江西九连山不同恢复模式林分的物种多样性特征
2020-09-09张参参汪金松李张敏欧阳园丽陈伏生卜文圣
刘 斌,张参参,汪金松,李张敏,欧阳园丽,陈 维,陈伏生,卜文圣
(1. 江西农业大学林学院,江西九连山森林生态系统国家定位观测研究站,江西 南昌 330045;2. 生态系统网络观测与模拟重点实验室,中国科学院地理科学与资源研究所,北京 100101;3. 江西九连山国家级自然保护区管理局,江西 赣州 341700)
近几十年来,人类对森林资源以及农业土地的需求不断增加,使得大面积的天然林遭到毁坏,世界森林资源的面积和质量显著下降,大部分被人工林和次生林替代[1]。目前大多数人工林和次生林缺少原始森林的生境条件,生物多样性大大减少,生态系统稳定性降低,生态系统功能严重退化[2-3]。最近的研究表明,生物多样性对生态系统生产力有着显著的影响,其大小与气候和土壤肥力的影响相当[4]。生物多样性与生产力之间存在着全球一致的正相关性,这表明目前极高的生物多样性丧失速率将导致全球森林生产力的加速下降[5]。生境退化和森林过度砍伐的状况迫切需要采取干预措施来恢复生物多样性、生态系统功能和服务。随着原始林面积的不断减少和片段化,人类干预后的森林恢复速度以及能否恢复到干扰前的水平是目前极为关心的问题。
植被恢复是指运用生态学原理,通过保护现有植被、封山育林或营造人工林、灌、草植被,修复或重建被破坏的森林和其他自然生态系统,恢复其生物多样性及生态系统功能[6]。主要恢复模式有自然恢复与人工恢复模式两种,一般认为自然恢复模式的优点是投资少、效益高,森林的稳定性较好;而人工恢复模式的优点是恢复周期短、人为控制性高、具有较高的投入产出比[7]。目前主要的人工恢复模式是大量营造种类、结构单一的人工针叶纯林[8],部分皆伐或者间伐后的采伐迹地通过自然恢复模式成为次生林。我国于1998年启动的天然林保护工程,通过植树造林和自然更新来恢复森林生境,20年来使我国大大降低了森林面积的净损失率。然而,以下一系列问题都亟需评估和解决:如何评价森林恢复的成功?通过天然更新,人工林和次生林的林分结构及物种多样性如何变化[9]?人工恢复与自然恢复模式的优劣?
人工恢复模式构建的人工林通过快速生长提高了植被覆盖率,有助于木材的生产与供给,但并没有阻止森林的破碎化。由于新的森林大多是孤立的,这进一步破坏了残存的原始森林,而这些原始森林拥有最高的生物多样性[10]。研究普遍认为,在维持土壤和水分、保护生物多样性等方面,人工恢复植被均弱于自然恢复植被[11-13]。刘晓娟等[14]的研究证实了物种丰富度高的林分拥有更高的碳储量和碳通量,每增加一个树种,碳总存量增加6.4%。但也有研究表明,两种恢复模式林分的物种丰富度并没有显著差异[15],与自然恢复相比,人工造林往往有利于生物量和生物多样性的缓慢恢复,尤其是在演替的早期阶段[16]。
物种的更新状况对森林结构动态、演替进程和生态系统功能恢复都具有重要的作用。不同生长发育阶段的树木(幼树、小树和成年树)的多样性、密度、高度级结构等对未来群落的组成及群落动态稳定的维持和可持续发展具有深远影响[17]。径级结构是衡量植物群落生长发育状况和演替稳定性的重要指标。宫贵权等[18]发现小树个体会对邻域植物多样性产生促进作用,随着个体的变大促进作用会转变成抑制作用。闫满玉等[19]的结果显示不同大小的树木个体对邻域植物多样性的影响作用也因邻域植物个体大小而有所差别。