不同年龄人群来源长双歧杆菌体外生物学特性研究
2020-09-01赵建新
闫 爽 杨 波 赵建新 张 灏 陈 卫
(江南大学食品学院 江苏无锡214122)
双歧杆菌(Bifidobacteria)是人体肠道菌群中一类重要的优势微生物, 与其宿主之间存在复杂的相互作用关系[1]。某些特定的双歧杆菌菌株被证明具有拮抗肠侵袭性致病菌,抑制结直肠癌,抑制炎症性肠病以及缓解便秘等功能[2-5]。
尽管有大量关于人体肠道微生物组成的研究, 然而关于肠道共生菌和宿主相互作用的分子机制还不十分明确[6]。 有研究提出,共生菌的细胞被膜成分,包括蛋白质和碳水化合物,是微生物与其宿主之间发生相互作用的重要物质基础[7-9]。 胞外多糖是由一些特定菌株产生的一类碳水化合物聚合物,可附着于菌体表面形成一层被膜(荚膜多糖)或者释放到周围环境中[10]。 双歧杆菌的荚膜多糖可通过改变菌体表面的物理性质维持菌体与宿主的共生关系。此外,某些双歧杆菌产生的胞外多糖(Exopolysaccharides,EPS)可提高菌体对胃肠道环境的耐受性, 保护菌体免受宿主免疫系统的影响,同时可对宿主产生免疫调节等多种作用[11-12]。双歧杆菌所产的EPS 可能是其发挥益生功能的物质基础之一[13]。
短链脂肪酸(Short-chain fatty acid,SCFA)是人体肠道菌群的重要代谢产物, 具有为肠上皮细胞提供能量,促进肠上皮细胞增殖和矿物质吸收,缓解腹泻与便秘,抑制致病菌等多重生理活性[14]。近年来大量研究表明, 短链脂肪酸具有免疫调节功能,对维持免疫稳态起关键作用[9]。 双歧杆菌所产的短链脂肪酸主要为乙酸, 它可保护宿主免受肠道致病菌感染, 同时可被肠道中产丁酸菌利用产生丁酸,发挥抗炎等生理活性[2,15]。
本研究从婴幼儿、 成年人和老年人的粪便中筛选长双歧杆菌(B. longum),分析不同菌株的体外特性,包括生长特性、糖发酵特性、产短链脂肪酸和胞外多糖的能力, 并比较菌株胞外多糖合成关键基因——引导型糖基转移酶(Priming glycosyltransferase,pGT)基因的序列及其进化关系,为双歧杆菌特定菌株的筛选以及胞外多糖相关研究提供数据参考。
