豆油添加对尼莫克汀发酵的影响
2020-09-11薛佳韵赵明霞王泽建孙慧婕王永红庄英萍
薛佳韵, 赵明霞, 王泽建, 孙慧婕, 王永红, 储 炬, 庄英萍
(1.华东理工大学生物反应器工程国家重点实验室,上海 200237;2.苏州大学医学部药学院,江苏苏州 215123)
尼莫克汀(Nemadectin)是一种米尔贝霉素类(Milbemycin)十六元大环内脂类抗生素,它是由蓝灰色链霉菌Streptomyces cyaneogriseus ssp.noncyanogenus所生产的一种广谱杀虫剂[1-2]。通过在尼莫克汀的第23 位引入=N−OCH3基团可半化学合成莫西克汀(Moxidectin),其具有高效、安全、环保以及不易产生抗药性等特点[3-7],因此在农用驱虫类杀虫剂领域具有广阔的应用前景[4,8-11]。此外,莫西克汀在治疗盘尾丝虫病方面也十分有效[6-7,12-13]。然而目前有关尼莫克汀的研究多集中于育种、分子机制以及毒理药理等方面[14-19],鲜有关于发酵调控方面的研究。
为了尽可能发挥出尼莫克汀生产菌株的性能,需要对尼莫克汀的发酵工艺进行优化。在部分链霉菌生产抗生素过程中,添加豆油能够有效提高目标产物产量。赵素敏[20]通过在发酵培养基中添加0.5 g/L豆油,从而使雷帕霉素的产量提高至243 mg/L;卞晨光等[21]通过添加豆油使红霉素的效价有效提高至5 221 U/mL,并推测豆油通过三羧酸循环代谢形成红霉素合成前体2-甲基丙二酰辅酶A 从而促进红霉素的合成;Wang 等[22]通过在发酵培养基中添加豆油,有效促进FK506(Tacrolimus)的合成,并且通过蛋白组学和代谢组学分析发现豆油不仅作为碳源被利用,亦可促进FK506 前体代谢。综合上述发现豆油的添加对部分大环内酯类抗生素的生产是有利的,然而目前鲜有关于豆油对尼莫克汀发酵的影响的研究。
本文通过考察豆油添加的不同质量浓度及时间,研究豆油对尼莫克汀发酵过程中菌体生长量以及产物浓度的影响,同时通过进一步考察5 L 反应器上尼莫克汀发酵过程的相关参数变化,合理分析了豆油在尼莫克汀合成过程中的作用。
1 实验部分
1.1 原料和试剂
1.1.1 菌株及豆油来源 蓝灰色链霉菌S-1(Streptomyces cyaneogriseus ssp.noncyanogenus) ,华东理工大学国家生化工程技术研究中心保存。
豆油选用金龙鱼精炼一级大豆油(益海嘉里食品营销有限公司),食用级豆油。