群落物种多样性格局的形成不仅取决于生态过程,如以环境筛选为基础的确定性群落构建过程和以扩散限制为基础的随机性群落构建过程;还依赖于群落中的物种组成,如不同的功能群组成(生活型、耐荫性)或不同系统发育阶段的物种组成[20]。在森林群落中,光照是关键的生态因子,树木间的相互作用主要来源于对光资源的竞争[21]。耐荫性是植物对光环境适应的一种重要的生活史策略,能说明植物对外界光环境变化的响应[22]。在森林中,植物的功能性状如耐荫性、生长型等特征常常随着群落恢复的进程而发生显著变化,不同类型功能群随时间的更替决定着群落恢复的速率和方向[23]。
群落演替的速度和方向,以及驱动它们的生物非生物因素和过程,是群落生态学研究的核心[24]。本研究选择九连山自然保护区3个典型森林群落为对象,分别为未受人为干扰的原始常绿阔叶林、自然恢复32年的次生常绿阔叶林以及人工栽植杉木(Cunninghamia lanceolata(Lamb.) Hook.)后自然恢复36年的针阔混交林,在3个1 hm2固定样地设置和群落学调查的基础上,比较了次生林-原始林、杉木林-原始林两种不同恢复模式下,不同生长发育阶段树种的群落物种组成、结构和多样性的变化规律,以期回答以下两个问题:(1)两种恢复模式下不同生长阶段树种的林分结构、物种组成与原始林有何差异?(2)分析人工恢复模式与自然恢复模式的优劣性,探究两种恢复模式的恢复速度及恢复方向,为亚热带常绿阔叶林恢复和重建提供理论和实践依据。
1 材料与方法
1.1 研究区概况
本研究选址在江西省赣州市龙南县九连山国家级自然保护区境内,经纬度为24°31′~24°35′ N,114°26′~114°29′ E。保护区南北长度约17.5 km,东西宽度约15 km,总面积13 411.6 hm2[25]。九连山国家级自然保护区属中低纬度山地气候,温和湿润,年平均气温为16.4 ℃,年积温约4 852.5 ℃,1月份的平均气温6.8 ℃,7月份的平均气温24.4 ℃,区内年均降水量1 954.6 mm。研究区为中亚热带湿润常绿阔叶林与南亚热带季风常绿阔叶林的过渡地带,保存有完整的原生性常绿阔叶林。植被类型主要有常绿落叶阔叶混交林(次生阔叶林为主)、亚热带低山丘陵针叶林(人工林为主)、亚热带常绿阔叶林、竹林、山顶矮林及山地草甸[26]。
1.2 样地设置与调查
选择九连山国家级自然保护区内皆伐后人工种植杉木自然恢复36年的杉木针阔叶混交林、皆伐后自然恢复32年的次生常绿阔叶林以及未受人为干扰的原始森林(以下简称杉木林 CLP、次生林SF、原始林 PF),依据BCI大样地建设的技术规范[27],采用全站仪于2014年建立了3个1 hm2(100 m×100 m)样地,样地的4个角用水泥桩作永久标记。将1 hm2样地划分为25个20 m×20 m的小样方,对每个小样方内所有胸径(DBH)≥1 cm的木本植物个体使用铝牌进行编号,并在高度1.3 m处利用红漆进行标记。记录所有标记个体的树种名称、胸径、坐标、树高、枝下高和冠幅等信息。样地调查中所有植物的中文名和学名是基于2010版《中国生物物种名录》(中国科学院生物多样性委员会),样地基本环境条件见表1。

表1 样地基本环境条件Table 1 Basic environmental conditions of plots
1.3 数据处理