1 材料与方法
1.1 样品采集
共采集38 份不同年龄段健康志愿者的粪便样品。 其中男性20 名,女性18 名;小于3 岁的婴幼儿14 名,30~50 岁的中年人11 名, 大于60 岁的老年人13 名。将采集的粪便样品置于50 mL 无菌离心管中,添加30 mL 灭菌的30%甘油溶液(含0.05%半胱氨酸盐酸盐)混匀,在4 ℃下暂时保存,并于48 h 内运送至实验室,-80 ℃条件下保藏。
1.2 主要试剂和设备
MRS 培养基,青岛海博生物技术有限公司;半胱氨酸盐酸盐、硫酸、苯酚,国药集团;莫匹罗星、细菌基因组DNA 快速抽提试剂盒,上海生工生物工程有限公司;2×Taq MasterMix,北京康为世纪生物科技有限公司; 细菌16S rRNA 基因通用引物27F (5'-AGA GTT TGA TCC TGG CTC AG-3′)和1492R (5′-ACG GCT ACC TTG TTA CGA CTT-3′),上海桑尼生物科技有限公司;阿拉伯树胶、阿拉伯糖、纤维二糖、D-葡萄糖胺、D-葡萄糖醛酸酯、D-甘露糖、 松三糖、D-木糖, 美国sigma公司;低聚果糖、低聚半乳糖、低聚异麦芽糖,上海源叶生物科技有限公司。本试验所用MRS 培养基在配制时均添加0.05%的半胱氨酸盐酸盐。
超低温冰箱, 美国Thermo scientific 公司;超净工作台, 江苏苏净集团有限公司;Whitley DG250 厌氧工作站, 英国Don Whitley Scientific公司;PCR 热循环仪, 美国Biorad 公司;JY92-IIDN 超声波细胞破碎仪, 宁波新芝生物科技股份有限公司,UV1800 紫外-可见分光光度计,日本岛津公司。
1.3 双歧杆菌的分离纯化
