社群等级对金乌贼行为表型及能量代谢的影响
2020-09-17柏青青李江涛徐秀文张宗航李文涛张秀梅
柏青青,李江涛,徐秀文,张宗航,李文涛,张秀梅,2,*
1 中国海洋大学海水养殖教育部重点实验室,青岛 266003 2 青岛海洋科学与技术国家实验室,海洋渔业科学与食物产出过程功能实验室,青岛 266072
由于环境资源的有限和空间的异质性,动物社群等级在自然界中广泛存在[1]。在社群等级形成过程中,个体之间产生竞争交互作用,形成优势个体和劣势个体[2]。优势个体在不同程度上控制食物源、占据中心领域位置、对处于劣势地位的个体发起主动攻击行为、优先进行交配活动等;而劣势个体处于长期压力状态,会减少自身的活动及对食物、领地、生殖等资源的竞争[3-4]。社群等级的差异导致了个体间能量代谢、激素、基因水平的差异[5-7]。一般来说,优势个体争斗能力较强,在激烈争斗阶段的能量主要由无氧呼吸提供[8-9],而当个体处于慢速游泳状态或者悬浮状态时,主要通过有氧呼吸提供能量[10-11]。社群等级形成过程中不同个体之间的竞争交互作用对于亲体繁育及子代的生长发育具有显著影响[12]。有研究表明在繁殖过程中,雄性个体之间常通过打斗来获得交配权,但频繁的争斗行为也导致了交配成功率的下降,从而影响繁殖效率[13]。因而研究社群等级胁迫条件下,繁殖亲体的行为表型及其能量代谢机制对于动物繁殖行为的人工调控及子代健康苗种培育具有重要意义。关于繁殖期间个体优势等级的研究较少,仅见Jeffrey和Gilmour[12]等报道,处于劣势地位的雌性斑马鱼压力激素水平高,受下丘脑—脑垂体—肾间组织(Hypothalamic-pituitary-interrenal)轴基因调控且能够遗传到子代,从而影响子代的生长发育。金乌贼(Sepiaesculenta)也具有较强的领域行为[14],无论是与雌性交配还是护卫伴游,一旦有非护卫雄性闯入,护卫雄性立即做出反应,对非护卫雄性进行驱赶。处于优势地位的大规格雄性常常主动攻击小规格雄性[14]。然而关于社群等级对于金乌贼繁殖期行为表型以及能量代谢影响的研究尚未见报道。