根据样地调查结果,按照树木的生理发育[18]、林业调查习惯及萌生能力[28-29],将所有被调查的木本植物个体按胸径大小划分成3个生长发育阶段:幼树(1 cm≤DBH<5 cm)、小树(5 cm ≤ DBH <10 cm)、成年树(DBH ≥10 cm)[30]。使用无度量多维标定(non-metric multidimensional scaling NMDS)方法来分析不同恢复模式群落3个生长发育阶段物种组成的差异,Chao的相似性系数被用于每个模型中的相似距离计算参数[31]。
按照《中国植物志》描绘的物种特性,结合种子特性、幼苗更新特点[23],按喜光的程度将出现在样地内的所有木本植物种类划分为3个功能群,即先锋种:如枫香(Liquidambar formosanaHance)、拟赤杨(Alniphyllum fortunei(Hemsl.) Makino)、南酸枣(Choerospondias axillaris(Roxb.) Burtt et Hill);非先锋喜光种:如木荷(Schima superbaGardn. et Champ.)、丝栗栲(Castanopsis fargesiiFranch.)、杜英(Elaeocarpus decipiensHemsl.)、黄杞(Engelhardtia roxburghianaWall.);耐荫种:如美叶柯(Lithocarpus calophyllusChun ex C. C. Huang et Y. T. Chang)、香桂(Cinnamomum subaveniumMiq.)、细齿叶柃(Eurya nitidaKorthals)、木莲(Manglietia fordianaOliv.)。
分别计算每个20 m×20 m样方不同生长发育阶段的α多样性(包括物种丰富度、Shannon-Wiener指数[32])以及3个功能群在不同恢复模式群落中的重要值。重要值=(相对多度+相对频度+相对显著度)/3。按照组合样方法计算群落不同面积尺度的种-面积曲线、种-多度曲线和种-物种等级曲线[33]。种-物种等级(Species-Species rank)曲线用于同时解释群落多样性的两个方面,即群落所含物种的丰富程度和均匀程度[34]。用Microsoft Excel 2013进行数据整理,R 2.12.0和 Origin 8.5进行数据分析和作图。
2 结果与分析
2.1 物种组成和林分密度
据数据统计分析,杉木林共计29科75种,其中多度最大的为杉科(32 %),其次为壳斗科(21 %)、樟科(16 %)、山茶科(7 %)、杜鹃花科(7 %)等;次生林共计26科73种,多度上依次为壳斗科(31 %)、山茶科(29 %)、樟科(13 %)等;原始林共计38科126种,多度最大的为山茶科(19 %),其次为杜鹃花科(18 %)、樟科(17 %)、壳斗科(11 %)。杉木林中占优势的树种主要有杉木、米槠(Castanopsis carlesii(Hemsl.) Hayata.)、润楠(Machilus nanmu(Oliver)Hemsley)、罗 浮 柿(Diospyros morrisianaHance)、毛棉杜鹃花(Rhododendron moulmainenseHook.)等;次生林中占优势的树种主要有木荷、丝栗栲、润楠、罗浮锥(Castanopsis fabriHance)、甜槠(Castanopsis eyrei(Champ.) Tutch.)等;原始林中占优势的树种主要有毛棉杜鹃花、君迁子(Diospyros lotusLinn.)、木荷、细齿叶柃、香桂等(表2)。因而,从物种更替的角度来看,杉木林-原始林的人工恢复过程中,杉科和壳斗科植物减少,山茶科和杜鹃花科植物增多;次生林-原始林的自然恢复过程中,壳斗科和山茶科植物减少,杜鹃花科和樟科植物增多。