冻存的粪便样品在室温下融化,混匀。将粪便悬液用无菌生理盐水以10-1梯度稀释。 选取适当稀释度的稀释液涂布MRS 平板(添加0.05%半胱氨酸盐酸盐和100 mg/L 莫匹罗星), 置于37 ℃厌氧工作站中培养72 h。长出单菌落后,挑取不同形态的菌落继续划线纯化,最终得到纯培养物,扩增16s rDNA,将扩增产物送至华大基因测序以确定物种,并保藏菌种。
1.4 生长曲线的绘制
待测菌株在液体培养基中连续培养3 代后,以2%的接种量接种至MRS 液体培养基 (添加0.05%半胱氨酸盐酸盐)中,于37 ℃厌氧工作站中培养。 培养过程中每2 h 取样1 次,测定发酵液的pH 值和波长600 nm 处的OD 值, 直至进入稳定期。
1.5 对数末期菌落数的测定
取对数生长末期的发酵液, 用无菌生理盐水以10-1梯度稀释, 选取适当稀释度的稀释液涂布MRS 平板,置于37 ℃厌氧工作站中培养72 h。 长出单菌落后,进行菌落计数。发酵液中的菌落数以CFU/mL 表示。
1.6 糖发酵特性分析
将不同种类的糖配制成溶液, 用0.22 μm 的微孔滤膜过滤除菌,以1 g/L 的质量浓度添加到不含糖的MRS 液体培养基中,另添加少量溴甲酚紫溶液, 作为酸碱指示剂。 待测菌株培养至对数末期, 离心收集菌体, 用无菌生理盐水吹洗菌体2次,用等体积的生理盐水重悬菌体,以2%的接种量将菌悬液接种到上述含不同碳源的MRS 培养基中,在37 ℃的厌氧工作站中培养24~48 h,记录培养基的变色情况。 双歧杆菌能生长的培养基变为黄色,双歧杆菌不能生长的培养基仍未紫色,由此判断菌株能否利用相应的糖。 以含葡萄糖的MRS 培养基和不含糖的MRS 培养基分别作为阳性和阴性对照。
1.7 发酵上清液中短链脂肪酸的测定
将菌株在MRS 液体培养基中培养至对数末期,6 000×g 离心10 min, 上清液用于测定短链脂肪酸(乙酸、丙酸、丁酸、异丁酸、戊酸和异戊酸)的含量。短链脂肪酸的提取和GC-MS 测定参考Mao等[16]的方法。 采用外标法计算各短链脂肪酸的质量摩尔浓度, 最终结果以单位生物量产生的短链脂肪酸表示(μmol/g 生物量)。
