3种生境蚂蚁群落组成和觅食动态调查
2020-09-14许益镌
聂 蕾,熊 煜,许益镌
(华南农业大学昆虫学系,广州 510642)
蚂蚁是地球上生物量最大的类群之一,发挥着重要的生态服务功能。迄今为止,已有超过12 000个物种被描述,并且有许多蚂蚁种类还没有被鉴定(Lachetal., 2009)。由于蚂蚁的多样性以及在所有能级水平的生态重要性都非常高、很容易采集以及对生态变化的敏感性,已经成为指示生物多样性和环境变化的重要物种(Andersen, 1990; Alonso, 2000; Kaspari and Majer, 2001)。随着人们对生态环境重视程度的提升,以蚂蚁作为指示生物对生态恢复状况进行评价受到了许多国家的重视(Osbornetal., 1999; Wattetal., 2002)。
生态位分化被认为是物种共存的主要机制,不同蚂蚁种类占据不同的营养生态位,但具有类似生态位的物种如何避免竞争排斥仍然存在争议(Houadriaetal., 2015)。调查生境中不同蚂蚁觅食动态规律可为深入理解这一问题提供科学依据。诱饵法是一种操作简便、经济合理、应用广泛的蚂蚁群落多样性调查以及蚂蚁觅食行为的研究手段(李巧等,2009; Kaspari and Yanoviak, 2001)。为研究相同生境中不同种类蚂蚁觅食规律的差异,及进一步比较不同蚂蚁营养生态位差异提供依据,本研究分别用20%蔗糖溶液和鸡肉火腿肠以诱饵法诱集湖边、树林、荔枝园3种生境中的蚂蚁,以此确定不同生境中的优势蚂蚁种群及其觅食行为随时间的动态变化。
1 材料与方法
1.1 调查地点
本研究在广东省广州市华南农业大学校园内进行调查,调查取样在2018年3月底至4月底完成,此时广州平均月雨量达200 mm,平均湿度达84%。在每日中午12点至下午5点间蚂蚁觅食活动较活跃时进行诱集,平均气温为25~30℃。所调查的3个生境分别为湖边、树林和果园。湖边生境选取校园内沼泽湖畔(图1左),该生境中植被种类丰富,主要有宫粉羊蹄甲BauhiniavariegataLinn.、水鬼蕉Hymenocallislittoralis(Jacq.) Salisb.、香蕉MusananaLour.、蒲桃Syzygiumjambos(Linn.) Alston、大叶相思AcaciaauriculiformisA. Cunn. ex Benth以及狗牙根Cynodondactylon(Linn.) Pers.、含羞草MimosapudicaLinn.、香菇草HydrocotylevulgarisL.、白花鬼针草BidenspilosaLinn. var.radiataSch.-Bip.、蟛蜞菊Wedeliachinensis(Osbeck.) Merr.等杂草。树林生境选取校园内树林(图1中),植被以人面子DracontomelonduperreanumPierre、假槟榔Archontophoenixalexandrae(F. Muell.) H. Wendl. et Drude、桉树EucalyptusrobustaSmith、短穗鱼尾葵CaryotamitisLour.为主。果园生境选取园艺学院荔枝园(图1右),该生境中除主要种植荔枝LitchichinensisSonn.外,密布着白花鬼针草BidenspilosaLinn. var.radiataSch.-Bip.等常见杂草。每种生境选取3个面积均约为1 300 m2的小区,小区间隔约为50 m。

