短时高温暴露对菊方翅网蝽成虫存活和生殖特性的影响
2020-09-14潘颖佳姚露晨李若南崔旭红
蔡 冲,潘颖佳,姚露晨,李若南,崔旭红
(中国计量大学生命科学学院/浙江省生物计量及检验检疫技术重点实验室,杭州 310018)
菊方翅网蝽CorythuchamarmorataUhler原产于北美,后传入欧洲,21世纪初先后在日本和韩国被发现报道,并迅速扩散。2012年在中国上海首次发现了菊方翅网蝽(党凯等,2012),随后在浙江、台湾、湖北、江苏、江西、河南等地发现了该虫的踪迹(虞国跃,2014;董立坤等,2015;卢和平等,2018;)。菊方翅网蝽喜食菊科植物,除了危害加拿大一枝黄花、野塘蒿、小蓬草、紫苑等野生杂草外,还可以对菊科的菊花、向日葵、菊芋、紫背天葵以及旋花科的甘薯等多种园艺植物、经济作物和蔬菜造成危害或威胁(Kato and Ohbayashi, 2009; Hoshino, 2011;党凯等,2012;Yoonetal., 2013; Kim and Kil, 2014)。昆虫是变温动物,对温度变化,尤其是极端温度的出现反应敏感。目前气候变化的一个显著特点就是气温上升、极端气温出现频繁,这难免会对昆虫种群的发生产生影响。根据报道,在韩国部分地区7月上旬至8月下旬是菊方翅网蝽的发生盛期;在日本大阪地区菊方翅网蝽在6月中旬至8月中旬发生量最大(Mamoruetal., 2006; Kim and Kil., 2014)。沈佳思等(2016)通过对菊方翅网蝽生物学特性的研究推测8月-10月是该网蝽在我国华中以及华东地区的发生盛期,这与我们在杭州地区的田间观察结果基本一致。以上地区的7月-8月份正是盛夏时节,部分地区日最高温度可持续多天达到35℃,甚至超过40℃(张立生等,2014),在这种环境条件下,活动性不强的菊方翅网蝽很容易暴露在高温环境下受到胁迫。根据报道,短时的高温暴露可以对昆虫造成直接或间接的伤害,从而影响昆虫种群的发展与分布。例如短时高温暴露能造成昆虫个体死亡、发育异常缺陷、生殖力降低、行为改变等(赵鑫等,2009;丛林等,2010;Piyaphongkuletal., 2012; Ma and Ma, 2012; Zhangetal., 2013)。为了明确菊方翅网蝽对高温的耐受性,了解菊方翅网蝽在我国的入侵潜能,本研究通过室内短时高温暴露实验,研究了菊方翅网蝽成虫经过高温暴露后的存活、繁殖、F1代存活能力、性比等特性的变化,希望为菊方翅网蝽种群的发生预测及治理提供理论依据。
1 材料与方法
1.1 供试昆虫与寄主植物
供试昆虫:菊方翅网蝽种群建立于2016年,寄主植物为加拿大一枝黄花。在进行高温暴露实验之前,从建立的种群中采集菊方翅网蝽成虫接入养虫笼(60 cm×60 cm×60 cm)中进行继代饲养,随后获得的网蝽用于实验。饲养条件为温度26±1℃,相对湿度75%±10%,光照14 h ∶10 h (L ∶D)。