金乌贼是我国近海经济价值较高的头足类之一,也是目前山东省重要的增殖放流物种。其世代周期短、产卵数量少的特性,使得金乌贼繁殖生物学的研究成为重点。目前已经开展了亲体培育与人工养殖模式[15-16]、繁殖行为和繁殖模式[17]等的研究,但繁育技术仍不成熟。在金乌贼人工繁育过程中,如亲体密度过高,个体间易产生激烈的竞争作用,形成社群等级。本文通过观察繁殖期金乌贼雌雄个体社群等级的形成过程,分析不同优势等级雌雄个体的行为表型以及能量代谢差异,探究行为表型与能量代谢的相关性,以期为金乌贼健康苗种培育以及规模化繁殖技术优化提供理论依据。
1 材料与方法
1.1 实验材料
2018年7月于青岛金沙滩金乌贼养殖基地开展实验,所用金乌贼亲体均采集于青岛薛家岛周边海域(35°59′N,120°19′E)。将采集的雌雄亲体分开暂养4 d,从中选取活力强、胴背部无损伤、色泽鲜明的雌雄亲本共48只(24♀24♂)备用,测量并记录实验亲本的胴背长和体重等形态学指标。亲体暂养采用砂滤流水方式,持续充氧,池内盐度30—32,温度20—21.5℃,pH值7.7—7.8,水深控制在80 cm。每天定时投喂鲜活的凡纳滨对虾(Litopenaeusvannamei)。由于雄性个体间的争斗喷墨,会导致水体能见度降低,每天9:00定时按50%比例换水1次。
1.2 实验设计
实验设置两个实验组,分别为两雄一雌组(2♂1♀)和两雌一雄(2♀1♂)组,每个实验组设置8个平行。实验开始前,在实验水槽中(0.9 m×0.9 m×0.9 m)按比例放入雌雄个体,为避免规格差异对优势等级形成的影响,在进行实验组配对时选择体长和体重差异小于5%雄性个体和雌性个体进行分组。(2♂1♀)组用于观察雄性之间的竞争;(2♀1♂)组用于观察雌性之间是否存在竞争关系。表1为实验组中实验个体的体重、胴长、性腺重、肝脏重等基本生物学参数。实验开始前将实验对象放置在水槽中适应12 h后进行实验,通过录像连续观察行为变化。
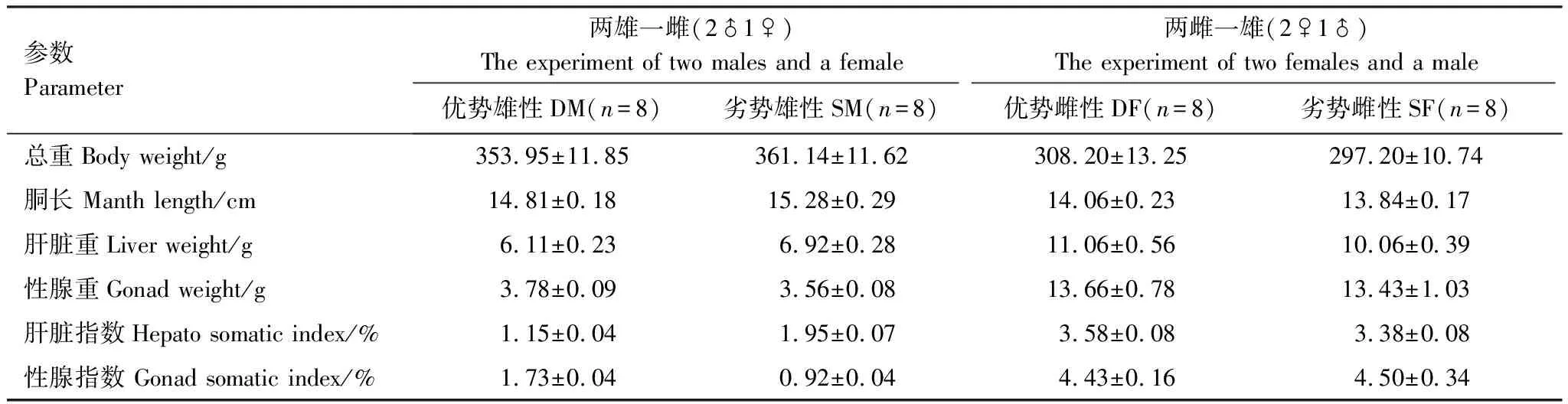
表1 实验组中金乌贼形态参数
1.3 行为分析方法
实验连续进行4 d,选择每天9:00—11:00以及13:00—15:00两个时间段录像进行行为观察。录像分析结果可将金乌贼的行为反应定义为两大类7种不同行为,包括主动攻击行为(攻击、追逐、取代、威胁),防御行为(喷墨、后退、逃逸)。参照Johnsson和Björnsson[18]、Filby等[19]的方法通过行为观测,计算平均每小时主动攻击和防御行为发生次数。主动攻击行为记1分,防御行为记-1分。根据个体得分状况判别雌雄个体的社群等级。