1.8 胞外多糖的测定
胞外多糖的测定分为细胞表面多糖(EPS-b)测定和游离胞外多糖(EPS-r)测定。将双歧杆菌菌株在MRS 液体培养基中培养至对数末期, 取10 mL 发酵液,6 000×g 离心15 min,收集菌泥和上清液分别用于测定EPS-b 和EPS-r。 胞外多糖的提取和测定参考Tallon 等[10]的方法。 测定结果以单位生物量产生的EPS 毫克数表示(mg/g 生物量)
1.9 引导型糖基转移酶 (pGT)基因的PCR 扩增、测序和生物信息学分析
根据NCBI 数据库, 长双歧杆菌的pGT 基因分为cpsD 基因和rfbP 基因。 根据数据库中的序列, 设计pGT 基因特异性引物如下:cpsD-F (5'-TTCTCYGTGCGCATGGAATC-3′)和cpsD-R(5'-CCCATAATSGACCAGTTCTGCAC-3′);rfbP-F(5'-GATTCYGAGACCATGCGTAC-3′)和rfbP-R(5'-GCATARTCCGACTGTTCCTGAG -3′)。 待测菌株培养至对数末期,离心收集菌体,采用细菌基因组DNA 快速抽提试剂盒提取细菌总DNA。以细菌基因组DNA 为模板, 采用pGT 基因特异性引物PCR 扩增,将PCR 产物送至华大基因测序,所得序列用Clustal X2 软件比对,用MEGA5.1 软件的neighbor-joining 方法建立系统进化树。 选取标准菌株长双歧杆菌NCC2705 的pGT 序列作为参考序列。
1.10 数据分析
所有指标均做3 次重复试验, 所得数据用SPSS 20.0 软件统计分析,独立样本t-检验用于分析组间显著性差异,P<0.05 被认为具有显著性差异。
2 结果与讨论
2.1 不同长双歧杆菌菌株的生长特性
根据16s rDNA 序列比对结果,选取38 株分离自不同粪便样品的长双歧杆菌(表1)做后续分析。