寄主植物:参照沈佳思等(2016)方法进行。剪取加拿大一枝黄花长约20 cm的枝条,保留干净的叶片6~8张,每两张叶片间隔>2 cm,叶片长约5 ~8 cm。植株茎干用海绵包裹保湿并固定于装有水的塑料杯中。将其作为清洁苗供实验使用。
1.2 短时高温暴露对菊方翅网蝽成虫存活率和寿命的影响
采集菊方翅网蝽成虫,辨别雌雄后分别装入玻璃管(Φ=2.5 cm,h=9 cm)中进行短时高温暴露实验。玻璃管中有黄花叶片供其取食,管口塞有少量脱脂棉防止成虫逃逸,每管雌虫或雄虫30头。高温暴露实验在水浴循环仪(型号:9112AA2Y,Poly Science公司)中进行,共设置了6个靶标温度,分别为35℃、37℃、39℃、41℃、43℃和45±0.02℃。在每个温度梯度下,处理时间为2 h。处理结束后,将成虫放置在饲养条件下2 h让其恢复苏醒,随后调查其存活情况。以毛笔轻触虫体,若附肢不动则认为死亡。存活的成虫放在养虫盒后饲养于人工气候箱(RXZ-5008型,宁波江南科技有限公司),每隔24 h调查1次其存活情况,并及时更换新鲜的叶片,直至成虫死亡。以上每个处理重复5次。以未经过高温处理直接放置在26℃下同等时间的处理为对照。最后统计雌雄虫存活率及其寿命。
1.3 短时高温暴露对菊方翅网蝽的生殖特性的影响
高温暴露处理方法:采集羽化48 h内的菊方翅网蝽成虫,辨别雌雄后按照1.2中的方法进行短时高温暴露,随后在饲养温度下恢复2 h并取存活的成虫进行后继实验。
短时高温暴露后成虫的生殖特性:将完成高温暴露的成虫放入培养皿中(1对成虫/皿,♀ ∶♂=1 ∶1),培养皿中放有加拿大一枝黄花叶片供其取食,然后将其置于人工气候箱中饲养,饲养条件同1.1。根据前期的预实验结果情况,5 d以后开始调查雌虫的产卵情况,每隔24 h调查1次直至雌虫死亡。在实验过程中若雄虫先死亡,则补充雄虫。实验过程中及时更换新鲜的加拿大一枝黄花叶片。以上每个处理不少于20次重复,以放在26℃下饲养的处理为对照。最后统计菊芳翅网蝽的产卵前期、产卵期、产卵量等。
短时高温暴露后F1代存活能力和性比:采集上述实验雌虫所产的F1代卵,将产卵叶片放入培养皿中,标记产卵日期,每24 h调查1次卵孵化的情况,待卵孵化成若虫后,将若虫继续饲养,直至若虫发育成成虫。最后统计卵孵化率、卵至成虫的存活率、雌成虫比例等。每处理重复20次。以在26℃下饲养成虫所产的F1代为对照。饲养条件同1.1,F1代存活率=羽化成虫数量/卵量。
1.4 数据处理与分析
用统计软件SPSS(SPSS 24.0,SPSS Inc., USA)对数据进行处理分析。用t-Test检验在相同高温暴露条件下雌雄成虫存活率是否存在显著性差异;用单因素方差(One-way ANOVA)检验不同高温暴露条件下菊方翅网蝽的存活率、寿命、产卵前期、产卵量和后代存活率等是否存在显著性差异。显著性检验水平均为P<0.05。
2 结果与分析