观察发现,雄性个体尝试交配数次后得以成功交配,通过观察尝试交配次数以及成功交配次数,可得出雄性金乌贼成功交配率。其中,雄性个体成功交配率=成功交配次数/尝试交配次数。而雌性个体在繁殖过程中,间歇性靠近产卵基放卵,统计不同等级雌性个体平均每小时放卵次数。
观察发现,实验水槽中的金乌贼通常处于悬浮状态,可将其分为游动悬浮、静止悬浮、静止伏底和争斗状态四种不同的状态,统计单位时间内不同运动状态的时间。
1.4 样品的采集与测定方法
连续4 d行为观察结束后,每组随机取出2个实验个体(共16♀16♂)用吸水纸吸取乌贼体表的水分,在解剖盘上进行活体解剖,取腕部肌肉和性腺组织进行超低温保存。准确称量各组织重量,按重量(g)∶体积(mL)=1∶9的比例加入9倍体积的生理盐水,冰水浴条件下机械匀浆,2500 r/min,离心10 min,制备成10%的匀浆上清液置于-20℃冰箱中保存待测。取得的上清液需进行总蛋白测定,测定方法采用标准牛血清蛋白作标准曲线的考马斯亮蓝染色法。己糖激酶(Hexokinase, HK)、丙酮酸激酶(Pyruvate kinase, PK)、乳酸脱氢酶(Lactate dehydrogenase, LDH)、柠檬酸合酶(Citrate synthase, CS)、苹果酸脱氢酶(Malate dehydrogenase, MDH)的活性和总蛋白、乳酸(Lactic acid, LD)浓度均采用南京建成生物研究所提供的试剂盒测定(A077-1、A076-1、A020-2、A108、A021-2、A019-2和A045-2),具体方法参照说明书。
计算每个实验个体有氧代谢酶(CS、MDH)和无氧代谢酶(HK、PK、LDH)含量以及无氧代谢产物乳酸(LD)含量。通过录像资料,对应行为分析结果,比较不同优势等级个体间有氧和无氧代谢的差异,进一步分析每个实验个体的运动状态与有氧和无氧代谢酶的相关性。
1.5 数据处理