图1 湖边(左)、树林(中)和荔枝园(右)生境现场照片Fig.1 Photos of lakeside (left), forest (middle) and litchi orchard (right) habitats
1.2 试验材料
蔗糖(太古白砂糖,太古糖业(中国)有限公司,广州)溶液用水配制为20%浓度后,用移液枪加入30~60 mL透明塑料瓶中,用纸折叠后的一节卷纸纸巾塞入瓶中蘸吸糖液以防止蚂蚁觅食时被淹死。火腿肠(双汇精制鸡肉肠,河南双汇集团有限公司,漯河市)切为厚度为2~3 mm的薄片,每诱瓶放入一片备用。
1.3 调查方法
每种诱饵在每个小区放置50个诱瓶进行取样,每瓶间隔4~6 m,重复3个小区。用蔗糖溶液引诱,设置30 min、60 min、90 min、120 min、150 min共5个调查时间点,每个时间点回收10个诱瓶。用火腿肠引诱,设置15 min、30 min、45 min、60 min、75 min共5个调查时间点,每个时间点回收10个诱瓶。
1.4 标本鉴定
诱集到的蚂蚁用90%酒精浸泡,整理后进行种类鉴定。
1.5 数据处理
采用卡方检验比较不同生境和不同饵剂诱集的蚂蚁种类数和个体数量。用单因素方差分析比较3种生境蚂蚁在5个不同观察时间分别对蔗糖溶液和火腿肠的觅食强度以及每个观察时间3种不同生境蚂蚁对蔗糖溶液的觅食强度,采用DMRT法进行多重比较。采用单因素方差分析比较不同生境不同诱饵优势种在5个不同观察时间的觅食强度以及每个观察时间下不同优势种的觅食强度差异,采用DMRT法进行多重比较。
2 结果与分析
2.1 3种生境蚂蚁种类多样性和结构分析
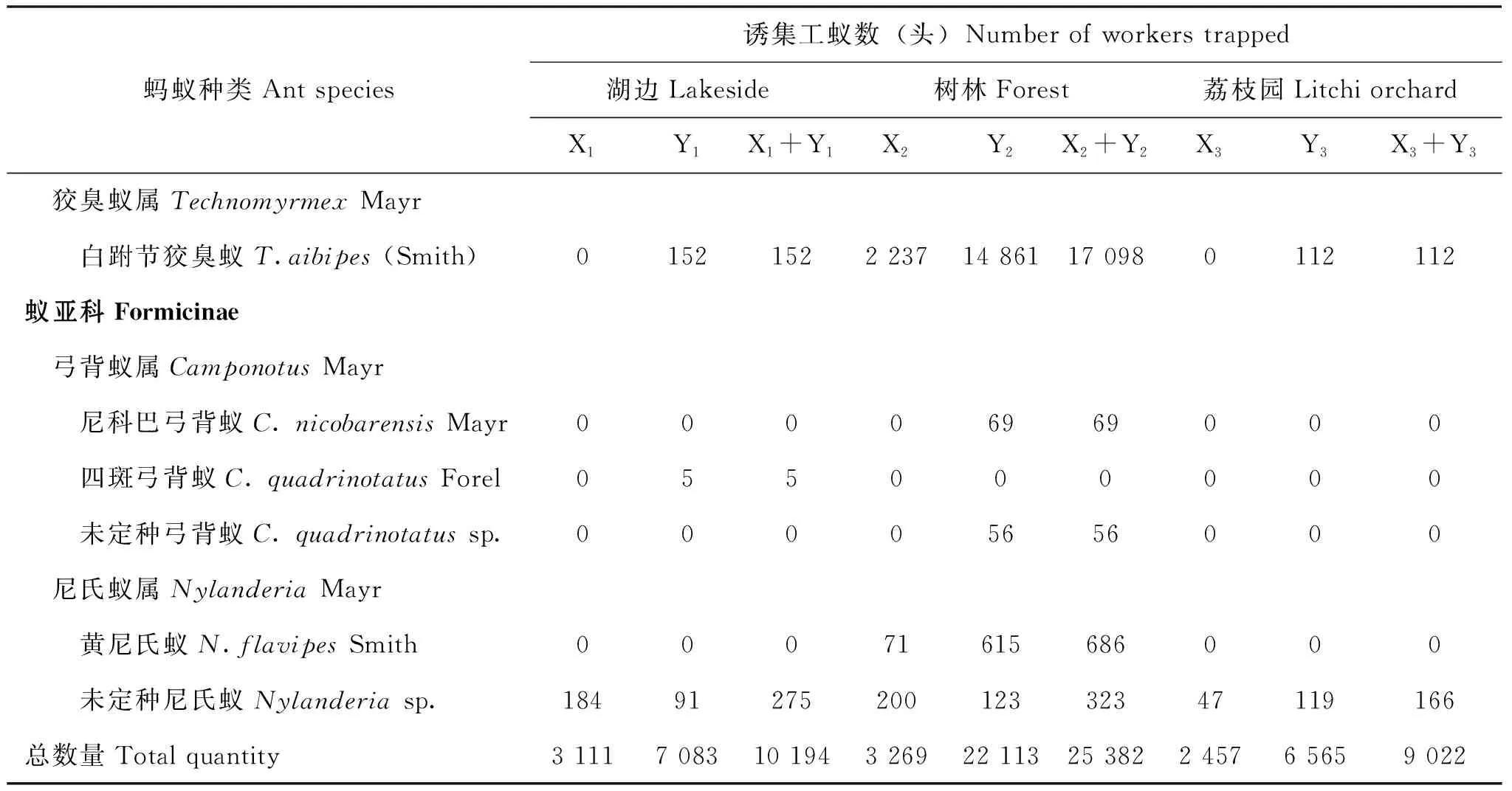
综合蔗糖溶液和火腿肠诱集的结果,在湖边生境共诱集10 194头蚂蚁,分属于3亚科、8属、8种(表1)。其中,切叶蚁亚科4属4种,臭蚁亚科2属2种,蚁亚科2属2种。在树林生境共诱集25 382头蚂蚁,分属于4亚科、6属、9种。其中,猛蚁亚科1属1种,切叶蚁亚科2属3种,臭蚁亚科1属1种,蚁亚科2属4种。在荔枝园生境共诱集9 022头蚂蚁,分属于4亚科、7属、7种。在诱到的蚂蚁中,小家蚁属、弓背蚁属、尼氏蚁属未能鉴定到种的蚂蚁均各算作1种。