1.1.2 实验仪器 5 L 反应器(中国上海国强生化工程装备有限公司):微生物发酵;MAX300-LG 型尾气质谱仪(美国Extrel 公司):尾气中O2、CO2检测;1100 series 型高效液相色谱(HPLC,美国Agilent Technologies Inc.Agilent):尼莫克汀检测;LC-20A 高效液相色谱(日本岛津制作所):有机酸检测;显微镜(舜宇恒平科学仪器有限公司(上海):菌形分析。
1.1.3 培养基
(1)斜面/平皿培养基(g/L):麦芽糖1.3,酵母提取物4.0,硫酸镁0.5,碳酸钙1,琼脂粉20,pH 7.0。
(2)种子培养基(g/L):葡萄糖20,麦芽糊精20,黄豆饼粉15,棉籽饼粉15,K2HPO4·3H2O 1,MgSO4·7H2O 1,pH 7.0。
(3)发酵培养基(g/L):葡萄糖20,麦芽糊精85,黄豆饼粉25,玉米浆8,硫酸镁1.0,硫酸铜0.01,氯化钴0.002,硫酸锌0.001,硫酸锰0.001,钼酸钠0.002,碳酸钙6,消泡剂0.8(反应器上培养),pH 7.4。
1.1.4 培养方法
(1)孢子悬浮液制备
将蓝灰色链霉菌S-1 接种于斜面培养基中,在28 °C 下培养7 d,待孢子成熟时用无菌0.9 g/L 氯化钠溶液将孢子从斜面上刮洗下来并转接至无菌的带玻璃珠的250 mL 圆底三角瓶中,在220 r/min 转速下振荡3 min 以打碎菌丝释放出孢子,转接至无菌保菌管中备用。
(2)种子培养
使用500 mL 圆底三角瓶培养种子液。500 mL摇瓶装液量50 mL,并使用8 层纱布封口。将1 mL约3×107CFU/mL 的孢子悬液加入种子培养基中,在28 °C,240 r/min 转速下培养36 h,当离心压缩体积(PMV)达到原来体积的20%时,转接至发酵培养基中。
使用500 mL 圆底三角瓶或5 L 反应器进行发酵培养。500 mL 摇瓶装液量30 mL,并使用8 层纱布封口,5 L 反应器装液量为3 L。按8%的接种量将种子液转接到摇瓶发酵培养基中或反应器发酵培养基中,28 °C 下培养240 h。
1.1.5 豆油补加方案 在500 mL 摇瓶发酵过程中,初始0 h 以及菌体分化36 h 时一次性添加不同的最终质量浓度(0.25~15 g/L)的豆油,添加体积根据相应质量浓度计算。5 L 培养过程中实验是在初始发酵培养基中直接加入4.5 g 豆油。
1.2 参数测定
(1)菌体浓度测定
通过PMV 法测定发酵液中菌体浓度。取样10 mL发酵液于10 mL 离心管中,在4 000 r/min 转速下离心10 min后,将上清液与沉淀分离,沉淀物体积在10 mL 体积中所占百分数即为菌体PMV。
将沉淀用生理盐水清洗过后,重悬并抽滤,然后在烘干的滤纸(质量已知)上,85 °C 下烘至恒重后测定滤纸和沉淀质量,沉淀物的质量除以取样体积即为干重(DCW)。
(2)尼莫克汀产量测定
