达到最大。氮添加对Pn的影响因土壤水分有效性而异,土壤水分低于W5时,同一土壤水分下,Pn随着氮添加浓度增加而增加,N2显著高于N1和N0,在N2W4达到最大,且N1和N0处理之间没有显著差异。W5处理下表现为:N2>N0>N1(图3)。
2.4 土壤水分和氮添加对云杉根系氮素吸收速率和土壤有效氮的影响
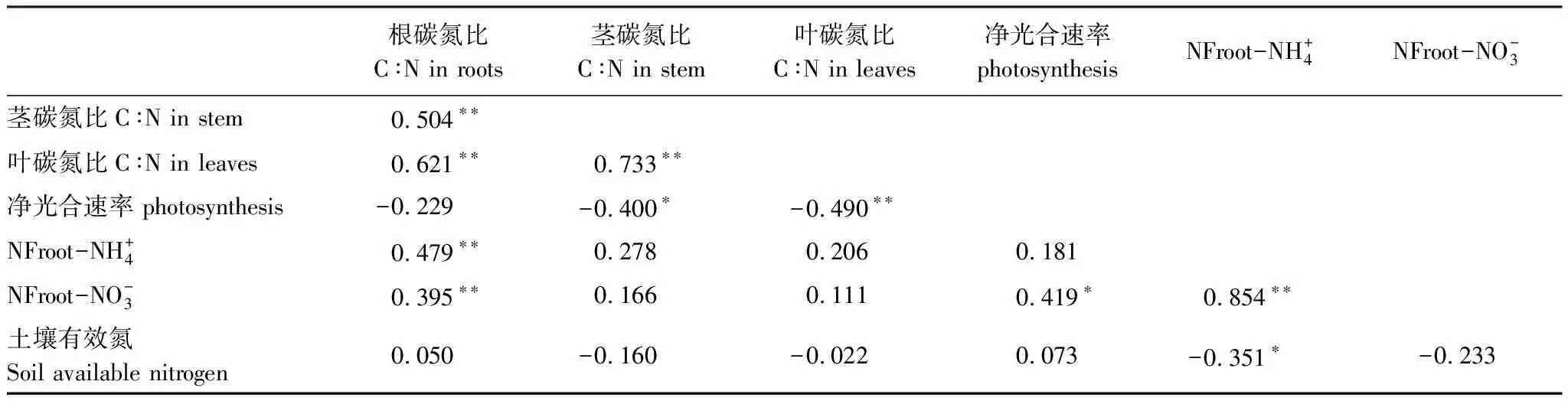
土壤有效氮含量受到土壤水分、氮添加及其交互作用的显著影响(P<0.05)(图3)。土壤水分有效性的降低显著提高了土壤有效氮含量,W1和W2处理下的有效氮含量显著高于其他水分处理。氮添加的效应与土壤水分有效性密切相关:W1时,氮添加显著降低了土壤有效氮含量,呈N2 图3 云杉根对 的净吸收速率、根对 的净吸收速率、土壤有效氮含量和净光合速率对土壤水分和氮添加的响应(平均值±标准误) 植物体内的碳、氮含量不仅与其长期进化过程中的结构有关,还与植物生长速率和养分利用效率密切相关[36]。一般来说植物叶片氮含量越高,其光合速率和生长速率越高,对资源的竞争能力也高,而叶片碳含量高则说明其具有大的比叶重,弱的光合速率和生长速率,但有较强的抵抗不利环境的防御能力[37-38]。研究表明氮添加浓度的增加促进了云杉叶片氮含量的增加,从而提高净光合速率、生长速率,并提高叶对于资源的竞争能力[37];但叶碳含量随氮添加浓度增加而降低,导致叶碳氮比和养分利用效率的下降,说明过量氮添加虽然能够提高叶的生长速率和资源竞争能力,但不利于叶的养分利用效率。石贤萌[39]等研究也有类似结论,过量氮添加提高了植物器官氮含量,尽管满足了植物生命活动所需的营养元素以提高生长速率,但降低了各器官碳氮比,导致植物养分利用效率的降低。此外,与对照W5相比,叶碳含量随土壤水分有效性减小而增加,且叶碳含量显著高于根和茎(P<0.05),结果表明在应对干旱环境时,植物为了增强对不利环境的抵御能力,会将相对多的碳留在叶片,从而减缓了叶片碳同化过程,叶碳含量高则说明生长速率缓慢[38],减小对水分的消耗,导致养分利用效率的降低[40]。相同的碳、氮分配规律也适用于植物根系,研究表明,植物根系在干旱下会分配到更多的碳[41],根碳含量的增加能够提高抵抗外界不良环境的能力,缓解植物生存压力[38]。例如,干旱下根系的形态和生理特征会发生变化,通过提高根冠比、增加菌根真菌的侵染或加快内源激素的产生等方式提高水分和养分吸收能力以应对干旱胁迫[42-43]。综上所述,植物体在应对土壤水分和氮有效性变化时,通过调整体内碳、氮含量分配,改变植物的生长速率和养分利用效率以更好的适应其生存环境。 植物碳氮比依赖于碳、氮含量变化,不仅体现了植物应对外界环境变化时自身所做出的养分利用策略调整,也间接反映植物体的生长速率[44-45]。本试验结果表明,云杉植株及器官碳氮比受到土壤水分和氮添加的显著影响(P<0.05)。与对照W5相比,植株及器官碳氮比随土壤水分有效性的降低先升高后降低,在W4处理下达到最大,说明W4处理下的氮利用率最高,主要由于器官碳、氮含量虽然随土壤水分有效性的降低而升高,但不同器官的碳、氮含量的变化仍有显著差异(P<0.05),从而导致W4处理下的碳氮比最高。且过多的土壤水分不利于植物养分利用效率的提高。这与马百兵等[11]结果一致,他们在研究藏北高寒草地时发现植物碳氮比随降雨量的增加而降低,主要原因是增加的水分降低了植物的碳含量,却不影响氮含量,从而导致氮利用效率的降低。