数据为平均值±标准差。优势和劣势个体之间对比采用T-检验;不同实验组合之间对比采用单因素方差分析(one-way ANOVA)和Duncan两两比对。游动悬浮时间与有氧代谢酶含量以及争斗时间与无氧代谢酶活性之间的关系采用曲线估计进行分析,并用 ANOVA方差分析对回归的显著性进行检验。所有统计分析均采用SPSS 19.0。
2 结果
2.1 社群等级的形成
根据观察,放入同一实验水槽不同组合的金乌贼,其社群等级一般在2 d之后形成。如图1所示,社群等级形成初期个体间交互行为发生次数多,随着时间推移个体间交互行为次数逐渐降低,并达到稳定状态。雌性个体的防御行为次数无显著差异(P>0.05),无论是主动攻击还是防御行为的发生概率均低于雄性个体。

图1 金乌贼个体间交互行为及社群等级的形成过程
金乌贼具有明显的主动攻击行为和防御行为。经计算得到优势雄性和劣势雄性个体的攻击和防御行为得分分别为(11.8±1.98)、(-5.95±2.69)(P<0.05),优势雌性个体和劣势雌性个体分别为(1.48±1.22)、(-4.96±1.02)(P<0.05)(图2)。观察并记录雄性个体尝试交配次数与成功交配次数,结果发现优势雄性和劣势雄性个体的成功交配率分别为(28.27%±4.66%)、(11.94%±2.81%)(P<0.05)。优势雌性个体和劣势雌性个体放卵次数分别为(10.72±1.65)次/h、(1.85±0.68)次/h(P<0.05)。
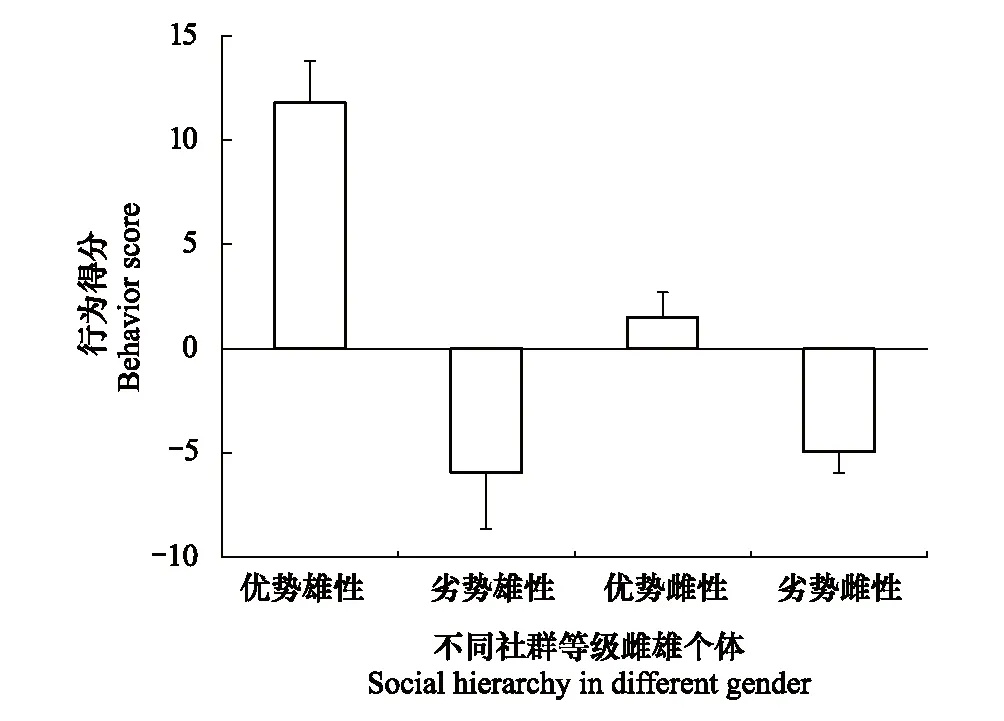
图2 金乌贼雌雄不同社群等级行为得分
2.2 不同等级金乌贼行为表型的差异
如图3所示,优势雄性个体游动悬浮时间和争斗时间显著高于劣势个体(P<0.05);与之相反,优势雄性个体静止悬浮时间和静止伏底时间显著小于劣势个体(P<0.05)。优势雌性个体游动悬浮时间显著小于劣势个体(P<0.05),静止伏底时间显著高于劣势个体(P<0.05)。雌性个体在繁殖期争斗行为极少发生,争斗时间和静止悬浮时间显示为优势和劣势个体二者之间无显著性差异(P>0.05)(图3)。
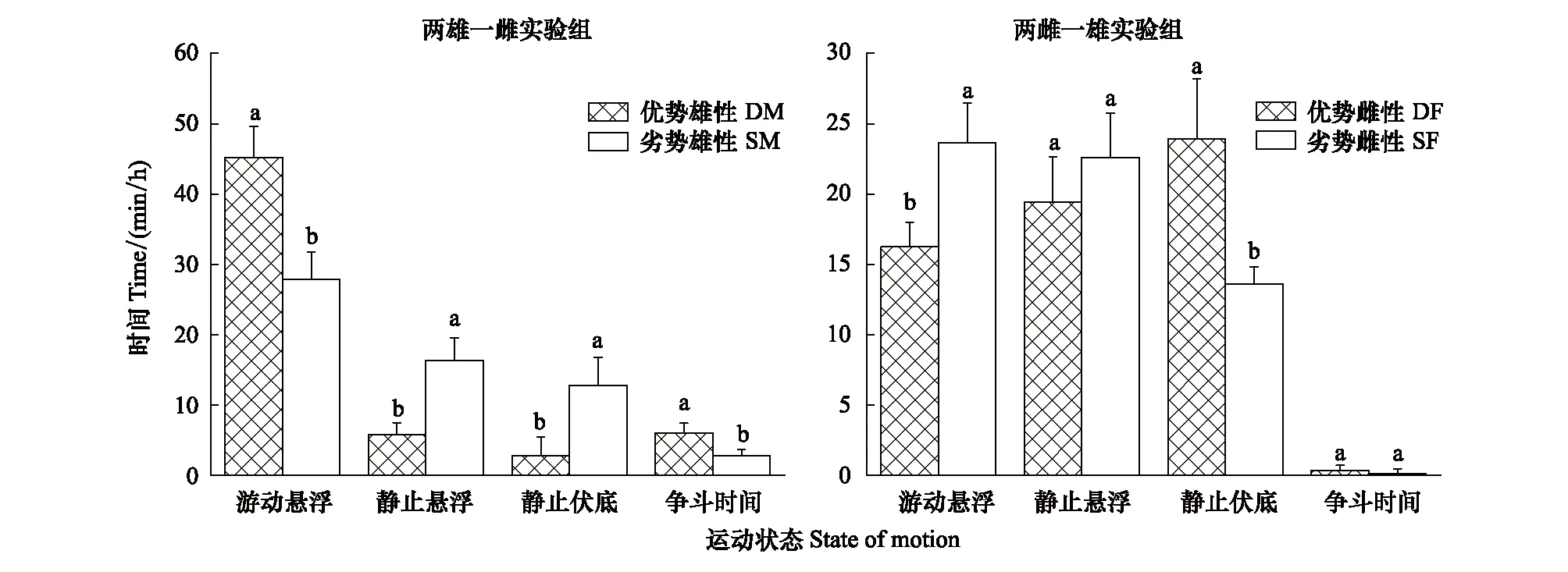
图3 不同优势等级个体四种不同运动状态的时间分配
2.3 社群等级对金乌贼代谢酶活力的影响