尼莫克汀产量通过HPLC 进行检测。采用安捷伦C18柱进行分离,流动相为1 mL/min 的甲醇和水混合液(甲醇和水体积比为9∶1)进行洗脱,柱温为30 °C,检测波长为245 nm。
(3)pH 测定
学生成绩的好坏,跟他的听课效率是密不可分的。而听课也要讲究方法。教师要引导学生明确听课的重要性,听课要专心致志,边听边思考,前后联系,不懂就问。同时还要将自己所听到的信息加以分析、归纳、整理、记录,这样才能更好地记忆与复习。
采用METTLER TOLEDO F120 型测试仪进行pH 测定。
(4)总糖浓度测定
总糖浓度采用3,5−二硝基水杨酸(DNS)法测定[23]。
(5)细胞活性测定
采用氯化三苯基四氮唑(TTC)法测定细胞活性:在发酵液中添加200 μL 发酵液、1 mL、0.1 mol/L Na2HPO4-NaH2PO4缓冲液(pH=7.0)和50 μL、0.5 g/L TTC 溶 液 混 匀 后 在40 °C 下 培 养3 h,然 后 通 过12 000 r/min 的转速离心去除上清液,在沉淀中加入1 mL 甲醇并超声30 min 以提取胞内TPF,最后使用96 孔板以及酶标仪在485 nm 下进行吸光度测定。
(6)胞外有机酸测定
胞外有机酸通过HPLC 进行分析。HPLC 配备离子排斥柱和吸收检测器分光光度计。采用安捷伦H 柱进行分离,流动相为0.5 mL/min 的10 mmol/L H2SO4,柱温为50°C,检测波长为210 nm。
(7)在线氧摄取率(OUR)检测

2 结果与讨论
2.1 摇瓶发酵中豆油添加对尼莫克汀产量的影响
2.1.1 摇瓶发酵中豆油不同添加模式对尼莫克汀效价的影响 通过在不同时间(0 h 和36 h)添加不同质量浓度(0.25,0.5,0.75,1,1.5,2.3,5,7,10,15 g/L)豆油,考察豆油添加对尼莫克汀的发酵影响,结果如图1所示。500 mL 摇瓶发酵周期为192 h。如图1 (a)所示,随着添加豆油质量浓度的提高,尼莫克汀产物质量浓度均呈先增长后降低的趋势,在所有实验组中,0 h 添加1.5 g/L 豆油以及36 h 添加1 g/L 豆油时尼莫克汀产物质量浓度最高,分别达到1 370 μg/mL 和1 355 μg/mL,比对照组(1 210 μg/mL)分别增长了13.22%和11.98%。当添加的豆油质量浓度超过5 g/L时,尼莫克汀的质量浓度则会急剧下降。
另一方面,豆油添加对菌浓维持有利。如图1(b)所示,所有的豆油添加均能有效提高菌浓。其中在初始培养基中添加1.5 g/L 豆油,发酵终菌浓比对照组增长了20.7%。但随着豆油质量浓度的进一步提高,菌浓会逐步下降。