前人研究也有类似结论,即降水量的变化影响了植物体内碳、氮含量的再分配,从而改变碳氮比,以体现植物对养分利用的权衡[20]。例如,Lu等[46]研究发现在湿润环境中的适度干旱有利于提高植物的氮吸收速率,却降低植物生长速率,从而改变植物碳、氮含量,导致植物碳氮比的降低。此外,氮添加浓度的增加显著降低了云杉植株及器官的碳氮比,N2处理显著低于N0处理,原因是器官氮含量随氮添加浓度增加而增加,碳含量却降低,从而降低氮的利用效率,导致碳氮比的减小,这与之前的研究结果相同[47]。杨惠敏和王冬梅[48]研究也证明,与高养分条件相比,植物在低养分条件下虽然生长速率缓慢,但有较高的碳氮比,植物养分利用效率较高。综上所述,在低的土壤水分和养分有效性下,云杉有较高的碳氮比,即资源(特别是氮)的利用率高,过量降雨和氮添加会降低资源的利用率。 不同生境的植物通过改变其碳、氮含量分配和碳氮比以应对环境变化。在本区域的相关研究中,杨欢等[23]发现粗枝云杉生长季的叶片碳氮比显著高于非生长季,主要是生长季的碳含量高,氮含量低而导致氮的利用效率较高,该研究中生长季的叶片碳氮比平均值为28.5±1.2,与本试验中N2处理的平均值持平(32.1±3.0),却低于本研究所有处理的平均值(40.2±4.2),可能的原因本研究试验设计中的土壤水分干旱水平设置较多,导致留在叶片中的碳相对多,从而提高碳氮比。此外,赵维俊等[49]在祁连山探究青海云杉叶片化学计量比的研究也说明了这一点,即该区的水热条件影响了碳、氮分配,导致青海云杉叶片碳氮比平均值(22.95—36.72)低于本研究的结果。 本研究中的试验材料(云杉幼苗)均采购自当地苗圃,严格按照株高、基径等生长状况完全一致的要求进行挑选,以确保各处理中植株管理方式、年龄、生长、生理状态等的一致性。虽并未测定初始植株和各器官的总碳、总氮含量,但根据相关研究表明[50],同种植物在相同土壤条件、管理方式等条件下,其碳氮比是相对稳定的,在确保试验处理前各植株状况基本一致的基础上进行的,仅考虑实验因素(土壤水分和氮添加)的效应是可行也是可信的。 植物生长发育的两个重要关键过程是植物地上部分的光合作用(碳同化过程)和地下根系对营养物质的吸收及其向地上部的运输(氮过程),碳同化过程提供碳源和能量给氮过程,并依靠氮过程提供的酶和光敏色素,这两个过程密切联系并受到外界环境的影响[51]。本研究中土壤水分和氮添加显著影响了云杉叶片的Pn和根系养分吸收速率(P<0.05)。其中,云杉叶片Pn随土壤水分有效性的降低先升后降,随氮添加浓度增加而增加,在N2W4处理下达到最大,结果说明适宜的土壤水分和充足的养分有利提高植物生长速率和光合速率。郭昉晨[52]也有类似的结果,氮添加促进了植物叶片的吸光强度和光反应速率,提高植物的光合作用和蒸腾速率。植物的碳同化过程与氮过程密切相关,两者在其生存环境下的平衡与协调决定了植物的生长与品质[53]。研究表明云杉根系氮吸收过程也受到土壤水分和氮添加的显著影响(P<0.05)。氮添加浓度的增加在土壤水分有效性较低时抑制根系氮素速率,在土壤水分有效性相对高时促进根系吸收速率,W4处理下的氮吸收速率高于其他处理。这与孙誉育[54]等的研究结果一致,他们的研究表明低土壤水分有效性下的氮添加加剧了干旱胁迫,导致植物吸水困难;而充足土壤水分下的氮添加则提高根系活力和呼吸速率,从而促进植物对水分和养分的吸收。如上所述,与对照W5和N0相比,云杉的碳、氮过程均在W4处理及氮添加(N1和N2)下达到较高水平,说明适宜的水-氮配比有利于植物的碳、氮过程与平衡,从而促进植物的生长。