通过比较分析肌肉组织和性腺组织有氧代谢通路中的苹果酸脱氢酶(MDH)、柠檬酸合酶(CS)活性发现,(2♀1♂)实验组优势雌性个体肌肉中的MDH、CS活性分别为(170.982±11.157)U/gprot、(19.747±1.562)U/gprot,显著低于劣势个体MDH、CS活性(281.402±10.977)U/gprot、(24.125±1.164)U/gprot(P<0.05)。而性腺中MDH、CS活性显著高于劣势个体(P<0.05)。在(2♂1♀)实验组中,优势雄性个体肌肉的MDH、CS活性分别为(309.69±16.147)U/gprot、(34.927±1.479)U/gprot,显著高于劣势个体(212.131±6.936)U/gprot、(26.696±1.21)U/gprot(P<0.05),而在性腺中优势和劣势个体MDH活性无显著差异(P>0.05)(图4)。
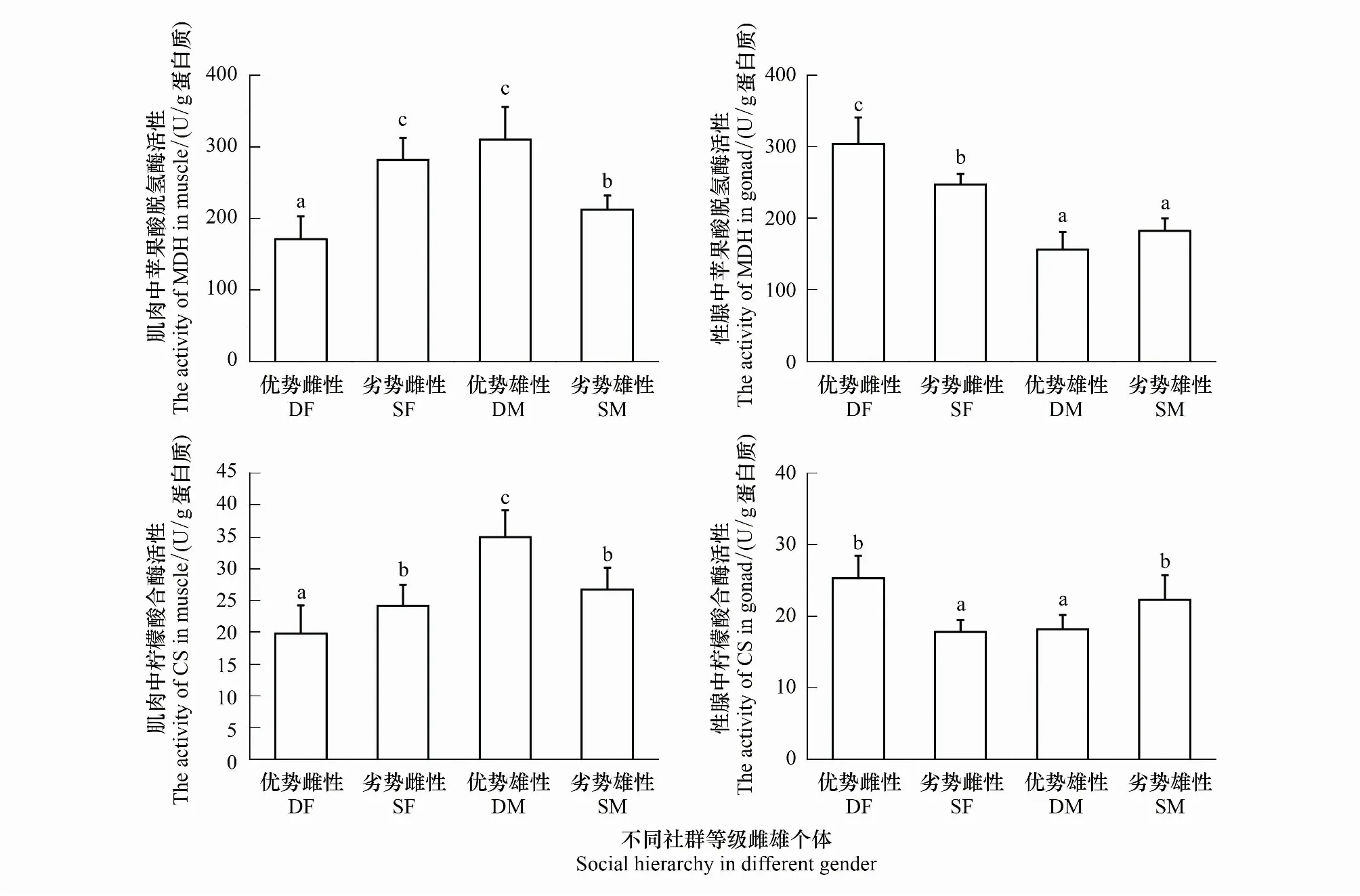
图4 不同优势等级雌雄个体肌肉和性腺组织中有氧呼吸代谢酶活性(n=8)
如表2所示,优势雄性个体肌肉中己糖激酶(HK)、丙酮酸激酶(PK)、乳酸脱氢酶(LDH)3种无氧代谢酶活性分别为(12.453±0.858)U/gprot、(18.269±0.583)U/gprot、(63.220±2.509)U/gprot,显著高于劣势雄性个体(P<0.05)。优势雄性个体性腺中的无氧呼吸代谢酶活性与劣势雄性个体之间差异不显著(P>0.05)。优势雌性个体和劣势雌性个体肌肉和性腺之间无氧代谢酶活性差异均不显著(P>0.05)。
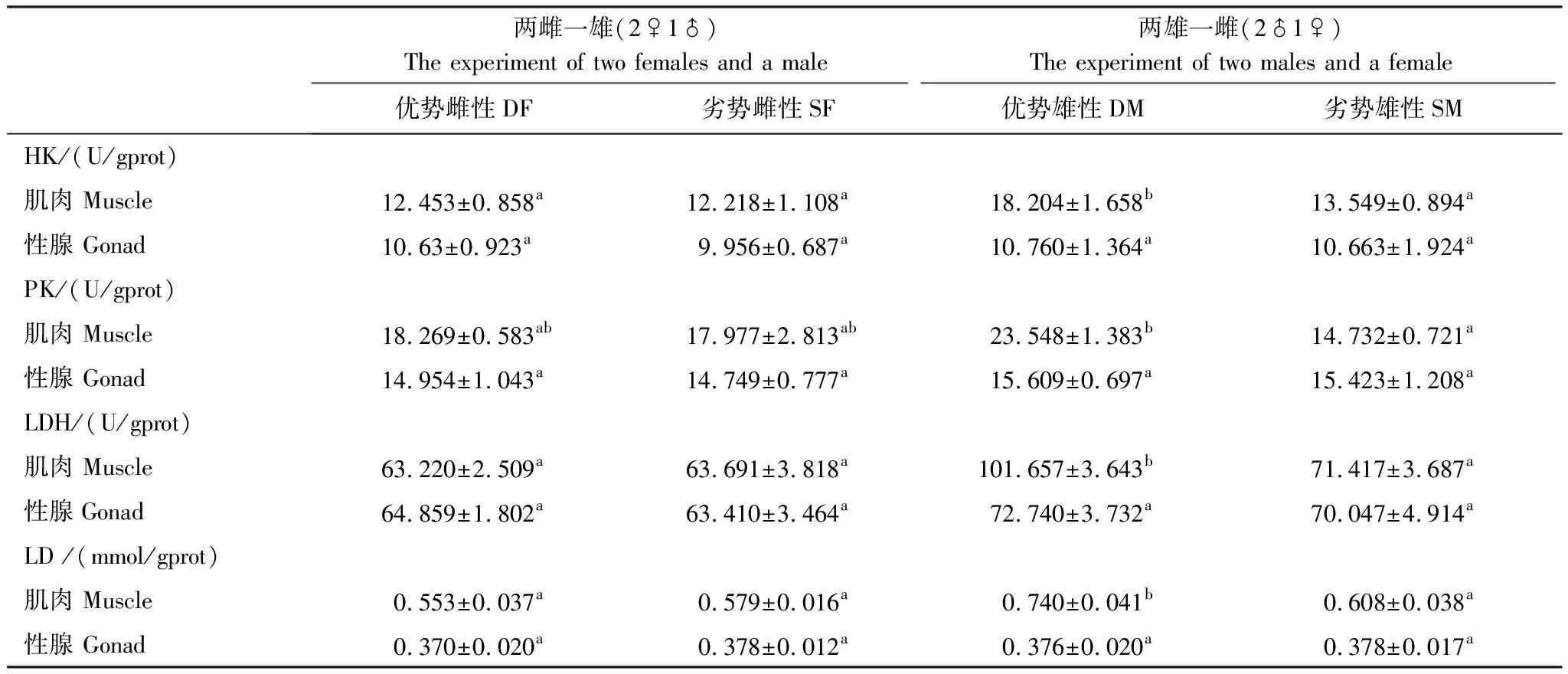
表2 不同优势等级雌雄个体肌肉和性腺组织中无氧代谢酶活性及乳酸含量
2.4 繁殖期金乌贼行为表型与代谢酶活力的相关性
比较分析发现社群等级对于金乌贼行为表型和能量代谢具有显著影响,不同优势等级雌雄亲体的游动悬浮时间分别与苹果酸脱氢酶(MDH)、柠檬酸合酶(CS)两种有氧代谢酶活性之间呈线性相关:y=5.093x+99.519(R2=0.367,F=17.419,P<0.001)、y=0.510x+11.940(R2=0.706,F=2.165,P<0.001)(图5)。
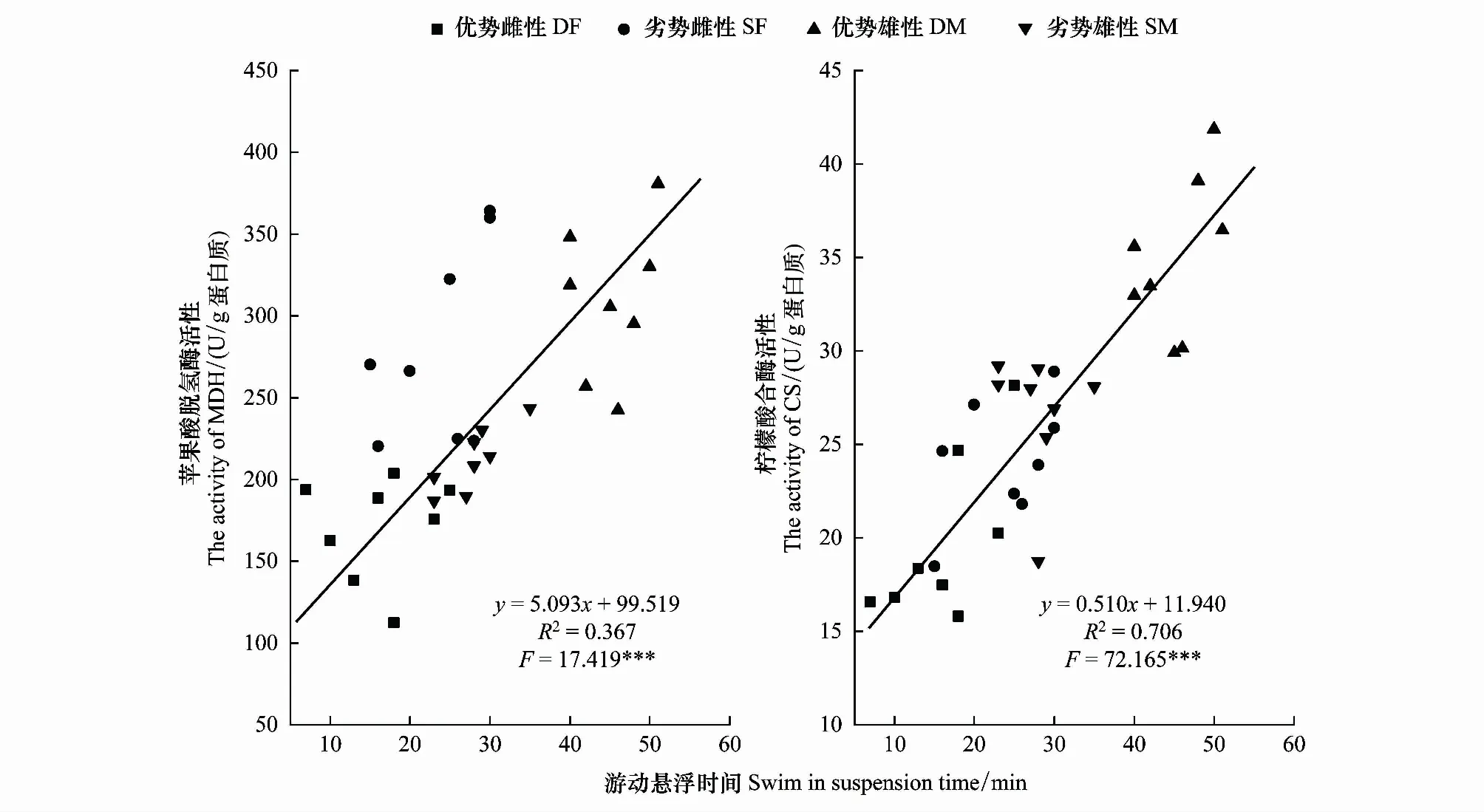