2.1.2 摇瓶发酵中豆油不同添加模式对发酵比产率的影响 从基于菌体的产物得率Yp/c结果(图1(e)、(f))可知,豆油初始添加组的Yp/c普遍低于对照组,且在低质量浓度豆油范围下Yp/c与对照组相近,而随着豆油质量浓度的增加,Yp/c在不断降低。而在36 h 添加组中,Yp/c的变化趋势与总效价的变化趋势相似。其中,添加1 g/L 豆油条件下Yp/c达到63.59 mg/g,远高于对照组。
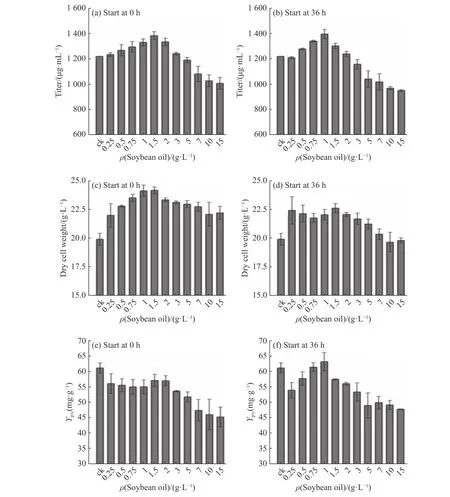
图1 不同豆油添加质量浓度下对蓝灰色链霉菌生长代谢影响Fig.1 Effects of soybean oil on S.cyaneogriseus growth and metabolism under different feeding modes
豆油作为一种碳源,在供氧充足的条件下很容易被好氧菌代谢利用,而在这个过程中,能够产生抗生素合成所需的酰基辅酶A 和大量能量。综上可以推断,0 h 添加的豆油主要被菌体用作碳源或提供初级代谢能量,有效提高菌体生长量,但随着豆油质量浓度进一步增加,过于丰富的碳源使得菌体短时间内生长旺盛,导致发酵液黏度大幅度上升,造成摇瓶上氧传递限制,反而对菌体次级代谢产生负面影响;而在36 h添加豆油时,低质量浓度的豆油添加直接被利用作前体或是次级代谢能量供给,提高了菌体产物合成能力,增强了菌体的前体代谢,同时也能有效维持菌浓。而高质量浓度的豆油可能导致气泡的快速聚并,从而引起氧传递限制,反而不利于菌体的代谢。
2.2 摇瓶豆油添加对发酵周期影响
此外,豆油添加对尼莫克汀发酵周期也有一定影响。分别在发酵了192 、216 、240 h 的摇瓶中进行取样,考察豆油添加对尼莫克汀的发酵过程菌体生长和产物合成的影响,结果如图2 所示。摇瓶发酵总周期为240 h。由图2 可知,豆油添加对整个尼莫克汀发酵周期有一定的延长作用。0 h 时添加低质量浓度豆油(0.25~1.5 g/L)可以合成大量前体物质和能量,使得产物合成期被延长,从而有利于产物合成,而36 h 时添加低质量浓度豆油则主要作为前体被利用,能够促进菌体次级代谢活性并延缓菌体死亡。当豆油添加质量浓度较高(7~15 g/L)时,由于摇瓶供氧能力有限,造成氧传递限制,不利于菌体呼吸代谢,致使合成期被延滞。随着豆油不断被消耗,浓度不断降低,氧传递限制逐渐被解除,菌体则又开始生长及合成,但由于整个发酵周期被延长,导致发酵结束时的效价仍处于较低水平,发酵过程生产效率较低。
2.3 5 L 反应器中豆油添加对尼莫克汀发酵过程的影响