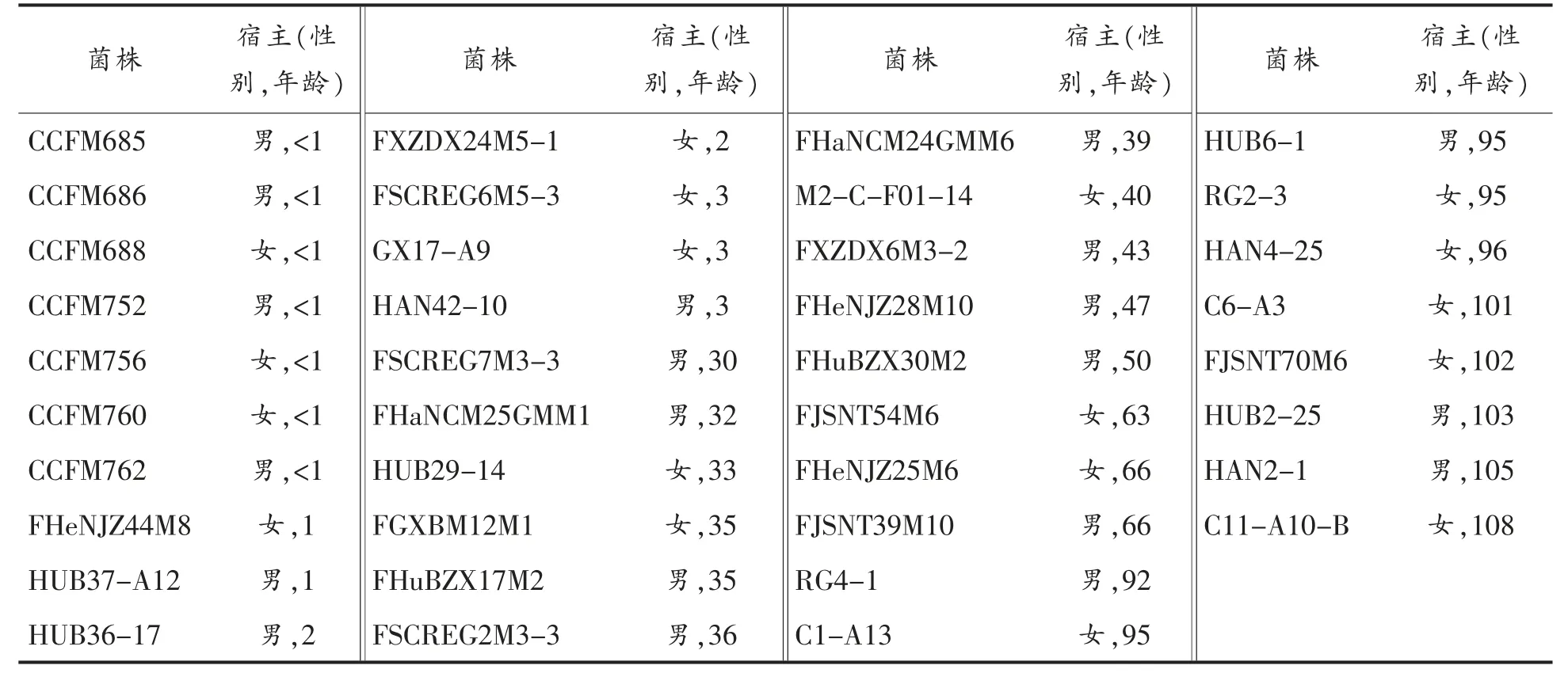
表1 筛选出的长双歧杆菌菌株及其宿主信息Table 1 Isolated strains and the information of their corresponded hosts
由于不同菌株对代谢底物的利用能力不同,其生长速率存在差异, 因此首先测定各菌株在MRS 培养基中的生长情况。 由图1a 可知,不同菌株在MRS 培养基中的生长特性具有显著差异。 整体来看,婴幼儿源菌株的生长速率相对较慢,延迟期为6~8 h; 而老年人源的菌株生长速率相对较快,延迟期为4~6 h,其中HUB6-1 的延迟期只有2 h,10 h 后进入稳定期, 而其它多数菌株在20 h左右进入稳定期。 另外,进入稳定期后,各菌株发酵液的OD 值也有显著差异,婴幼儿源菌株的OD值整体低于老年人源菌株的OD 值, 对各菌株对数末期活菌数的计数结果也显示, 婴幼儿源菌株的CFU 值显著低于老年人源的菌株(图1b)。 然而, 不同性别人群来源的菌株之间没有类似的差异性。 该结果可能是由于不同年龄宿主来源的菌株在体外培养条件下对培养基中营养成分利用能力存在差异所致。
2.2 不同长双歧杆菌菌株对碳源的利用特性
根据伯杰氏系统细菌学手册所述, 长双歧杆菌又分为长亚种(B. longum subsp. longum)、婴儿亚种 (B. longum subsp. infantis)和猪亚种((B.longum subsp. suis), 不同亚种间通过16s rDNA序列很难区分。 由于不同亚种在糖发酵特性上存在差异, 因此菌株的发酵特性可以作为区分亚种的方法。图2 列出38 株长双歧杆菌对不同碳水化合物的利用情况。 由图2 可知,只有FHeNJZ44M8不能利用阿拉伯糖而可以利用D-葡萄糖醛酸酯。根据伯杰氏手册, 作者认为该菌株为长双歧杆菌婴儿亚种。