图5 金乌贼游动悬浮时间与肌肉中有氧代谢酶活性的关系
同理分析可得,个体的争斗时间与己糖激酶(HK)、丙酮酸激酶(PK)、乳酸脱氢酶(LDH)活性之间的关系呈显著正相关,拟合的曲线方程分别为y=1.072x+1.559(R2=0.578,F=41.141,P<0.001)、y=0.770x+1.946(R2=0.137,F=4.762,P<0.05)、y=4.671x+63.902(R2=0.541,F=37.737,P<0.05)(图6)。进一步分析争斗时间与乳酸(LD)含量之间的关系发现,随争斗时间的增加,金乌贼腕部肌肉乳酸含量显著增加(P<0.001)(图6)。这表明金乌贼在发生剧烈争斗行为时,运动时间短,主要通过无氧代谢供应能量。

图6 金乌贼争斗时间与肌肉中无氧代谢酶活性和乳酸含量的关系
3 结论和讨论
3.1 行为交互作用与社群等级之间的关系
社群等级的形成对个体行为产生很大影响,社群等级高的个体具有较强的攻击性,能够占据主要食物源和领地[20]。在社群等级胁迫下,劣势个体处于长期压力状态,表现为运动能力下降,并且影响其生长、代谢、免疫等功能[21]。有研究表明,劣势鱼在遭到优势鱼的“视力”威胁后,会出现躲避、减小运动范围等行为[22]。本研究中,雄性个体之间具有较明显的争斗行为,社群等级现象显著。优势个体主动用腕迅速靠近劣势个体并击打劣势个体的胴部,同时伴随劣势个体的躲避、喷水、喷墨等防御行为。优势雄性个体长时间悬浮伴游在雌性个体周边,劣势雄性尝试靠近后被优势个体阻拦,静止悬浮在水体中或者底部。而雌性个体之间未发现明显的争斗行为,但优势雌性个体与雄性个体交配次数多、放卵次数多,多次放卵后会停留在水层底部,呈现静止状态。
3.2 社群等级与能量代谢之间的关系
Li等[23]研究发现,水生动物低强度运动时主要通过有氧代谢获取能量,随运动强度增加,能量代谢方式由有氧代谢转为厌氧糖酵解,原因主要是与氧气供应有关[10]。已有研究发现己糖激酶(HK)、丙酮酸激酶(PK)、乳酸脱氢酶(LDH)对糖酵解速率控制发挥关键作用,产生乳酸(LD)等代谢产物[24-26]。MDH能催化苹果酸与草酰乙酸之间的可逆转换,CS在三羧酸循环中可催化草酰乙酸和乙酰辅酶A之间的缩合反应,二者都是三羧酸循环的关键酶[27-28]。低强度运动氧气供应充足,因此动物通过有氧代谢获取能量[11]。本研究中,由于优势雄性个体长时间处于游动悬浮状态,主要依靠腕部肌肉和腹部肌肉轻微收缩运动,其肌肉有氧代谢酶(MDH、CS)活性显著高于劣势个体,进一步证实了金乌贼在游动悬浮时主要进行有氧代谢活动。
