低温胁迫对3种绿绒蒿幼苗叶绿素荧光特性的影响
2020-08-28侯祯丹严朋飞
黄 鸿,屈 燕,侯祯丹,严朋飞,刘 建
(西南林业大学园林园艺学院/国家林业局风景园林工程技术研究中心,云南 昆明 650224)
【研究意义】叶绿素是光合作用过程中用于捕获光能的主要色素,当PSⅡ反应中心叶绿素 P680 的一个电子被激发到高能量的激发态时,会开始电子传递过程[1]。叶绿素荧光分析技术已广泛应用于植物抗逆生理的研究,它具有快速、准确、灵敏、无损伤的特性,可以在保持植株完整的基础上测定出植株的光合作用过程的变化[2]。通常植物受到胁迫后的生理分析只能满足少量样品的研究,但由于受胁迫症状常是寒害的晚期表现,所以只单一的通过生理指标评价低温对植物的影响是较为困难的。而叶绿素在光合作用中产生的荧光信号对低温造成的细胞伤害较为敏感,同时包含着丰富信息,且不具有破坏性[3]。在低温下植物的抗寒性指标参数与叶绿素荧光参数的变化有一定相关性,通过测定和分析植物叶片的荧光参数的变化,可以更加快速准确地对植物抗寒性能力及受伤害的程度作出评价。【前人研究进展】一些研究发现低温会降低植物对光能的利用率,主要表现为光合速率、电子传递率、光合酶活性都会明显降低[4]。在低温胁迫下,植物会调节自身热耗散等一系列方式来保护光合机构免受破坏,PSⅡ反应中心可能会发生失活,这种失活分为可逆和不可逆2种[5]。植物在受到严重低温伤害时,可能会对PSⅡ反应中心产生不可逆失活,对光和机构产生严重伤害,PSⅡ反应中心是反映植物光化学反应的重要气管单位,PSⅡ的光化学效率参数能有效地反映植物光化学反映状况[6]。Fv/Fm是PSⅡ光系统中的最大光化学效率,可以直接、准确反映植物的光化学利用率,是评价植物是否受到光抑制的敏感指标[7]。在环境胁迫中,不同的植物的叶绿素荧光参数的变化也不相同,但大多数研究中,植物的PSⅡ的光化学量子产量Y(Ⅱ)、光化学淬灭和非光化学淬灭、电子传递速率等都会有上升或下降的变化[8]。低温通常是限制植物的生长和地域分配的关键因素[9]。低温胁迫不仅会影响植物生长形态,还会发生植物生理生化反应,伤害植物细胞结构,导致植物光合作用减弱[10],当温度降至引起冷害的临界点,植物光合酶活性、光合电子传递速率下降[11],光合作用会表现出明显的抑制作用[12]。王贺等[13]研究发现低温导致苹果最大光化学量子产量(Fv/Fm)降低,从而使叶片光合机制受阻。梁芳[14]等通过叶绿素荧光技术研究发现,菊花幼苗随低温胁迫程度的延长,光化学量子产量Y(Ⅱ)、电子传递速率ETR和Fv/Fm呈降低趋势。在一定胁迫程度下,植物会通过能量耗散途径等多种调节反应来保护光合机构收到破坏[15]。马博英等[16]通过研究植物的PSII实际光量子效率Y(II)、光合电子传递速率(ETR)和非光化学猝灭系数(NPQ)等叶绿素荧光参数,筛选出抗寒能力强的基因型草坪草。植物受严重低温胁迫时,可能对光合机构产生不可逆的破坏[17],影响光合电子传递和与暗反应有关的酶活性。陈曦等[18]发现0 ℃以下的低温胁迫对油松造成不可恢复的光合机构损伤。