有研究提出,长双歧杆菌长亚种可分为2 种亚型(biovar),其中亚型A 可以发酵甘露糖,而亚型B 不能发酵甘露糖[17]。 本研究发现,婴幼儿来源的菌株多数可以利用甘露糖,14 株菌株中只有2 株不能利用甘露糖;而成年人来源的24 个菌株中,只有6 株可以利用甘露糖。多数菌株可以发酵本试验中的3 种低聚糖, 只有C1-A13、FJSNT70M6、RG2-33 菌株对低聚糖的利用能力较差。多数菌株不能利用阿拉伯树胶、纤维二糖和D-葡萄糖胺。
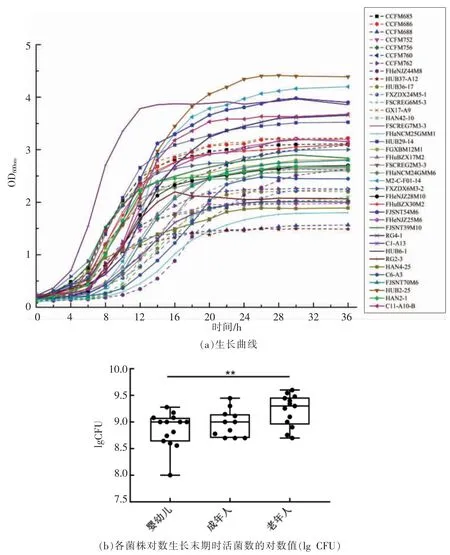
图1 不同长双歧杆菌菌株的生长曲线Fig.1 Growth curve of different strains in MRS broth
2.3 不同长双歧杆菌发酵上清液中SCFA 的组成
各菌株发酵上清液中短链脂肪酸的组成如图3a 所示。 长双歧杆菌发酵上清液中主要的短链脂肪酸为乙酸,另有极少量的丙酸、丁酸和戊酸,后几种短链脂肪酸的含量与新鲜MRS 培养基中的含量基本一致,因此可认为双歧杆菌在MRS 培养基中产生的短链脂肪酸只有乙酸, 扣除培养基中本身存在的乙酸, 即得各菌株产乙酸的能力(图3b)。 双歧杆菌对六碳糖的代谢途径为“双歧杆菌途径”(Bifidus pathway), 代谢产物主要为乙酸和乳酸,二者比例为3∶2[17]。 乙酸是双歧杆菌的主要代谢产物之一, 它在保护肠道免受侵袭性治病感染方面起到重要作用[2]。不同菌株之间产乙酸的能力有一定的差异, 其中HUB37-A12、C1-A13、HAN4-25 短链脂肪酸的产量明显高于其它菌株,而不同年龄以及性别人群来源的菌株间并没有呈现规律性差异。

图2 各菌株对不同碳水化合物的利用情况Fig.2 Carbohydrates fermentation test of different strains
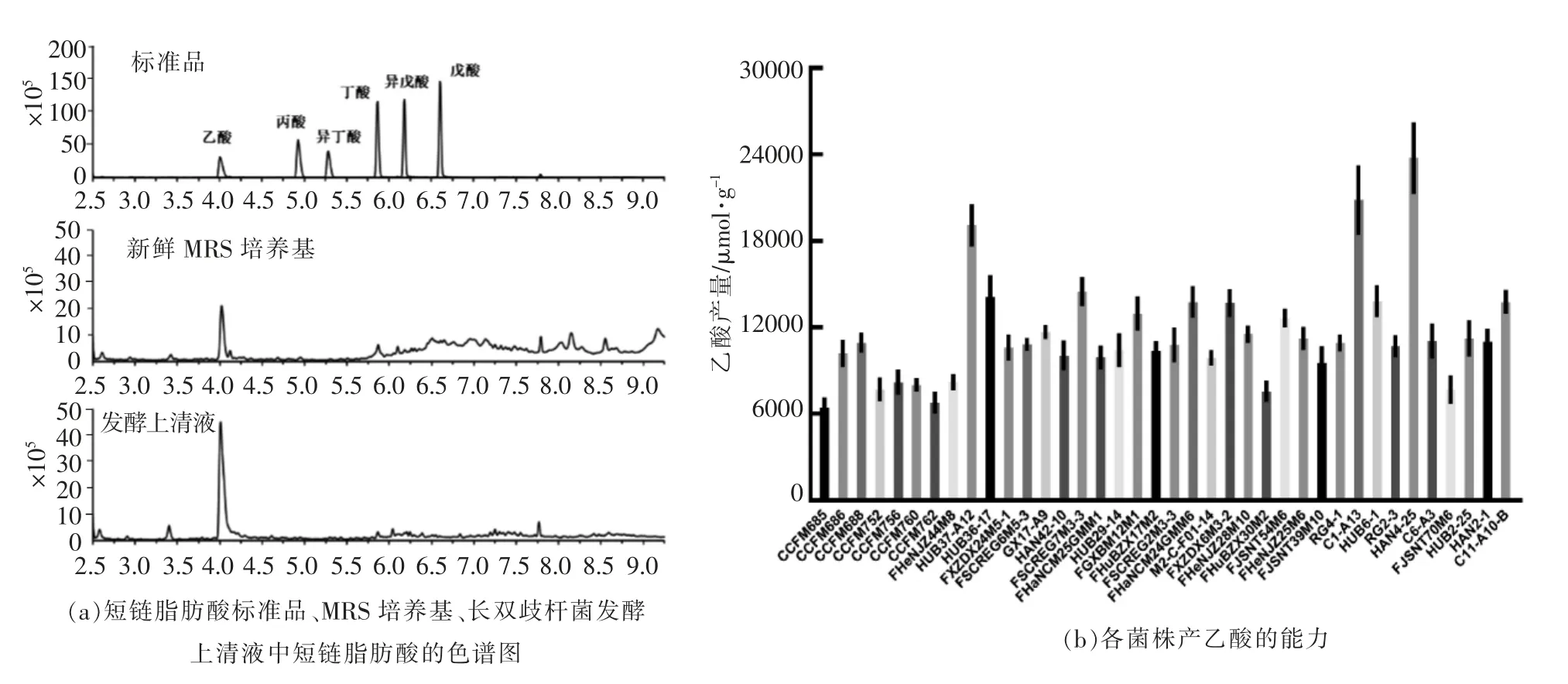