为了进一步详细分析豆油对生产菌菌体生长和产物合成代谢的影响,本文在5 L 高级发酵罐系统中进行了过程动态多参数采集,结合离线(菌形、菌浓及尼莫克汀产量等)和在线参数(溶氧、氧摄取率及pH 等)分析豆油对生产菌代谢的影响及控制。摇瓶实验结果显示,初始添加1.5 g/L 豆油能有效提高生物量以及尼莫克汀的产量,因此,选择在5 L反应器上考察初始添加1.5 g/L 豆油对尼莫克汀发酵过程的影响。
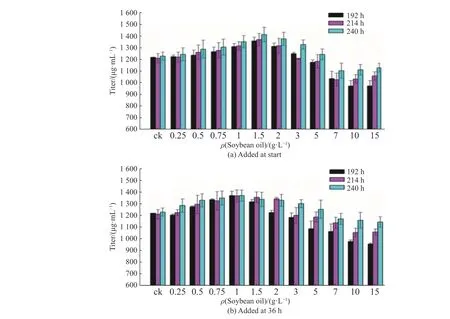
图2 不同豆油添加质量浓度下尼莫克汀的效价变化Fig.2 Changes of nemadectin titer at different mass concentrations of soybean oil
2.3.1 豆油添加对菌形的影响 图3 所示为不同培养条件下蓝灰色链霉菌的菌形变化。从图中可发现,生长期的菌丝逐渐伸长,菌丝体聚集成团粒,48 h时菌丝团开始分化,菌丝开始变短,断裂的小菌丝也有所增多。比较豆油组和对照组,结果表明72 h 以后,豆油组还有部分小菌丝团,而对照组已基本完全碎裂成小菌丝,这说明豆油的添加对菌丝团的维持有一定的作用。相关研究表明,部分链霉菌的产物合成与菌形变化存在一定关系,部分链霉菌在丝状菌丝状态下的产物合成能够有所增加[24],但如果菌丝断裂严重,对产物合成速率也会有显著的负面影响[25]。尼莫克汀发酵过程中后期,产物质量浓度较高的条件下菌丝会更长,同时也能维持一定的菌丝团形态[17],而豆油的添加能够较好地维持这种菌丝团,这有可能有效维持次级代谢产物的合成。
2.3.2 豆油添加对菌体生物量及细胞活性的影响 图4 示出了初始添加豆油(1.5 g/L)条件下5 L 反应器中蓝灰色链霉菌生长相关参数的变化情况。在菌体生物量方面,从图中可以发现,0~36 h 菌体处于生长期,菌浓不断提高,经过短暂稳定期后生物量从48 h开始逐渐下降并进入次级代谢,而此时尼莫克汀大量合成。比较豆油组和对照组的发酵过程曲线变化发现,在初级代谢阶段,添加豆油后菌体的生长量较对照组有明显增加(图4 (a)、(b)、(c)),豆油添加组最大菌浓(PMV 达到了38.76%)比对照组的最大菌浓(PMV 为30.05%)高出了29%,并且最高比生长速率分别达到0.024 9 h−1和0.023 7 h−1。当菌体进入尼莫克汀合成期后,豆油组的PMV 一直处于较高的水平,至发酵中后期PMV 仍能维持在22%以上,表明豆油的添加能够有效维持次级代谢阶段的菌浓。
另一方面,豆油组的细胞活性水平始终高于对照组(图4(d)),而葡萄糖消耗情况(图5)表明在生长期(0~48 h)间,豆油添加组的葡萄糖消耗速率要略高于对照组。这些都表明豆油添加能够有效提高蓝灰色链霉菌的初级代谢活性并提供能量,从而有利于菌体的生长和维持。