2.1 短时高温暴露对菊方翅网蝽成虫存活和寿命的影响
菊方翅网蝽雌虫在35~39℃下将暴露2 h,其存活率均在100%;在41℃下暴露2 h,其存活率为92.0%;在43℃下暴露2 h后,雌成虫的存活率迅速下降至40%,与对照相比存在显著差异。雄成虫在35~37℃下暴露2h,其存活率与对照相比没有显著差异。39~41℃下暴露2 h,其存活率下降至94.7%和88.7%,显著低于对照。在43℃下暴露2 h,雄虫存活率迅速下降至34.7%,与对照及其它暴露温度存在显著差异;在45℃下暴露 2 h,没有成虫能够存活(表1)。因此,41℃及以上高温对菊方翅网蝽成虫的存活率有显著影响(P<0.05)。在35~43℃温度范围内,同一暴露温度下,通过t检验结果,雌虫比雄虫的存活率高,但差异均不显著(P>0.05)。35~39℃的短时暴露对雌雄成虫的寿命没有显著影响,当暴露温度上升至41℃时,雌雄虫的寿命分别为54.1 d和51.3 d,与对照存在显著差异。43℃下暴露2 h,对雌雄虫的寿命有显著影响,没有成虫存活的时间超过1 d。

表1 菊方翅网蝽成虫高温暴露2 h后的存活率和寿命Table 1 Average survival rate and longevity of Corythucha marmorata adults after brief exposure to high temperature for 2 hours
2.2 短时高温暴露对菊方翅网蝽成虫产卵特性的影响
在35~41℃范围内暴露2 h,随着暴露温度的升高,菊方翅网蝽的产卵前期逐渐延长(表2)。在对照(26℃)条件下,菊方翅网蝽的产卵前期为6.1 d;在35~41℃下暴露2 h,其产卵前期由8.3 d上升到9.4 d,与对照相比存在显著差异(P<0.05)。在43℃下暴露2 h,菊方翅网蝽由于存活不超过1 d,没有产卵行为(表2)。在35~39℃范围内暴露2 h,菊方翅网蝽的产卵期由51.7 d下降至49.3 d,与对照相比没有显著差异。在41℃下暴露2 h,菊方翅网蝽的产卵期下降至44.5 d,显著低于对照。在26℃时,平均每头雌虫总产卵量为96.3粒,在35~39℃范围下暴露2 h,产卵量与对照比较,没有显著变化。当暴露温度上升至41℃时,产卵量显著出现下降。
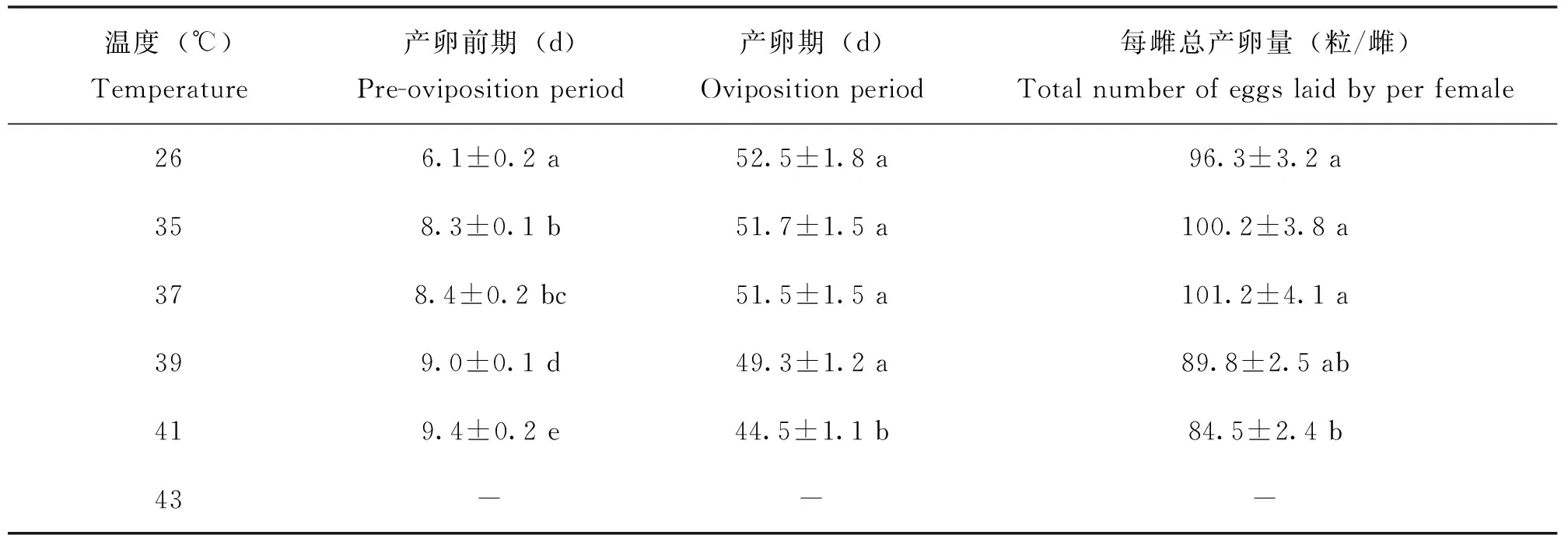
表2 菊方翅网蝽雌虫成虫高温暴露2 h后产卵前期及每雌总产卵量Table 2 The pre-oviposition period, oviposition period and the total number of Corythucha marmorata eggs laid by per female adult after brief exposure to high temperature for 2 hours