表2 3种林分多度排前五的科信息Table 2 The messages of top five families among three stands

图1 3种林分不同生长发育阶段(s:幼树;t:小树,at:成年树)的无度量多维标定图。正方形代表杉木林样地,圆形代表次生林样地,三角形代表原始林样地Fig. 1 NMDS among three stands at different growth stages(s: sapling; t: treelet, at: adult tree). The square represents the Chinese fir forest, the circle represents the secondary forest, and the triangle represents the primary forest
NMDS排序可以明显地区分出原始林与两种恢复林分(图1)。说明随着植被恢复的进行,两种林分的物种组成逐渐向原始林方向靠近,但整个群落的物种组成还存在较大的差异。同时,NMDS排序还表明对于同一种恢复林分不同生长发育阶段的个体,其物种组成的差异小于不同林分之间的差异,说明整体上同一林分内部不同发育阶段的树种更新良好。
在群落植株个体数上,杉木林的总个体数(4 983)及幼树个数(2 571)与原始林相当(4 770和2 823),而次生林的总个体数(2 879)和幼树个体数(1 023)约占原始林的60 %和36 %。次生林中小树(926)及成年树个体数(930)与原始林相近(990和957);而杉木林的小树个体数(797)约占原始林的80 %,成年树个体数(1 615)约为原始林和次生林的1.7倍。原始林和杉木林在幼树阶段的胸高断面积(1.98和1.75 m2·hm-2)是次生林的2倍以上(0.77 m2·hm-2);在小树阶段,3种林分的胸高断面积差异不大(3.18、3.89和3.74 m2·hm-2);而在成年树阶段,次生林的(28.56 m2·hm-2)和杉木林(53.46 m2·hm-2)约占原始林胸高断面积(76.76 m2·hm-2)的37 %和70 %(图2)。
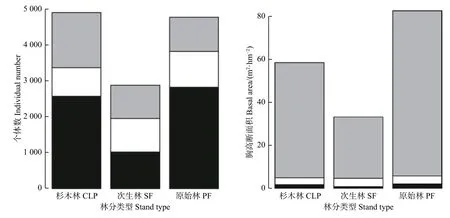
图2 3种林分不同生长阶段的个体数(左)和胸高断面积(右)。黑色面积表示幼树阶段,白色面积表示小树阶段,灰色面积表示成年树阶段Fig. 2 Number of individuals (left) and basal area (right) at different growth stages for the three stands. Black area represents sapling stage, The white area represents the treelet stage, The gray area represents the adult tree stage
2.2 物种多样性和优势度