2.3.3 豆油添加对产物合成的影响 在产物合成方面(如图6 和表1),与对照组相比,豆油添加组尼莫克汀效价为1 475 μg/mL,相对于对照组(1 158 μg/mL)提高了27.37%。与此同时,豆油组的最高合成速率约为10.839 μg·/(mL·h),比对照组高了约24.61%,过程中合成速率也明显高于对照组。此外,豆油组的比产率至发酵结束依然能维持在约0.247 mg/(g·h),表明豆油的添加能有效维持次级代谢阶段的菌体活性,有利于产物合成。
2.3.4 豆油添加对细胞呼吸代谢相关参数的影响 除了上述尼莫克汀发酵过程中生理代谢相关的离线参数外,结合动态采集及分析的在线参数能够更全面地了解豆油对尼莫克汀发酵过程的影响。在实时采集的溶氧(DO)变化曲线中(图7(a)),两种培养工艺之间的变化趋势基本一致,但整个过程中,豆油组溶氧始终略低于对照组。实验过程中进气量、罐压、转速等均保持一致,由此可知豆油组对氧的需求要大于对照组组,且豆油组的生长状态优于对照组。

图3 不同培养条件下蓝灰色链霉菌菌体形态变化Fig.3 Morphological changes of S.cyaneogriseus under different conditions
另一方面,发酵过程中在线计算的OUR 变化曲线能够直接反映细胞的代谢活性情况,结果如图7(b)所示。如图所示,豆油添加后菌体的OUR 最高能够达到29.8 mmol/(L·h),也明显高于对照组的26.7 mmol/(L·h),表明豆油添加能够显著促进菌体的初级代谢活力,促使菌体浓度的增加。在192~240 h间,豆油添加组的OUR 能维持在9 mmol/(L·h)左右,而对照组则呈快速下降趋势,最低降至5 mmol/(L·h)左右,表明次级代谢阶段豆油组菌体活力优于对照组,能够较好地维持产物合成活性。结合比生长速率变化(图4 (c))、细胞活性结果(图4 (d))及残糖变化(图5 (a)),印证了豆油组条件下细胞代谢活性始终优于对照组,有利于维持产物的合成。
2.3.5 豆油添加对pH 及胞外有机酸的影响 5 L 反应器系统动态采集的pH 变化情况如图7(c)所示,对照组的pH 最低降至5.6,而豆油添加组的pH 最低则仅降至6.3;从24 h 起两组pH 均开始逐步回升,豆油组在66 h 左右pH 稳定在7.7 附近,而对照组则在约96 h 时才稳定在7.7 附近。
为了考察发酵过程中pH 产生变化的原因,对胞外有机酸进行了测定(图8 )。发酵过程中主要检测到的胞外有机酸为α-酮戊二酸(AKG)和丙酮酸(PYR)。通过考察这两者的变化情况,发现在生长期(0~48 h)胞外有机酸积累量较大,间接表明菌体的初级代谢活性较高,并且积累了大量能量,这也佐证了之前的推测。在48 h 后,两组的胞外丙酮酸质量浓度趋于0,而α-酮戊二酸的质量浓度也开始下降,表明菌体开始进入次级代谢状态。