表1 3种生境蚂蚁类群物种组成Table 1 Species composition of ants in three habitats
续表1 Continued table 1
虽然不同生境诱集到的蚂蚁种类数无显著差异(χ2=0.25, df=2,P=0.882),但诱集到的蚂蚁数量则表现生境差异(χ2=11204.5, df=2,P<0.001)。其中宽结大头蚁、尼科巴弓背蚁、未定种弓背蚁、黄尼氏蚁仅见于树林。未定种小家蚁、四斑弓背蚁仅见于湖边。且树林不见黑褐举腹蚁和黑头酸臭蚁。湖边不见横纹齿猛蚁。据调查结果显示,采用火腿肠诱集到的蚂蚁种类和数量都比蔗糖溶液诱到的多(P<0.05)。
3种生境中占有优势地位的蚂蚁种类存在差异(表2)。在湖边生境有3种蚂蚁占据优势地位,分别是小大头蚁(44.21%)、未定种小家蚁(26.27%)和黑头酸臭蚁(24.72%)。在树林生境也有3种蚂蚁占据优势地位,分别是白跗节狡臭蚁(67.36%)、小大头蚁(20.98%)和宽结大头蚁(6.93%)。在荔枝园生境中只有两种蚂蚁占据优势地位,分别是小大头蚁(63.46%)和黑头酸臭蚁(32.50%)。