【本研究切入点】绿绒蒿常生长在海拔3000~5000 m高山草甸或者温差大、气温低、空气稀薄的高海拔地区的流石滩。其幼苗生长过程中必须忍受高山寒冷多变的气候特征。因此,绿绒蒿是研究高山植物在低温胁迫下的叶绿素荧光变化规律的理想材料。由于不同种类的绿绒蒿在生理适应性上存在明显差异。有学者通过对生长在海拔3000 m以上的总状绿绒蒿和全缘叶绿绒蒿的光合响应进行比较分析,发现在低温下全缘叶绿绒蒿的光合效率有明显下降,总状绿绒蒿对低温和强光有较强的适应能力,能在光抑制下迅速恢复,这可能是因为总状绿绒蒿叶片表面着生有细密绒刺能有效减轻强光的刺激。总状绿绒蒿在强光下会通过调节叶片比叶重、CO2导度来适应强光,较全缘叶绿绒蒿更适合在低海拔地区引种栽培[14]。【拟解决的关键问题】文章分别选取贝利叶绿绒蒿(Meconopsisbaileyi),多刺绿绒蒿(Meconopsishorridula)和高茎绿绒蒿(Meconopsissuperba)3种绿绒蒿幼苗,利用叶绿素荧光分析技术,对低温胁迫条件下3种绿绒蒿幼苗叶绿素荧光特性的变化进行研究,以此来探讨低温胁迫对不同亚属绿绒蒿幼苗叶绿素荧光特性的影响,为绿绒蒿属植物的引种栽培、以及高山植物抗寒因子研究提供借鉴。
1 材料与方法
1.1 试验材料及实验设计
试验材料为多刺绿绒蒿、高茎绿绒蒿、贝利叶绿绒蒿半年生实生苗。选取大小一致、长势良好的绿绒蒿幼苗置于人工气候箱中,先于20 ℃/16 ℃(昼/夜)适应培养2 d,每天光照12 h,然后以4 ℃/h降温至4 ℃进行低温胁迫处理,分别在0 h(CK)、低温处理12、24、48、72 h、恢复48 h时进行叶绿素荧光测定。
1.2 测定方法及指标
取2片暗适应20 min的绿绒蒿幼苗叶片,采用IMAGING-PAM(德国WALZ公司)叶绿素荧光测定仪进行叶绿素荧光参数测定。仪器基本参数:检测光强度0.1 μmol/m·s,光化光强度81 μmol/m·s,饱和脉冲强度2700 μmol/m·s,脉冲光0.8 s,每隔20 s打开1次。
1.3 数据处理
采用Excel 2003和SPSS 13.0统计分析软件进行数据分析,各参数数值为3次重复的平均值。
2 结果与分析
2.1 低温胁迫下幼苗最大荧光(Fm)变化
由图1可见,随着低温胁迫时间的延长,3种绿绒蒿的最大荧光(Fm)均有下降趋势,在72 h时Fm达到最低值,其中多刺绿绒蒿下降幅度最明显,比胁迫前下降了30 %。在胁迫24、48、72 h后,贝利叶绿绒蒿最大荧光Fm分别比0 h(CK)下降8.9 %、11.1 %、15.6 %;多刺绿绒蒿分别比0 h(CK)下降 4.6 %、13.9 %、30 %;高茎绿绒蒿分别比0 h(CK)下降2.1 %、11.2 %、16.1 %。经过常温48 h的恢复,3种幼苗的Fm均有上升趋势,但未恢复到0 h(CK)水平。
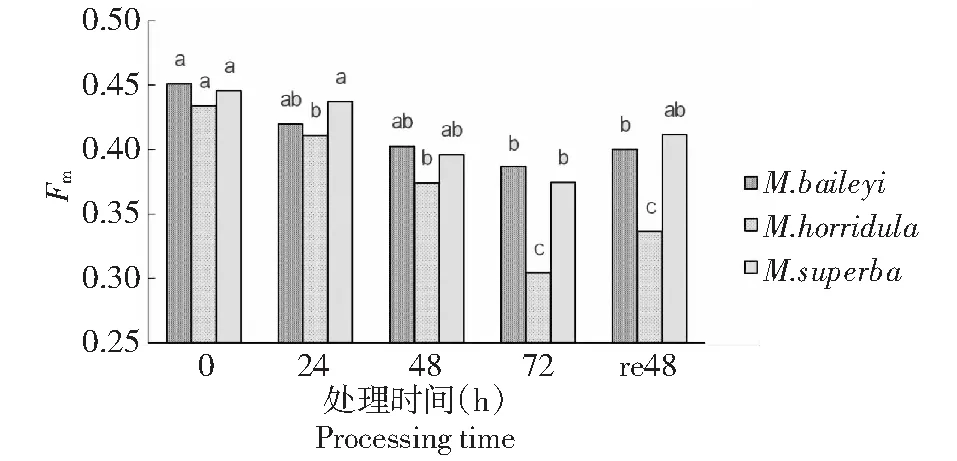