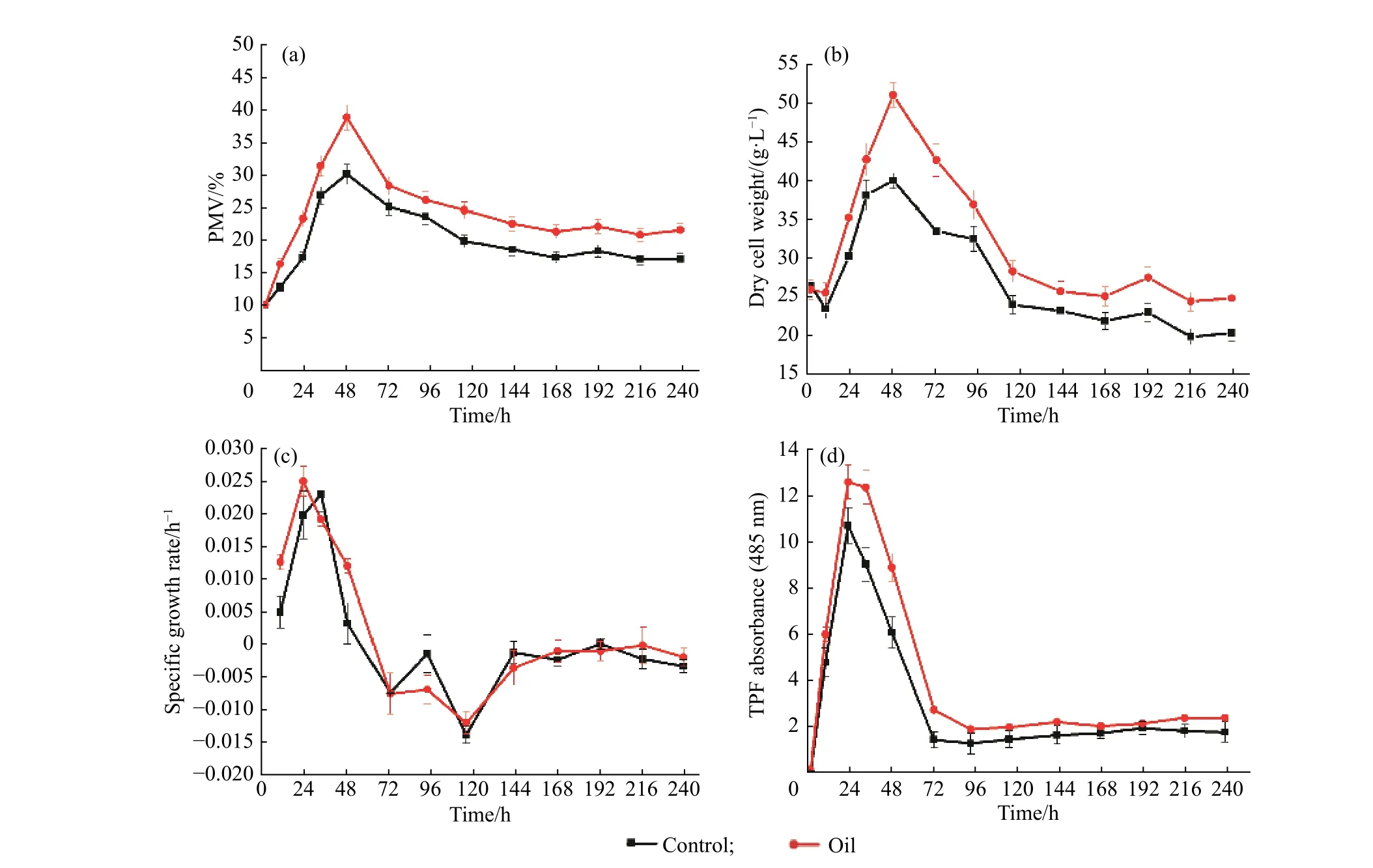
图4 初始添加豆油(1.5 g/L)条件下的5 L 反应器中蓝灰色链霉菌生长相关参数变化Fig.4 Effects of the initial addition of soybean oil (1.5 g/L) on the growth of S.cyaneogriseus during the 5 L fermentation process
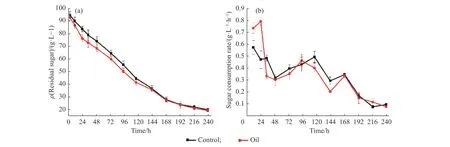
图5 初始添加豆油(1.5 g/L)条件下5 L 反应器中总糖的变化Fig.5 Effects of the initial addition of soybean oil (1.5 g/L) on the sugar consumption during the 5 L fermentation process
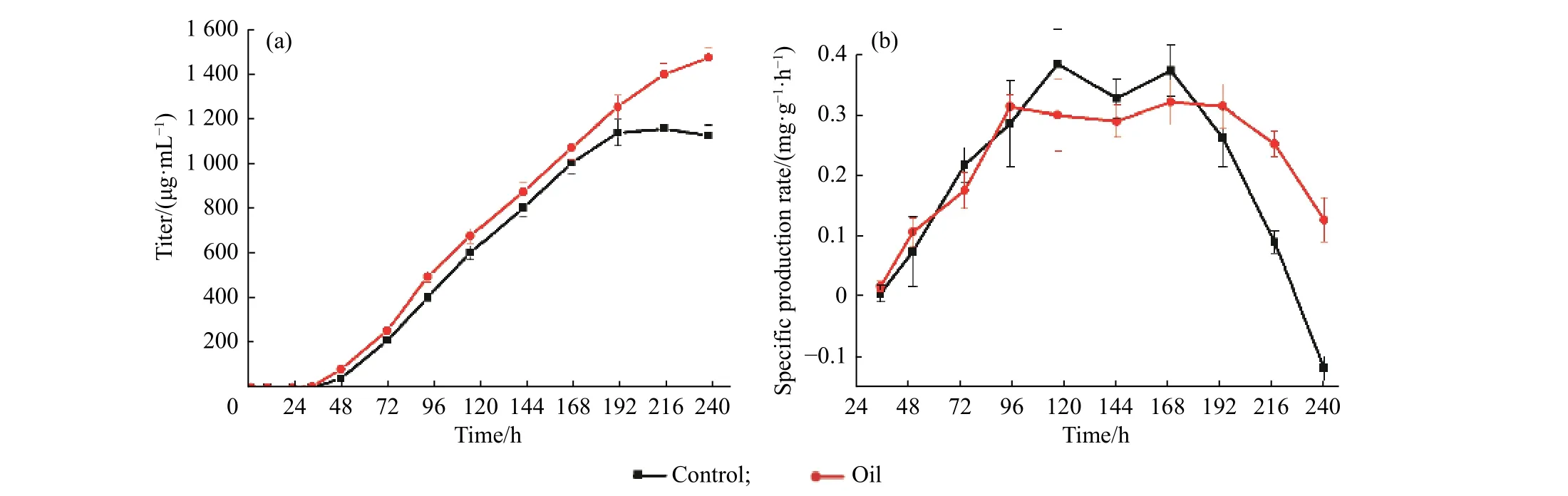
图6 初始添加豆油(1.5 g/L)条件下5 L 反应器中尼莫克汀产物质量浓度的变化Fig.6 Effects of the initial addition of soybean oil (1.5 g/L) on the mass concentration of nemadectin production during the 5 L fermentation process

表1 不同发酵条件下尼莫克汀发酵参数汇总表Table 1 Calculated parameters of the different fermentation conditions by S.cyaneogriseus
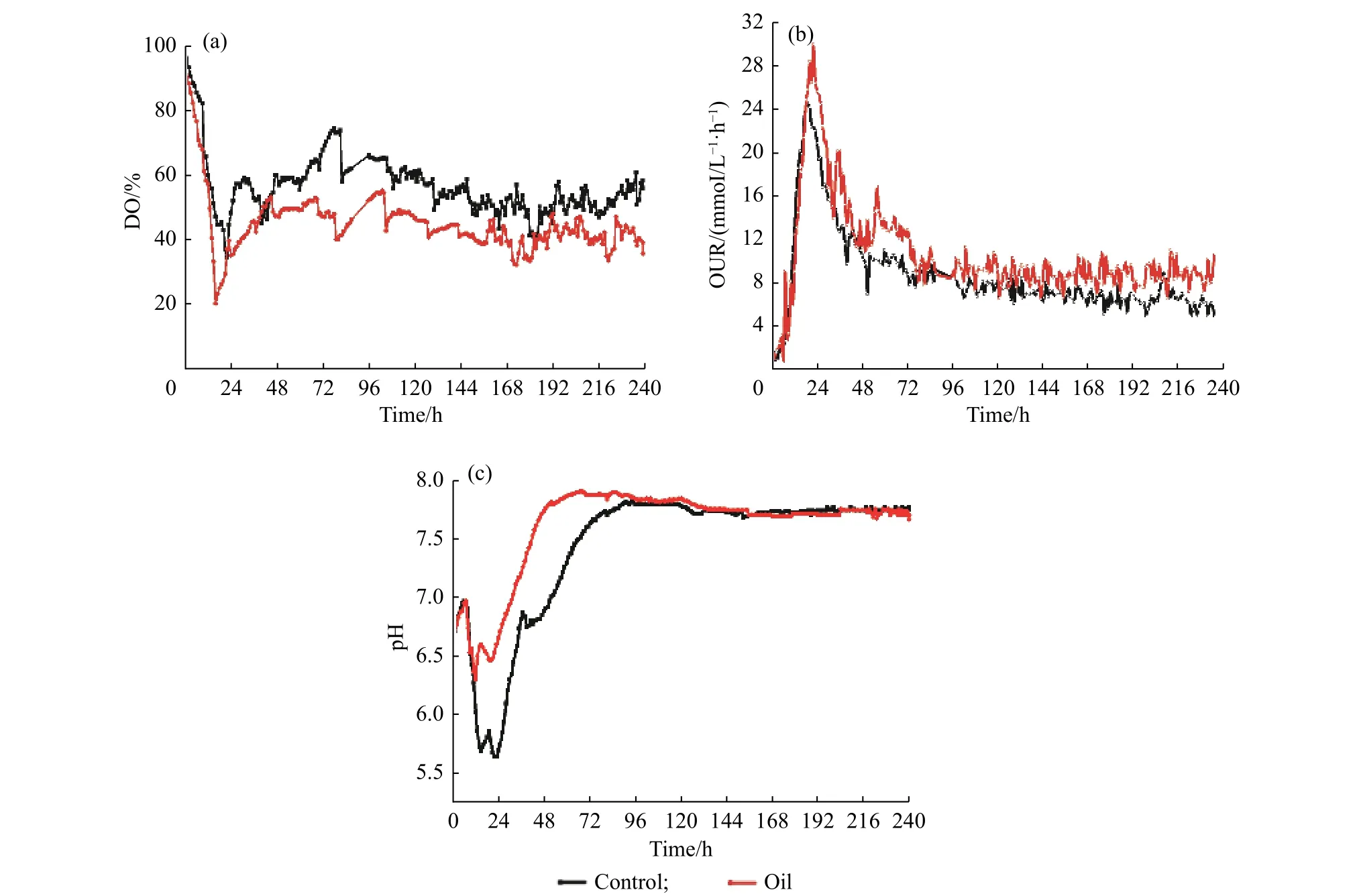
图7 初始添加豆油(1.5 g/L)条件下5 L 反应器中尼莫克汀合成相关的代谢参数变化Fig.7 Effects of the initial addition of soybean oil (1.5 g/L) on physiological parameters of nemadectin during 5 L fermentation process
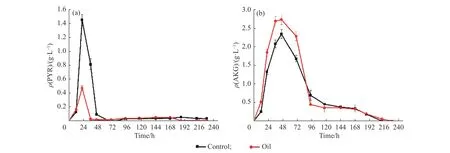