2.3 短时高温暴露对菊方翅网蝽后代生存能力的影响
菊方翅网蝽成虫在35~41℃高温暴露下2 h,其F1代卵的孵化率由70.2%逐渐下降到40.7%,各温度之间孵化率存在显著差异,并显著低于对照(P<0.05);F1代总存活率也表现出相同的趋势(P<0.05),对照的F1代若虫总存活率达到70.9%,而经过各高温暴露后F1代的总存活率由64.5%下降至34.8%(表3)。菊方翅网蝽成虫在35~41℃高温暴露下2 h后,其F1代成虫的雌成虫比例在49.5%~50.5%之间,与对照相比无显著差异(P>0.05)。
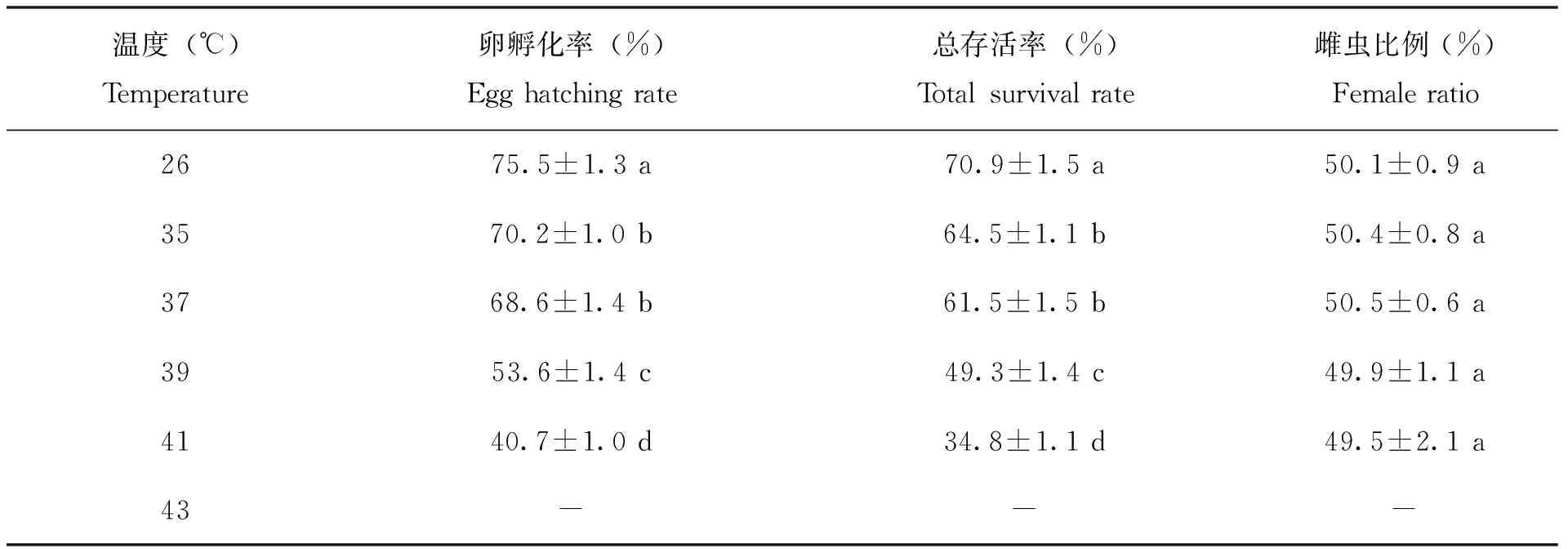
表3 菊方翅网蝽雌虫成虫高温暴露2 h后F1代卵存活率、总存活率及雌虫比例(%)Table 3 The egg hatching rate, total survival rate and female ratio of F1 progeny of Corythucha marmorata after brief exposure to high temperature for 2 hours
3 结论与讨论
在近年来全球变暖的趋势下,高温热浪天气发生频次的上升已经影响到一些动物的种群动态(Pacificietal., 2017)。昆虫是变温动物,对温度变化敏感,环境温度是决定昆虫存活、分布、行为、种群发展等的关键因素。根据报道,不同动物对气候变化的响应不同,这些影响可能最终会在种群动态上得以反映。例如,高温胁迫能提高跳虫、卤虫Artemia、果蝇Drosophilamelanogaster后代对高温的适应性或抗性(Gilchrist and Huey, 2001; Zizzari and Ellers, 2014; Norouzitallabetal., 2014)。但也有大量的报道表明高温胁迫不仅可以使昆虫迅速死亡,还可以造成产卵量下降、性比失调、雄虫对雌虫的吸引力降低、后代的存活力下降等(Huangetal., 2007; Cuietal., 2008;丛林等,2010;Zhouetal., 2011; Zizzari and Ellers, 2011; Bodlahetal., 2017; Greenetal., 2019),从而使得后代种群在高温条件下处于不断衰退的趋势之中。入侵我国的B型烟粉虱和温室白粉虱在中国北方混合发生,其种群消长有明显的季节性差异(罗晨和张芝利,2000;Ramosetal., 2002),这与两者对高温的耐受性差异有关,高温对温室白粉虱存活、生殖等方面的不利影响均要明显大于B型烟粉虱(Cuietal., 2008)。因此,在夏季高温季节B型烟粉虱处于发生盛期,而温室白粉虱则在气候较为凉爽的秋季占据优势。本研究结果表明,菊方翅网蝽成虫对高温具有一定的耐受性,41℃及其以上高温暴露才会对菊方翅网蝽雌雄虫存活率、产卵前期、产卵期和产卵量等特性造成显著影响。