同一品种上不同小写字母表示差异达5 %显著水平。下同Different lowercase letters on the same breed indicate that the difference reaches a significant level of 5 %.The same as below
2.2 低温胁迫下幼苗的表现电子传递速率变化
由图2可见,低温胁迫处理期间,多刺和高茎绿绒蒿幼苗表观电子传递速率(ETR)随胁迫时间逐渐下降,贝利叶绿绒蒿ETR在24 h升高,继而开始下降;多刺绿绒蒿在48 h降至最低然后在72 h升高。贝利叶绿绒蒿、多刺绿绒蒿和高茎绿绒蒿的ETR在72 h下降,分别为0 h(CK)的88 %、45 %和89 %。经过48 h自然恢复,3种幼苗的ETR有所回升。

图2 低温胁迫下3种绿绒蒿幼苗的表现电子传递速率变化Fig.2 Changes of the expression electron transfer rate of three Meconopsis seedlings under low temperature stress
2.3 低温胁迫下幼苗的非光化学淬灭变化
由图3可见,低温胁迫处理期间,3种绿绒蒿幼苗的非光化学淬灭(NPQ)先上升后下降,均在恢复48 h后升高,贝利叶和多刺绿绒蒿达到最大值。自然恢复48 h后,高茎绿绒蒿的NPQ保持稳定,贝利叶绿绒蒿和多刺绿绒蒿有所上升,2种幼苗(NPQ)均显著高于0 h(CK)。

图3 低温胁迫下3种绿绒蒿幼苗的非光化学淬灭变化Fig.3 Changes of Non-photochemical quenching of three Meconopsis seedlings under low temperature stress
2.4 低温胁迫下幼苗PSⅡ实际光量子效率变化
由图4可见,低温胁迫处理期间,3种绿绒蒿幼苗的PSⅡ实际光量子效率Y(Ⅱ)逐渐降低,均在72 h达到最小值且显著小于0 h(CK,P<0.05),分别为0 h(CK)的84 %、66 %、82 %。经过48 h自然恢复,3种绿绒蒿的Y(Ⅱ)均有所回升,但仍然低于0 h(CK)。

图4 低温胁迫下3种绿绒蒿幼苗PSⅡ实际光量子效率变化Fig.4 Changes of PSⅡactual light quantum efficiency of three Meconopsis seedlings under low temperature stress
2.5 低温胁迫下幼苗的PSⅡ调节性能量耗散变化
由图5可见,低温胁迫处理期间,3种绿绒蒿幼苗的PSⅡ调节性能量耗散Y(NPQ)在胁迫24 h时升高,其中多刺绿绒蒿在24 h达到最大值。贝利叶绿绒蒿在24、48、72 h的Y(NPQ)与0 h(CK)差异不显著,经过48 h恢复显著高于0 h(CK)水平。多刺和高茎绿绒蒿在胁迫24 h后幼苗的PSⅡ调节性能量耗散Y(NPQ)均有不同幅度的下降。
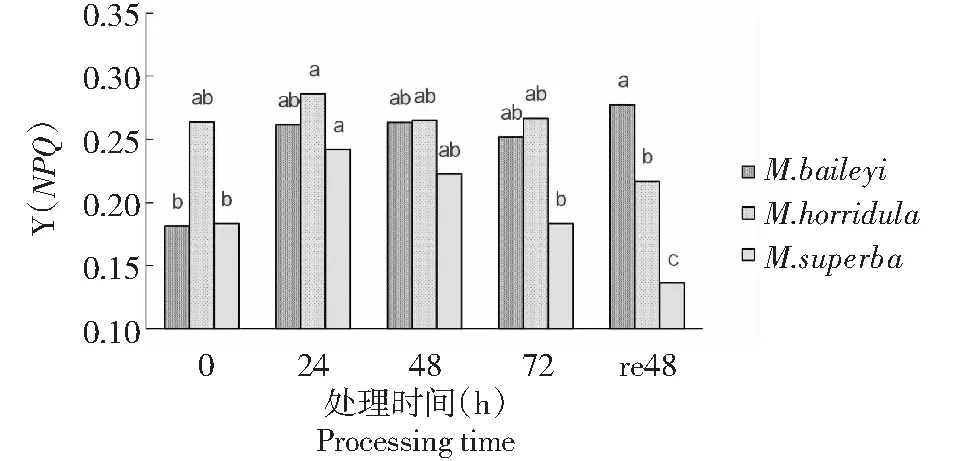