图8 初始添加豆油(1.5 g/L)条件下5 L 发酵过程中的胞外有机酸变化Fig.8 Effects of the initial addition of soybean oil (1.5 g/L) on extracellular organic acids during 5 L fermentation process
比较对照组和豆油组的胞外有机酸,发现0~48 h期间,对照组的胞外丙酮酸积累量高于豆油组,而α-酮戊二酸的含量则是豆油组高于对照组。丙酮酸是尼莫克汀合成前体辅酶A 物质的重要来源,而在供氧充足的条件下,豆油很容易通过Embden-Meyerhof-Parnas Pathway (EMP)以及Tricarboxylic Acid Cycle(TCA)途径产生丙酮酸,促进抗生素合成所需的酰基辅酶A 和大量能量的积累,保证有充足的能量能够顺利进入呼吸链从而进行氧化磷酸化,更有利于菌体的代谢以及增强尼莫克汀的合成途径。而由于对照组胞外丙酮酸的质量浓度较高,导致其pH 偏低,使得pH 回升会更慢。
3 结 论
本文通过在蓝灰色链霉菌的初始发酵培养基中添加豆油,得出如下结论:
(1) 添加低质量浓度豆油(0.25~5 g/L)可以有效提高菌体的生物量和产物合成单位,其作为碳源及能量供给,提高菌体的初级代谢活性,积累较多的生物量和能量,用于次级代谢过程中产物的合成,同时它也在一定程度上维持菌体的次级代谢活性,延长了尼莫克汀的有效发酵周期。
(2) 添加高质量浓度豆油时(7~15 g/L),主要可能是由于高质量浓度的豆油带来的氧限制问题不利于菌体的生长和代谢,只有当培养基中的豆油被不断消耗至氧限制解除才能进行产物合成,导致菌浓维持较高但产物合成较低。
(3) 通过离线和5 L 反应器上的在线参数采集,并结合相关离线参数进行分析,结果表明,在蓝灰色链霉菌菌浓方面,初始添加1.5 g/L 的豆油比对照组提高29%,尼莫克汀的产物浓度提高27.37%,达到1 475 μg/mL,说明豆油的添加能显著提高尼莫克汀的生物合成。此外,在线分析的细胞呼吸代谢相关参数表明,豆油添加能够有效提高蓝灰色链霉菌的初级代谢活性,其三羧酸循环效率更高,提供产生的大量的Adenosine Triphosphate (ATP)和Nicotinamide Adenine Dinucleotide (NADH)用于菌体的生长和维持,同时在一定程度上也能维持菌体的次级代谢活性,延长尼莫克汀的合成期。
综上所述,在尼莫克汀的发酵过程中,通过添加一定量的豆油能够较为有效地提高菌体合成,有利于生物量的积累和产物的生产。后续可以通过结合蛋白组学或代谢物组学深入研究豆油在蓝灰色链霉菌中的代谢机理,从而更全面、更深入地了解豆油对尼莫克汀的发酵影响。