表2 3种生境中蚂蚁优势种Table 2 Ant dominant species in three habitats
2.2 蚂蚁的觅食动态分析
2.2.1蚂蚁对不同类型诱饵的觅食动态
在湖边生境,蚂蚁对蔗糖溶液的觅食数量在90 min达到最大;在树林生境,蚂蚁对蔗糖溶液的觅食数量在120 min达到最大;在荔枝园生境,蚂蚁对蔗糖溶液的觅食在150 min达到最大(表3)。尽管在3种生境蚂蚁觅食数量随时间变化偶有上下波动,蚂蚁对蔗糖溶液的觅食数量基本随时间增长而增加。但觅食蔗糖溶液的蚂蚁数量在每种生境的5个观察时间差异均不显著(湖边:F=0.648, df1=4, df2=10,P=0.641; 树林:F=1.461, df1=4, df2=10,P=0.285; 荔枝园:F=1.547, df1=4, df2=10,P=0.262)。在每个观察时间,3种生境间蚂蚁对蔗糖溶液的觅食数量差异均不显著(30 min:F=1.215, df1=2, df2=12,P=0.361; 60 min:F=0.786, df1=2, df2=12,P=0.498; 90 min:F=2.600, df1=2, df2=12,P=0.154; 120 min:F=0.691, df1=2, df2=12,P=0.537; 150 min:F=0.305, df1=2, df2=12,P=0.748)。

表3 不同生境蚂蚁对20%蔗糖溶液的觅食数量(Mean±SE)Table 3 Number of ants foraging for 20% sucrose solution in different habitats
在湖边生境,蚂蚁对火腿肠的觅食数量在75 min达到最大;在树林生境,蚂蚁对火腿肠的觅食数量在45 min达到最大;在荔枝园生境,蚂蚁对火腿肠的觅食在75 min达到最大(表4)。尽管在3种生境蚂蚁觅食数量随时间变化偶有上下波动,蚂蚁对火腿肠的觅食数量基本随时间增长而增加。觅食火腿肠的蚂蚁数量在湖边的5个观察时间差异显著(F=1.976, df1=4, df2=10,P=0.174)。但觅食火腿肠的蚂蚁数量在另外两种生境的5个观察时间差异均不显著(树林:F=0.799, df1=4, df2=10,P=0.553; 荔枝园:F=1.504, df1=4, df2=10,P=0.273)。蚂蚁在树林的觅食数量明显多于湖边和荔枝园。在第45 min,3种生境间蚂蚁对火腿肠的觅食数量差异显著(F=27.211, df1=2, df2=12,P=0.001)。但在其他4个观察时间,3种生境间蚂蚁对火腿肠的觅食数量差异均不显著(15 min:F=3.632, df1=2, df2=12,P=0.093; 30 min:F=4.152, df1=2, df2=12,P=0.074; 60 min:F=1.571, df1=2, df2=12,P=0.283; 75 min:F=1.098; df1=2, df2=12,P=0.392)。

表4 不同生境蚂蚁对火腿肠的觅食数量(Mean±SE)Table 4 Number of ants foraging for chicken and ham sausage in different habitats
2.2.2优势种蚂蚁觅食工蚁数量的时间动态
湖边3种优势蚂蚁在5个不同观察时间觅食蔗糖溶液的数量差异均不显著(表5)(小大头蚁:F=0.707, df1=4, df2=10,P=0.605; 黑头酸臭蚁:F=0.554, df1=4, df2=10,P=0.701; 未定种小家蚁:F=0.669, df1=4, df2=10,P=0.628)。在第120 min,黑头酸臭蚁觅食蔗糖数量明显高于另外两种优势蚂蚁,3种优势蚂蚁对蔗糖溶液的觅食数量差异显著(F=4.584, df1=2, df2=12,P=0.062)。在其他4个观察时间,小大头蚁对蔗糖溶液的觅食数量高于另外两种优势蚂蚁,但3种蚂蚁对蔗糖溶液的觅食数量差异均不显著(30 min:F=0.826, df1=2, df2=12,P=0.482; 60 min:F=1.898, df1=2, df2=12,P=0.230; 90 min:F=1.412, df1=2, df2=12,P=0.314; 150 min: F=2.876; df1=2, df2=12,P=0.133)。