图3 不同菌株发酵上清液中短链脂肪酸(SCFA)的组成Fig.3 Composition of SCFA in culture supernatant of different strains
2.4 不同长双歧杆菌菌株的胞外多糖产量
胞外多糖作为肠道微生物与宿主肠道之间相互作用的重要介质, 对于宿主具有一定的免疫调节作用。 本研究测定了不同菌株的菌体表面多糖(EPS-b)和游离胞外多糖(EPS-r)的产量。 由表2可知, 不同菌株之间胞外多糖的产量具有明显差异。 其中CCFM760、HUB37-A12、HAN4-25 几乎不产胞外多糖, 而HUB2-1 和HUB6-1 被检测到表面胞外多糖(EPS-b)。 另外,将菌株按照宿主年龄分组后发现, 婴幼儿来源菌株的细胞表面多糖(EPS-b)产量低于中年人和老年人来源菌株,3 组的平均值分别为9.93,21.13,22.58 mg/g,其中婴幼儿源和老年人源菌株的细胞表面多糖(EPS-b)产量存在统计学显著性(P<0.05)(图4)。这些特性的差异可能与不同来源的菌株适应其宿主肠道环境有关[18-19]。而不同性别人群来源的菌株之间没有规律性差异。
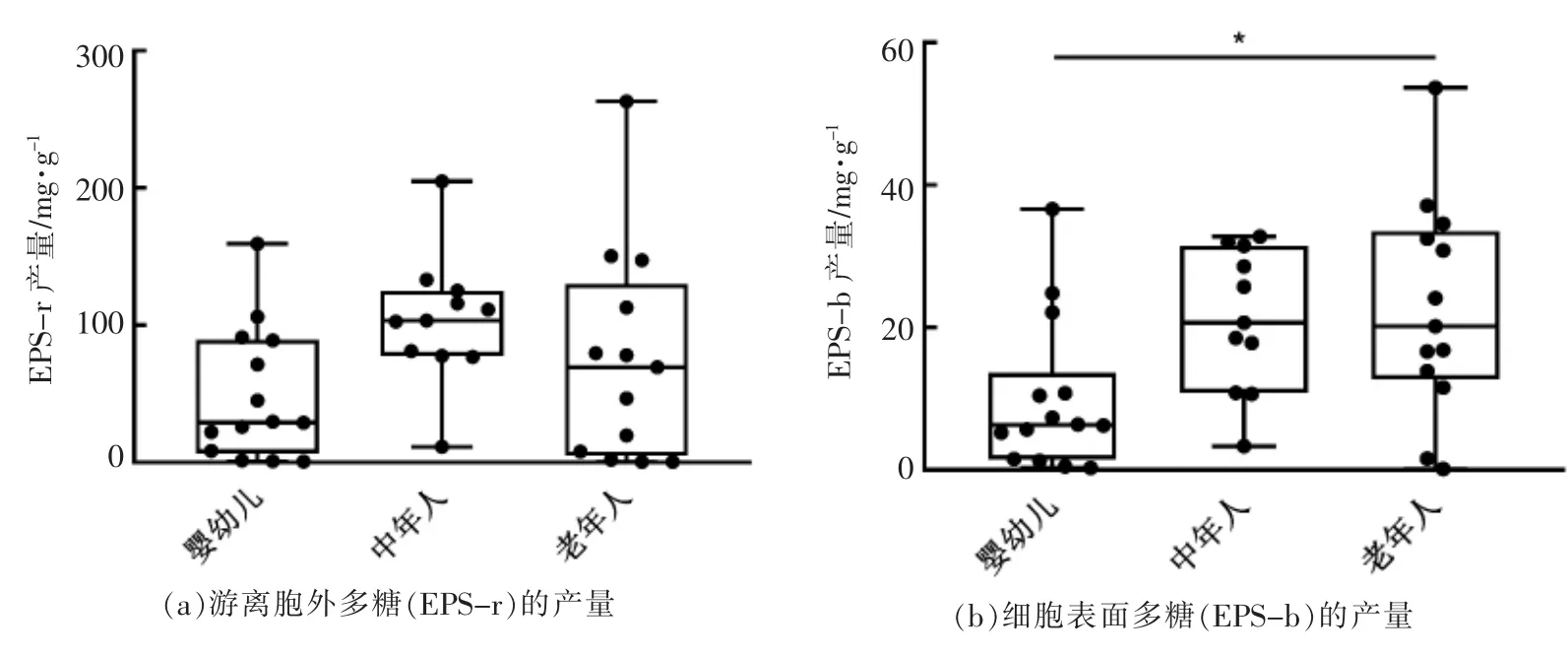
图4 不同年龄段宿主来源菌株的胞外多糖产量Fig.4 EPS production of strains from hosts with different ages
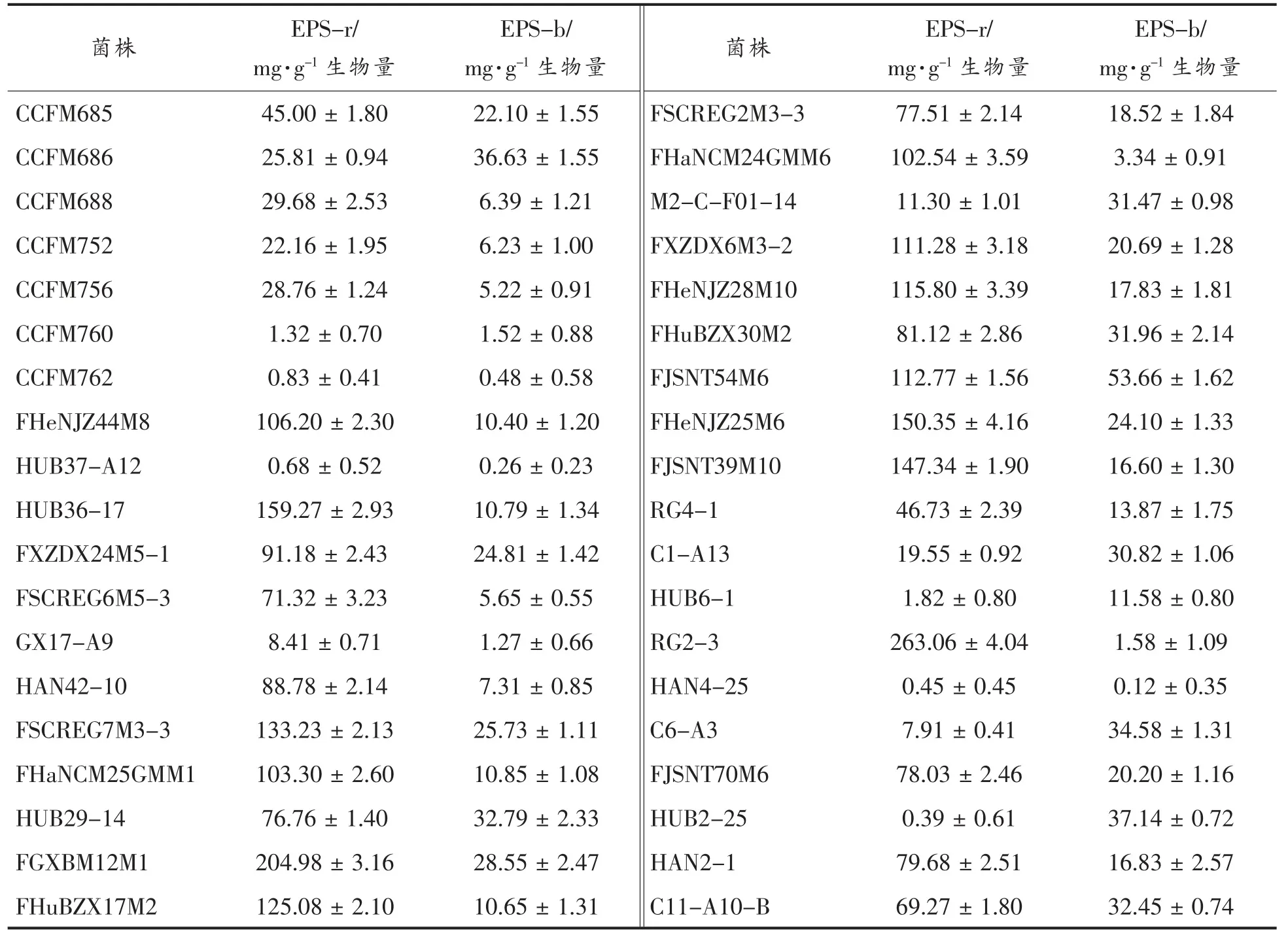
表2 不同长双歧杆菌菌株胞外多糖的产量Table 2 EPS productions of different strains
2.5 不同长双歧杆菌菌株pGT 基因测序及进化分析