社群等级形成过程中,个体间通过争夺配偶以及领域宣誓主权,易产生剧烈争斗行为[29]。而动物在产生急速运动或剧烈争斗时,短时间内需要提供大量能量,主要由无氧呼吸提供[30]。在凡纳滨对虾(Litopenaeusvannamei)、尼罗罗非鱼(Oreochromisniloticus)中随着游泳速度的增大,无氧呼吸酶活力显著增加[9,26]。本研究发现,优势雄性个体的无氧代谢酶活性(PK、HK、LDH)显著高于劣势雄性个体,分析认为优势雄性个体主动发起急速攻击次数多,时间短,腕部以及胴部肌肉剧烈收缩和拉伸,无氧代谢活动加剧。在岸蟹(Carcinusmaenas)、寄居蟹(Birguslatro)中发现由于争斗导致体内乳酸含量的增加[31-32],本研究中优势雄性个体肌肉中乳酸含量也显著高于劣势雄性个体。而雌性个体之间争斗行为少,无激烈竞争交互作用,无氧代谢酶活性差异不显著。无氧代谢酶活性及乳酸含量与雄性争斗时间呈显著正相关,也进一步证实了金乌贼在争斗过程中主要以无氧代谢供能。
3.3 繁殖期雌雄个体的能量分配
研究表明,繁殖是一个高能量需求过程,性腺的生长发育以及卵子的形成等都需要能量供应[33-34]。由于动物摄入的食物资源有限,即能量摄入有限,用于繁殖能量的增加必然会导致用于其他生命活动的能量支出减少[33]。在“生物行为节能假说”中也提到繁殖与运动行为间存在能量竞争关系;动物通过减少自身运动时间,将能量优先用于繁殖过程[35-36]。在自然条件下,为了保证个体适应性,繁殖期的生物常通过降低生长率、减少运动能量消耗等来保证生殖成功,这一过程也称为“生殖代价”[33,37],合理的能量投资能够使生殖价最大化。Ru等[38-39]研究发现仿刺参(Apostichopusjaponicus)在繁殖过程中通过减少运动时间和运动距离以及降低生长率、摄食率从而减小能量分配以维持繁殖过程中的高能量需求。而在鱼类中同样有此现象发生,繁殖期的短脚床杜父鱼(Myoxocephalusscorpius)和食蚊鱼(Gambusiaaffinis)的最大游泳速度出现显著下降[40-41]。