图5 低温胁迫下3种绿绒蒿幼苗的PSⅡ调节性能量耗散变化Fig.5 Changes of PSⅡregulatory in energy dissip of three Meconopsis seedlings under low temperature stress
2.6 低温胁迫下幼苗的PSⅡ非调节性能量耗散变化
由图6可见,低温胁迫处理期间,3种绿绒蒿幼苗的PSⅡ非调节性能量耗散Y(NO)逐渐上升,其中贝利叶和多刺绿绒蒿在48 h达到最大值,且显著大于0 h(CK,P<0.05),高茎绿绒蒿在72 h时达到最大值。经过48 h恢复3种绒蒿比较0 h(CK)均有回升,多刺绿绒蒿上升幅度最为明显。

图6 低温胁迫下3种绿绒蒿幼苗的PSⅡ非调节性能量耗散Fig.6 Changes of PSⅡregulatory energy dissipation of three Meconopsis seedlings under low temperature stress
2.7 低温胁迫下幼苗的PSⅡ潜在活性的变化
由图7可见,低温胁迫处理期间,3种绿绒蒿幼苗的PSⅡ潜在活性(Fv/F0)呈下降趋势,其中3种绿绒蒿在72 h达到最小值,且显著低于0 h(CK,P<0.05),说明低温影响了PSⅡ受体的电子传递。高茎绿绒蒿的PSⅡ潜在活性(Fv/F0)较低温胁迫前有下降但并无太大变化,说明受低温伤害相对较小。经过48 h恢复,3种绿绒蒿的PSⅡ潜在活性均有回升。
2.8 低温胁迫下8幼苗的PSⅡ最大光化学效率的变化
由图8可见,经低温胁迫72 h后,多刺绿绒蒿的最大光化学效率Fv/Fm的值显著低于0 h(CK) (P<0.05),胁迫72 h较胁迫前降低了36.2 %。在胁迫24 h贝利叶绿绒蒿和高茎绿绒蒿的最大光化学效率Fv/Fm分别0 h(CK)降低了13.2 %,15.6%,并随着胁迫时间延长缓慢下降。在恢复48 h后,3种绿绒蒿的Fv/Fm均有升高,但仍未恢复至0 h(CK)水平。这表明3种绿绒蒿幼苗都受到低温的影响并对低温具有较强的适应力,其中,多刺绿绒蒿叶片受低温胁迫的影响,在胁迫72 h时光能利用率最低,但在恢复48 h后,其最大光化学效率有显著升高,说明多刺绿绒蒿在72 h受到低温伤害,但又能在温度恢复之后通过自我调节来抵御伤害。

图 7 低温胁迫下3种绿绒蒿幼苗的PSⅡ潜在活性的变化Fig.7 Changes of PSⅡ potential activity of three Meconopsis seedlings under low temperature stress

图8 低温胁迫下3种绿绒蒿幼苗的PSⅡ最大光化学效率的变化Fig.8 Changes of maximum photochemical efficiency of PSⅡ of three Meconopsis seedlings under low temperature stress
3 讨 论
低温胁迫对植物荧光特性的影响与植物种类有关,不同种类具有一定差异。本试验研究结果表明,3种绿绒蒿在低温胁迫下,PSⅡ的电子传递速率受到影响都发生了不同程度的光抑制。在低温胁迫后3种绿绒蒿最大光化学效率(Fv/Fm)、光合电子传递量子效率ΦPSⅡ、光化学猝灭qP均显著降低,非光化学猝灭qN升高,电子传递速率(ETR)减慢,导致实际光化学量子产量(Y)下降,与低温胁迫对其他植物影响的类似研究结果相一致[19]。