在菊方翅网蝽发生地区,超过40℃的气温天气较少,因此,高温气候对该网蝽种群直接的影响比较少。但高温可以通过降低昆虫的产卵量、后代存活力和性比等来间接影响种群的发展。例如,37~45℃范围内短时暴露1 h对 B型烟粉虱成虫的产卵量没有显著影响,但后代的存活率随着暴露温度的升高迅速下降;43℃以上的高温暴露1 h还会导致B型烟粉虱Bemisiatabaci成虫后代雄虫的比例的上升,从而影响到种群的结构和发展(Cuietal., 2008)。在本研究中发现,在35~39℃范围内菊方翅网蝽成虫受到高温暴露后,其存活、产卵量等虽然不受显著影响,但其F1代的孵化率和存活率下降明显,与对照存在显著差异。在自然界中夏季35~39℃高温比较常见,正午温度达到最高时也可以持续超过2 h,因此,遭受了高温暴露菊方翅网蝽后代的存活力难免会受到影响。但由于菊方翅网蝽生长发育周期和产卵期均较长,高温对其的影响会表现出明显的滞后性。因此在田间表现出7月-9月大量发生,10月中下旬后出现种群数量的显著下降的情况。
在自然条件下,气温变化比较复杂,若气温缓和升高,昆虫一般会经历热锻炼(rapid heat hardening),这可能会影响到昆虫对高温胁迫的耐受性。有研究表明,一些昆虫在经历暂时性的或季节性的温度升高时,其存活、繁殖力和性比等会相应发生变化,进而影响其种群发展。例如,悬铃木方翅网蝽Corythuochaciliata可以通过快速热锻炼迅速适应38℃左右的高温(Juetal., 2011)。悬铃木方翅网蝽在不同升温模式下的种群增值研究表明,其种群内禀增长率在夏季均温升高和极端高温频发的情况下会显著上升,种群的世代周期则显著缩短。高温对该网蝽种群爆发具有刺激作用(Juetal., 2015)。若温度变化迅速剧烈,昆虫所面临的挑战可能更大,虽然仅经历1次的短时高温胁迫,也有可能对其种群发展造成负面影响。例如,小菜蛾Plutellaxylostella在经历40℃高温胁迫数小时后,其存活、交配、寿命等虽然没有影响,但其前2 d的产卵量及卵的孵化率显著下降。推测后代卵孵化率的降低可能与精子发生有关(Zhangetal., 2013)。高温还可能会通过干扰神经激素延迟卵黄的形成和卵的成熟对生殖进行调节(Rauschenbachetal., 1995; Rauschenbachetal., 1996; Gruntenkoetal., 2003);高温胁迫后生物体产生的热激蛋白Hsp70也有可能对卵的孵化产生有害影响(Silbermann and Tatar, 2000)。本研究结果表明,高温胁迫对菊方翅网蝽后代生存的影响较大,在35~39℃范围内菊方翅网蝽成虫受到高温暴露后,其F1代的孵化率和存活率下降明显,为了更好的明确菊芳翅网蝽入侵潜能,后期还需要对不同虫态、性别的菊方翅网蝽在不同升温模式下的存活和生殖特性以及耐热机制开展研究。