原始林在种-面积和种-多度的物种累积速度要远大于两种恢复模式林分,而在种-物种等级曲线的物种多度下降速度低于次生林和杉木林(图3)。从种-面积累积曲线来看(图3 A.B.C),杉木林与次生林在幼树和小树两个阶段具有相似的变化趋势;而在成年树阶段,杉木林物种个数的积累速度要快于次生林,说明杉木林更有利于成年树稀有种的累积。在种-多度曲线上(图3 D.E.F),杉木林和次生林在小树及成年树阶段表现出相似的变化规律;但在幼树阶段,次生林物种数随着抽取个体数增加积累速度高于杉木林,说明次生林的幼苗库中物种个体分配更为均匀。3种林分类型的物种多度均随着物种等级的增高而降低(图3 G.H.I),在不同生长发育阶段中,原始林物种多度随物种等级的下降速度低于杉木林和次生林,而杉木林和次生林的变化趋势基本一致,表明两种恢复林分存在较明显的优势种,偶见种和稀有种较少,物种多样性更低;而原始林的曲线下降的更为平缓,意味着其拥有多个优势种,也有较多的偶见种和稀有种,物种组成更丰富,物种多样性更高。
从表3可知,杉木林和次生林不同生长发育阶段的物种丰富度明显小于原始林,但二者在小树阶段物种丰富度相似,次生林的幼树及成年树物种数少于杉木林,总体上杉木林和次生林的物种丰富度比较接近,约为原始林的55 %。总体上原始林的Shannon-Wiener指数显著地高于两种恢复林分,并且次生林显著地高于杉木林,但杉木林和次生林在不同生长阶段的Shannon-Wiener指数无显著差异。
2.3 群落结构
由图4可知,不同恢复模式林分各生长发育阶段的胸径和树高通常存在显著差异。次生林幼树胸径(2.88 cm)显著地大于杉木林(2.78 cm),但次生林和杉木林幼树的胸径与原始林(2.85 cm)无显著差异;次生林小树的胸径(7.18 cm)显著地大于原始林(6.93 cm)和杉木林(6.99 cm),但杉木林小树的胸径与原始林无显著差异;原始林成年树的胸径(24.85 cm)显著地大于两种恢复林分,并且杉木林成年树的胸径(19.39 cm)显著地大于次生林(18.31 cm)。次生林幼树的树高(5.04 m)显著地高于原始林(4.52 m)和杉木林(4.37 m),并且杉木林幼树的树高显著地小于原始林;次生林小树的树高(9.84 m)显著地高于原始林(6.56 m)和杉木林(7.27 m),并且杉木林小树的树高显著地大于原始林;次生林和杉木林成年树的树高显著地高于原始林(12.41 cm),但次生林(13.16 cm)和杉木林(13.74 cm)之间无显著差异。
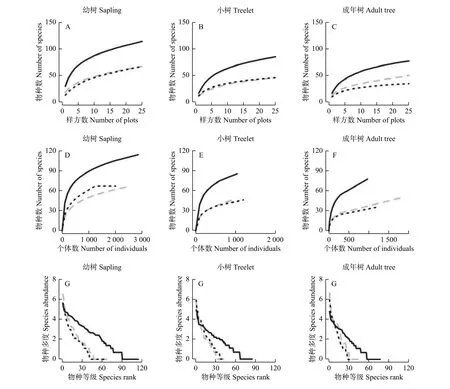
图3 3种林分不同生长发育阶段(幼树、小树、成年树)的种-面积累积曲线、种-多度曲线、种-物种等级曲线。灰色虚线表示杉木林,黑色实线表示原始林,黑色点线表示次生林Fig. 3 Species - area accumulation curve, species - abundance curve and species - species rank curve among three stands at three growth stages (sapling, treelet, adult tree). The dotted gray line indicates the Chinese fir forest, The black solid line indicates the primary forest, The black dotted line indicates the secondary forest