3.1 低温胁迫下3种绿绒蒿的荧光特性
本研究中,随着低温胁迫时间的延长,3种绿绒蒿的最大荧光Fm均有下降趋势,在低温胁迫72 h时Fm达到最低值,说明PSⅡ的电子传递受阻并受到光抑制[20],其中多刺绿绒蒿下降幅度最明显。经过常温48 h的恢复,3种绿绒蒿幼苗的Fm均有上升趋势,仍未恢复到0 h时的水平。贝利叶绿绒蒿在不同胁迫时间电子传递速率ETR和光化学量子产量Y(Ⅱ)值较高,说明在低温下其光合电子传递速率较高[21]。高茎绿绒蒿的非光合效率和调节性热耗散量子产额的值随着胁迫时间延长有明显上升,表明高茎绿绒蒿相较贝利叶和多刺绿绒蒿来说对寒冷的敏感度不高,在低温下有较强的自我保护能力。低温胁迫后多刺绿绒蒿的电子传递速率ETR和光化学量子产量Y(Ⅱ)降幅大,自然恢复相对缓慢,常温恢复后Y(NO)仍和0 h时差异不显著,表明了多刺绿绒蒿在一定程度上对低温的适应力要弱一些。
3.2 低温胁迫对3种绿绒蒿光化学能量转换的影响
NPQ是植物对光合机构的一种自我保护机制,表示在PSⅡ电子传递中热耗散的程度[22-24]。3种绿绒蒿幼苗的非光化学淬灭系数NPQ先上升后下降,均在恢复48 h又上升并达到最大值。表明3种绿绒蒿都在低温胁迫下通过热耗散来减轻PSⅡ的损伤。胁迫72 h时3种绿绒蒿的NPQ值均有下降趋势,表明PSII热耗散的保护能力随着低温胁迫时间延长而有所减弱。温度恢复48 h后,高茎绿绒蒿的NPQ保持稳定,贝利叶绿绒蒿和多刺绿绒蒿有所上升。
非调节性能量耗散Y(NO)是光损伤的重要指标[25],Y(NO)值较高表示光化学能量转换和热耗散不能将植物所吸收的光完全消耗[26]。3种绿绒蒿幼苗的Y(NO)随着低温胁迫时间延长逐渐上升,在胁迫48 h时,均大于0 h时,说明此时3种绿绒蒿幼苗光和机构可能已经受到了损伤[27]。经过48 h恢复,3种绿绒蒿Y(NO)均高于0 h时水平。PSⅡ最大光化学效率Fv/Fm反映PSII光合反应下最大光能转换效率,能检测植物在自然环境下是否受到光抑制或其他胁迫。在本研究中,在低温胁迫下,3种绿绒蒿的最大光化学效率Fv/Fm的值均有所下降,即表明产生了不同程度光抑制,其中多刺绿绒蒿在胁迫72 h时下降幅度最大,光能利用率最低,其叶片受低温胁迫的影响较大。
4 结 论
对3种绿绒蒿幼苗的Fm、ETR、NPQ、Y(II)、Y(NPQ)、Y(NO)、Fv/F0和Fv/Fm等一系列荧光参数进行比较,持续低温胁迫下,贝利叶绿绒蒿ETR和Y(II)值随胁迫时间延长下降幅度较小且在温度恢复48 h后有明显上升,高茎绿绒蒿的NPQ和Y(NPQ)值在低温胁迫下上升得较快,相较于多刺有较好的低温自我保护。多刺绿绒蒿的电子传递速率ETR和光化学量子产量Y(Ⅱ)降幅大,自然恢复相对缓慢,常温恢复后Y(NO)和0 h时差异不显著,表明多刺绿绒蒿在一定程度上对低温的适应力要弱一些。