表5 湖边不同蚂蚁对蔗糖溶液的觅食数量(Mean±SE)Table 5 Number of foraging for sucrose solution by different ants in Lakeside
湖边3种优势蚂蚁在5个不同观察时间觅食火腿肠的数量差异均不显著(表6)(小大头蚁:F=0.154, df1=4, df2=10,P=0.957; 黑头酸臭蚁:F=1.769, df1=4, df2=10,P=0.212; 未定种小家蚁:F=0.571, df1=4, df2=10,P=0.690)。在5个不同观察时间,3种蚂蚁对火腿肠溶液的觅食数量差异均不显著(15 min:F=1.306, df1=2, df2=12,P=0.338; 30 min:F=1.219, df1=2, df2=12,P=0.360; 45 min:F=2.482, df1=2, df2=12,P=0.164; 60 min:F=0.526; df1=2, df2=12,P=0.616; 75 min:F=0.074; df1=2, df2=12,P=0.929)。

表6 湖边不同蚂蚁对火腿肠的觅食数量(Mean±SE)Table 6 Number of ham sausage foraging by different ants in lakeside
树林优势蚂蚁白跗节狡臭蚁和宽结大头蚁在5个不同观察时间对蔗糖的觅食数量差异显著(表7)(白跗节狡臭蚁:F=1.783, df1=4, df2=10,P=0.209; 宽结大头蚁:F=3.015, df1=4, df2=10,P=0.071),小大头蚁在5个不同观察时间对蔗糖的觅食数量差异不显著(F=1.369, df1=4, df2=10,P=0.312)。白跗节狡臭蚁觅食蔗糖数量明显高于另外两种优势蚂蚁。在第150 min,3种优势蚂蚁对蔗糖溶液的觅食数量差异显著(F=13.602, df1=2, df2=12,P=0.006)。但在其他4个观察时间,3种蚂蚁对蔗糖溶液的觅食数量差异均不显著(30 min:F=1.000, df1=2, df2=12,P=0.422; 60 min:F=3.207, df1=2, df2=12,P=0.113; 90 min:F=0.036, df1=2, df2=12,P=0.965; 120 min:F=0.715, df1=2, df2=12,P=0.527)。

表7 树林不同蚂蚁对蔗糖的觅食数量(Mean±SE)Table 7 Number of sucrose foraging by different ants in forest
树林3种优势蚂蚁在5个不同观察时间觅食火腿肠的数量差异均不显著(表8)(白跗节狡臭蚁:F=0.363, df1=4, df2=10,P=0.829; 小大头蚁:F=1.185, df1=4, df2=10,P=0.375; 宽结大头蚁:F=1.336, df1=4, df2=10,P=0.322)。白跗节狡臭蚁觅食火腿肠数量明显高于另外两种优势蚂蚁。在第45 min,3种优势蚂蚁觅食火腿肠的数量差异显著(F=5.486, df1=2, df2=12,P=0.044)。但在另外4个不同观察时间,3种蚂蚁对蔗糖溶液的觅食数量差异均不显著(15 min:F=2.499, df1=2, df2=12,P=0.162; 30 min:F=1.912, df1=2, df2=12,P=0.228; 60 min:F=0.797, df1=2, df2=12,P=0.493; 75 min:F=1.447, df1=2, df2=12,P=0.307)。

表8 树林不同蚂蚁对火腿肠的觅食数量(Mean±SE)Table 8 Number of ham sausage foraging by different ants in forest
荔枝园优势蚂蚁黑头酸臭蚁在5个不同观察时间对蔗糖的觅食数量差异显著(表9)(F=1.759, df1=4, df2=5,P=0.214),小大头蚁对蔗糖的觅食数量差异不显著(F=0.495, df1=4, df2=5,P=0.740)。在5个不同观察时间,两种荔枝园优势蚂蚁对蔗糖的觅食数量差异均不显著(30 min:F=2.107, df1=1, df2=8,P=0.220; 60 min:F=1.905, df1=1, df2=8,P=0.140; 90 min:F=1.311, df1=1, df2=8,P=0.316; 120 min:F=0.024, df1=1, df2=8,P=0.885; 150 min:F=2.298, df1=1, df2=8,P=0.204)。