表3 3种林分不同生长发育阶段及总体的物种丰富度和Shannon-Wiener指数Table 3 The species richness and Shannon-Wiener index of the three stands at different stages and overall(Mean ± SE)
2.4 功能群变化
图5表明,3种林分在幼树阶段各个功能群的物种重要值较为接近;但在小树阶段,次生林先锋树种的重要值约占30%,是原始林和杉木林先锋树种重要值的2倍左右,并且3种林分耐荫性树种重要值排序为原始林>杉木林>次生林;在成年树阶段,与小树阶段相似,次生林先锋树种的重要值仍大于原始林和杉木林,但3种林分耐荫性树种重要值排序为原始林>次生林>杉木林。

图4 3种林分不同生长发育阶段植株胸径和树高的变异Fig. 4 Variations of DBH and height at three stages among three stands
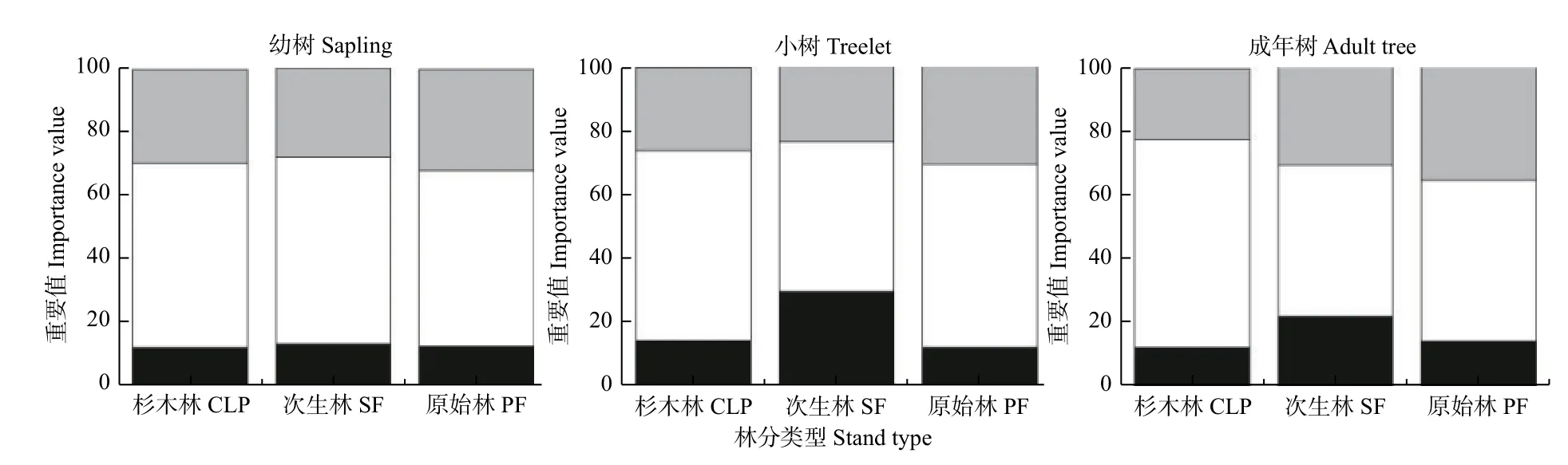
图5 3种林分不同生长发育阶段功能群的重要值变化Fig. 5 Variations of Importance value of three functional groups among three forests at different growth stages
3 讨论
3.1 物种组成
相对于森林植被结构的快速恢复,森林群落物种组成的恢复是最值得关注和研究的内容[35]。对比物种数量、群落结构和生物量积累等,物种组成的恢复速度更加缓慢[36]。杉木林-原始林的人工恢复过程中,杉科和壳斗科植物减少,山茶科和杜鹃花科植物增多;次生林-原始林的自然恢复过程中,壳斗科和山茶科植物减少,杜鹃花科和樟科植物增多。森林演替30多年后,两种林分的植被物种丰富度都达到了相当的水平,这表明无论是在人工恢复区还是自然恢复区,群落重组都在逐渐进行。演替过程中的光照资源变化可以通过改变种间关系以及限制更新方式等主导群落演替的方向,决定着群落的物种组成[37]。杉木林与次生林的物种组成均向着原始林方向靠近,但杉木林的物种组成与原始林更为接近,表明针阔混交林比阔叶林具有更强的正向演替趋势。
3.2 群落恢复进程
在个体数上,杉木林与原始林差异不大,并且成年树个体多于原始林;而次生林总个体数和幼树少于原始林,但小树和成年树的个体数与原始林差异不大。人工恢复如植树造林绕过了在早期演替中物种扩散和建立的障碍,增加了新生森林的物种个数的积累,加快了森林恢复[38]。相比之下,在自然恢复模式中,物种随机到达,受扩散和建立的限制[39]。物种个数和物种密度在分配上可能有着更多的变化,导致次生林物种个数与杉木林和原始林有显著差距。随着演替的进行,林分郁闭度逐渐增加,次生林进入自疏阶段,也会导致次生林物种个体数减少。从胸高断面积来看,在幼树和小树阶段,杉木林和原始林差异不大,成年树的约为原始林的70%;而次生林幼树胸高断面积约为原始林的一半,小树与原始林的差异不大,成年树仅为原始林的37%。这可能与人工种植的杉木有关,杉木是我国南方典型的速生造林树种,具有高的早期生长率,能迅速建立起林冠层[40]。恢复中的杉木林、次生林在物种个数和胸高断面积上与原始林存在较大差距。