Weybrew等[55]在研究不同水肥处理下的碳、氮代谢时也发现正确施肥和适量的水分条件有利于植物的生长与品质。 植物碳氮比反映出植物在不同环境下的养分利用权衡策略和植物控制碳水化合物的库-源关系[56-57]。研究表明云杉植株及器官碳氮比均在N0W4处理下达到最高水平,说明该处理下的氮利用效率高于其他处理,体现了碳同化过程与养分吸收过程之间的协调。与对照W5相比,低的土壤水分有效性促进了云杉叶的碳同化过程,增加了云杉叶和根的碳含量,说明在水分限制条件下,植物倾向于增加输送到植物重要器官的碳含量以保证植物的生长发育过程,即植物在不利环境下会调整养分分配,把资源优先提供给重要部位,体现了植物的生存策略[58];此外,氮添加浓度的增加显著促进了云杉叶片的碳同化过程和根的氮吸收过程,并提高各器官的氮含量,却降低了碳含量,说明与N0对照处理相比,氮添加浓度的增加导致植物在吸收氮的同时所同化碳的能力下降即降低了植物的氮利用效率。这与前人研究结果一致[48],说明植物在低养分条件下的养分利用效率较高。植物碳氮比与植物生长发育的两个重要过程密切相关,研究表明叶碳氮比与Pn存在显著负相关关系(P<0.01),可能的原因是植物叶片光合作用所同化的碳大部分留在叶片,分配给其他器官的碳含量降低,从而导致叶片碳同化过程减缓[38],而叶片中的氮是其进行光合作用的重要资源,叶片光合作用随叶氮含量增加而增强[59],叶片碳、氮含量的变化导致叶碳氮比与Pn呈负相关关系;而根系碳氮比与其养分速率之间存在显著正相关系(P<0.01),主要在于植物根接收来自叶片碳同化过程分配的碳,并利用这些碳完成其生命活动,碳含量越高则越有利于养分吸收,根碳含量促进其氮素吸收过程,而当根氮含量增加并满足植物生长需求时,根的养分吸收过程可能会减弱,氮素吸收速率随根氮含量增加而减缓,从而导致根碳氮比与氮素吸收速率之间的显著正相关关系。植物碳氮比的变化是植物碳同化过程与氮吸收过程的协调,体现了在生长过程中对植物对养分利用的权衡策略,通过改变碳、氮过程,调节植物生长以更好的应对外界环境变化。 表1 云杉各器官碳氮比与光合速率、根离子吸收、土壤有效氮含量之间的相关性 本研究通过探究云杉植株与各器官的碳氮化学计量比对土壤水分和氮有效性的响应过程,发现云杉碳氮比在不同器官之间存在显著差异,相同土壤水分和氮添加下,茎的碳氮比(70.5±6.9)显著高于叶的碳氮比(40.2±4.2)(P<0.05)和根的碳氮比(34.9±1.8)(P<0.05)。云杉各器官碳、氮含量及碳氮比受到土壤水分和氮添加的显著影响,与对照W5和N0相比,各器官碳含量随土壤水分和氮有效性减小而增加,各器官氮含量随氮添加浓度的增加而增加,各器官碳氮比则在N0W4处理下达到最高值,因此推断出云杉各器官的氮利用效率均在低土壤水分和氮有效性下较高,说明合适的水-氮配比有利于提高植物的生长速率和养分利用效率。不同土壤水分和氮添加下碳氮比的变化依赖于植物的碳同化过程和氮过程,其中,云杉净光合速率和氮素吸收速率均随土壤水分和氮有效性的增加而增加,但由于不同处理及器官的养分权衡策略不同,且植物在不利环境下倾向于将资源分配到重要部位,从而改变植物体的碳、氮分配,导致碳氮比的变化。 致谢:感谢中国科学院北京植物研究所唐波在非损伤微测技术方面的指导和帮助。
3 讨论
3.1 土壤水分和氮添加对云杉碳、氮含量及碳氮比的影响
3.2 土壤水分和氮添加对云杉主要碳、氮过程的影响
3.3 土壤水分和氮添加对云杉碳氮比的影响机制
4 结论