表9 荔枝园不同蚂蚁对蔗糖的觅食数量(Mean±SE)Table 9 Number of sucrose foraging by different ants in litchi orchard
荔枝园两种优势蚂蚁在5个不同观察时间对火腿肠的觅食数量差异不显著(表10)(小大头蚁:F=1.176, df1=4, df2=5,P=0.378; 黑头酸臭蚁F=2.53, df1=4, df2=5,P=0.136)。在5个不同观察时间,两种荔枝园优势蚂蚁对火腿肠的觅食数量差异均不显著(15 min:F=2.107, df1=1, df2=8,P=0.220; 30 min:F=1.905, df1=1, df2=8,P=0.140; 45 min:F=1.311, df1=1, df2=8,P=0.316; 60 min:F=0.024, df1=1, df2=8,P=0.885; 75 min:F=2.298, df1=1, df2=8,P=0.204)。

表10 荔枝园不同蚂蚁对火腿肠的觅食数量(Mean±SE)Table 10 Number of different ants foraging for ham sausage in litchi orchard
3 结论与讨论
在长期的进化中,不同蚂蚁形成了各自独特的觅食和生境偏好。本研究发现,虽然3种生境蚂蚁的种类数无显著差异,但是不同生境诱集到的具体种类则略有不同。比如,宽结大头蚁和黄尼氏蚁仅在树林生境中诱集到,而黑头酸臭蚁则只出现在湖边和荔枝园生境中。这可能是由于宽结大头蚁和黄尼氏蚁偏好在林边等相对荫蔽的生境中活动,而流浪蚁黑头酸臭蚁则喜欢在人类干扰活动强度较大的生境出没(King and Green, 1993;Katayama and Tsuji, 2010)。另外,树林生境中所诱集的蚂蚁数量最大,主要贡献来自白跗节狡臭蚁(67.36%),说明优势种的数量往往可能决定了整个生境的蚂蚁总数量(齐国君等,2015)。在蚂蚁类群中,种间的竞争往往受食物资源争夺量的直接影响(Wilderetal., 2011)。在多年无人管理的荔枝园内,黑头酸臭蚁、皮氏大头蚁PheidolepieliSantschi、伊大头蚁PheidoleyeensisForel为优势蚂蚁种群(吴碧球等,2008a)。在以白花鬼针草为主要植被的杂草地中,黑头酸臭蚁、亮红大头蚁PheidolefervidaF. Smith、皮氏大头蚁在该生境中占优势地位(黄煜权,2016)。这也说明,黑头酸臭蚁和皮氏大头蚁等入侵物种更倾向分布于被干扰强度较大的生境。
本研究发现大多数调查时间点上觅食工蚁数或同一时间点上不同蚂蚁的数量之间无显著差异,这可能是由于空间的异质性较大导致不同重复之间的差异较大所致。另外,温度、光照等环境因素以及投放诱饵数量有限、取样时间的控制不够准确等人为因素也可能是导致结果不准确的原因。在草坪、荒草地、荔枝园3种生境下,火腿肠、蜂蜜、黄粉虫幼虫及花生油4种诱饵的蚂蚁觅食研究中,生境对蚂蚁觅食效率的影响最大,其次是食物,而红火蚁入侵和生境两者间存在明显的交互作用,共同影响蚂蚁的觅食效率(吴碧球等,2008 b)。
不同蚂蚁在发现食物的速度以及攻击性方面的差异可能会导致觅食时间上的分化,如研究发现黑头酸臭蚁能比红火蚁更快发现食物,但是随着时间的延长红火蚁的竞争优势明显,逐渐赶走食物上的黑头酸臭蚁。在湖边用蔗糖溶液诱集蚂蚁,尽管小大头蚁在30 min、60 min、90 min、150 min时多于其他蚂蚁,但黑头酸臭蚁在120 min时占据显著数量优势。在树林用蔗糖溶液诱集蚂蚁,白跗节狡臭蚁在150 min时占据显著数量优势。但这不足以说明黑头酸臭蚁或白跗节狡臭蚁在觅食竞争后期会比其他蚂蚁占据更大优势,还需要设置更长的观察时间才能确定。
致谢:特别感谢广西师范大学陈志林博士在蚂蚁种类鉴定方面提供帮助。