引导型糖基转移酶是参与胞外多糖合成过程第1 步反应的酶。 不同菌株pGT 基因的C 端具有较高的保守性, 且在胞外多糖合成过程中参与脂质载体和糖基的识别[20],因此根据该基因C 端序列设计引物,对C 端的核酸序列测序,并对相应的氨基酸序列比对分析[20-21]。 除CCFM688 和M2-CF01-142 株菌外,其它36 株菌均通过该引物被检测到pGT 基因。 这可能是由于这2 株菌的pGT 基因序列与其它pGT 基因序列的相似性较低, 无法利用本试验设计的引物扩增所致[22]。 由图5 可知,扩增得到的长双歧杆菌pGT 的C 端具有较高的保守性。 根据系统进化树的分析结果(图5),这些序列可以分为3 组: 第1 组中主要为中年人和老年人源菌株的序列, 第2 组中主要为婴幼儿源菌株的序列, 而第3 组中主要为老年人源的菌株的序列。不同性别人群来源的菌株没有呈现聚类现象。
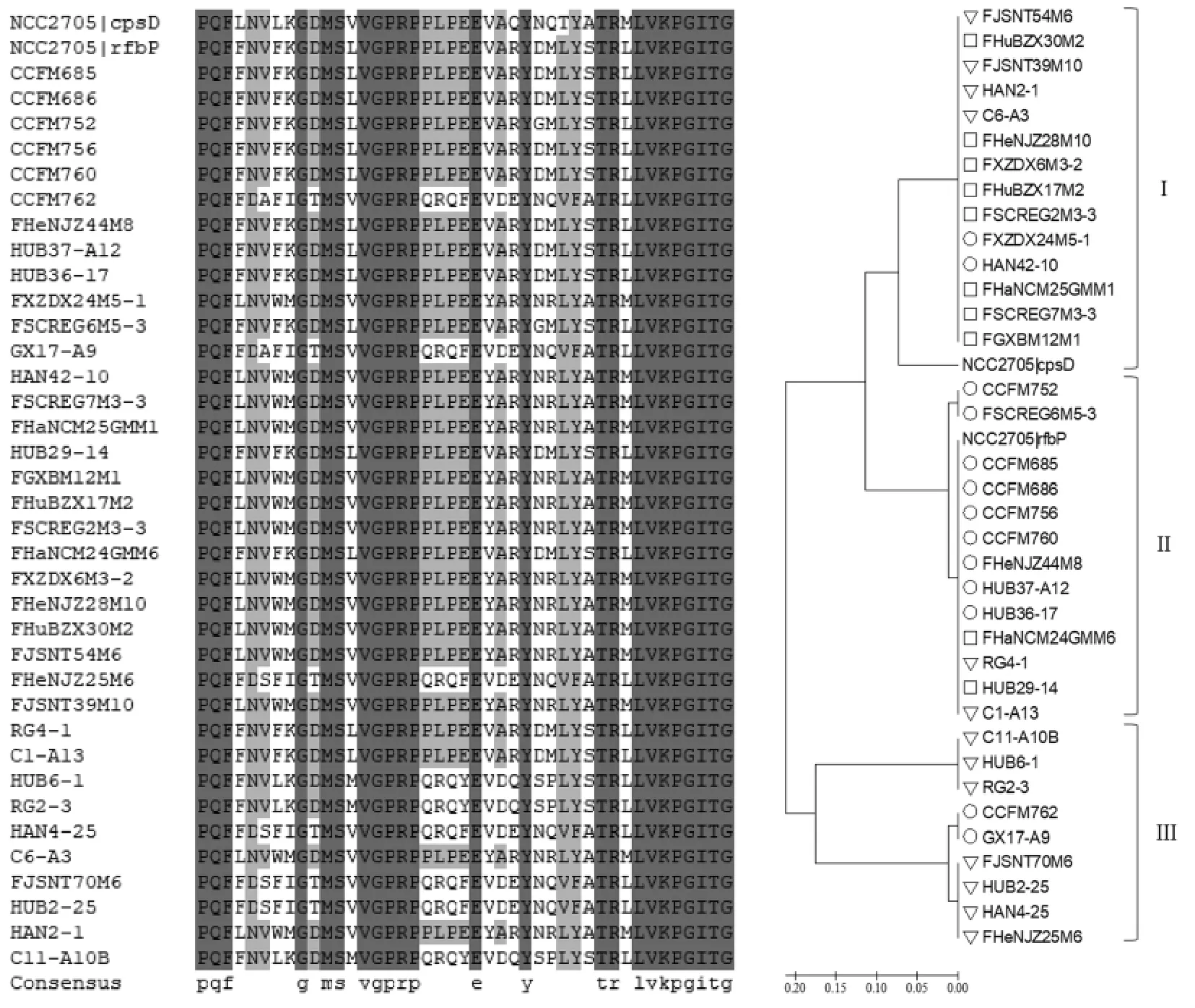
图5 不同菌株引导型糖基转移酶(pGT)C 端氨基酸序列的比对结果和进化树Fig.5 Alignment and phylogenetic tree of amino acid sequence of the carboxy terminus of priming glycosyltransferase of different strains
3 结论
从不同年龄志愿者的粪便样品中分离38 株长双歧杆菌,并分析各菌株的部分体外特性,包括生长曲线、碳水化合物利用试验、发酵上清液中短链脂肪酸组成、 胞外多糖产量和胞外多糖合成关键酶——pGT 酶的序列比对以及进化分析。 结果显示,在MRS 培养基中培养时,婴幼儿源长双歧杆菌的生长速率相对较慢, 在对数生长末期所能达到的活菌数也相对较低。 婴幼儿源菌株多数可利用甘露糖, 而中年人和老年人来源的菌株多数不能利用甘露糖。除了3 株老年人源的菌株,其它菌株都可利用低聚果糖、 低聚半乳糖和低聚异麦芽糖。 婴幼儿源菌株表面胞外多糖产量低于中年人和老年人来源的菌株, 与老年人源菌株具有显著性差异。系统进化分析结果表明,婴幼儿源菌株的pGT 酶C 端氨基酸序列和老人源菌株的pGT酶C 端氨基酸序列呈现部分聚类的趋势。