研究表明,头足类在繁殖期减少甚至停止摄食,Quetglas等研究发现尖盘爱尔斗蛸(Eledonecirrhosa)的性腺重量在非繁殖期占体重的1%,而在繁殖期达到了体重的20%—25%[42]。在减小或者停止外源性能量摄入的同时,机体储能减少,而性腺重量的增加,加大了机体的物理负担,导致运动能力下降。本研究中优势雌性个体产卵后会长时间静止伏底或静止悬浮于水体中,自发活动明显降低,用于运动的能量减少,肌肉的有氧代谢酶活性显著低于劣势个体,但性腺有氧代谢酶活性显著高于劣势个体,推测是优势雌性个体大部分能量用于卵子的生成和发育所致。综上,行为的可塑性是繁殖期能量分配的一种有效策略,雌性金乌贼通过减少运动能耗来保障繁殖的高能量需求。
繁殖期雌雄个体具有不同的能量分配方式,尼罗罗非鱼(Oreochromisniloticus)雄鱼比雌鱼生长速度快,在繁殖期间雌雄生长差异更加显著[43],雌性个体将更多的能量分配到生殖上,用于生长的能量减少。繁殖期中华绒螯蟹(EriocheirSinensis)雌蟹将44%能量分配至性腺和肝胰腺中,高于雄蟹的23%[44]。本研究中雄性肌肉中的MDH和CS两种有氧代谢酶活分别是雌性个体的1.2倍和1.4倍,繁殖期雄性个体运动能耗高于雌性个体。而雌性个体性腺中的MDH和CS酶活分别是雄性个体的1.6倍和1.1倍,雌性个体将更多能量用于配子形成和性腺发育。而雄性个体间通过相互争斗,获取更多交配机会,将精荚粘附在雌性个体口膜处,并伴游护卫雌性个体,以保证自己的繁殖成功率,因而将更多能量用于争斗和运动中。
4 总结
金乌贼人工繁育过程中,社群等级的形成使得不同等级雌雄个体间的行为产生差异,优势雄性个体以交配投资为主,因而具有更强的攻击性、争斗和运动能力。雌性个体以产出更多的卵子作为主要生殖投资,具有较强的领域性,优势雌性个体用于产卵繁殖的能量较多。行为活动的差异也导致了繁殖亲体有氧和无氧代谢酶活性的差异。有关金乌贼社群等级的差异是否会影响亲体的繁殖效率以及子代的质量等仍需要进一步研究。金乌贼为一年生头足类,繁殖期经多次交配多次产卵,繁殖后快速死亡。在其短暂的生命周期中,生殖代价与衰老和死亡之间错综复杂的关系仍有待考证。