次生林的Shannon-Wiener指数显著地高于杉木林,但在各年龄阶段间却无显著差异。物种多样性较低与先锋种的高度优势有关[41],杉木林中杉木依旧占据最高的个体数和重要值,根据Janzen-Connell效应,物种多样性与群落的同物种负密度有关[42],因此杉木林物种多样性与次生林有差异。两种恢复模式林分的物种多样性随着演替进行逐渐趋同于原始林,但与原始林的物种丰富度还存在明显的差距,杉木林和次生林由于许多演替后期物种的缺失,生态系统功能只是部分得到恢复[43]。根据各林分成年树的物种丰富度及Shannon-Wiener指数可知,原始林的林冠层具有更高的均匀度及物种多样性。常绿阔叶林乔木上层混交程度越高、林木分布状况越均匀,其林下层物种的均匀度指数与多样性指数则越大[44]。所以原始林在种-面积和种-个体数的物种累积速度要远大于两种恢复模式林分,而在物种多度-物种等级曲线的物种多度下降速度低于次生林和杉木林;杉木林与次生林在幼树和小树阶段的种-面积曲线具有相同的变化趋势。物种丰富度格局的形成也依赖于群落中的物种组成(如稀有种和常见种)[20],在成年树阶段,杉木林的物种密度要大于次生林,种-面积曲线的上升速度也高于次生林,这说明杉木林的成年树阶段拥有更多的稀有种。
3.3 群落演替
群落喜光性是群落恢复过程中的一个重要适应性特征[28]。随着演替的进行,林分郁闭度变大,导致一些阳性树种幼苗、幼树因光照不足而被淘汰出群落,阴性树种开始在群落中定居,因此呈现出喜光植物比例下降、耐荫植物比例上升的规律[45]。次生林林下层树种(幼树、小树)先锋种和非先锋喜光种的重要值高于杉木林与原始林,表明次生林的正向演替进程慢于杉木林,自然恢复30年左右的次生林中,先锋树种在群落组成中依旧占据主导地位[46]。恢复初期,阳性的先锋树种通过萌生个体扩展林冠面积来适应或利用高光条件;随着自然演替的进行,群落环境变得荫蔽,喜光树种的主导地位逐渐减弱,大量中性及耐荫树种侵入并占据较大的生态位[47],因此耐荫的后期物种能在先锋树种死亡后空出的位置中成功定居[48]。次生演替进程中,负密度制约效应在幼树和小树阶段的存活中起着重要的作用,激烈的种内竞争会抑制树木个体的生长发育[29], 容易导致林分发生自疏现象,形成大小不一的林窗、林隙,使得林下层光照更为充足。群落个体数减少后,各树种生态位变得更为宽泛,种内、种间竞争减弱。因此,林分自疏后创造的资源空间和林分密度降低可能是次生林幼树、小树的DBH和树高都要大于杉木林与原始林的原因。原始林、杉木林的径级结构接近倒“J”型,群落更新良好;而次生林各生长阶段分布均匀,群落稳定性较差。原始林为地区顶级群落,垂直结构更为立体,拥有复层林冠层及大径级成年树。因此成年树阶段原始林的胸径显著高于两种恢复林分。杉木林和次生林中缺乏大径级和超高的个体,例如DBH≥40 cm、树高>20 m的个体主要分布在原始林中。
4 结论
在恢复30多年后,人工恢复与自然恢复群落的物种组成、物种多样性、物种个数都能朝着当地顶级群落发展,但杉木林的演替进展更为顺利。相对于自然恢复的次生林,人工恢复的杉木林在短期恢复中有利于物种个体数及生物量的积累,有利于稀有种的定居;但自然恢复的次生林更有利于物种多样性的恢复,尤其是在林下层。为此本研究认为以快速积累生物量为目标的固碳森林,选择人工恢复模式更为适宜;而以恢复生物多样性为目标的多功能森林则选择自然恢复模式。对于本研究而言,由于缺乏较长恢复时间的物种数据,未对不同演替阶段林分进行连续观测,很难预测其最终的演替方向。在更长时间的恢复中,人工模式是否能优于自然模式,从而更快地接近顶级群落的物种组成和群落结构仍存在很大的不确定性。
