早实核桃与晚实核桃光合特性比较研究
2020-08-28李秋煜丁欣欣贾莉芳张小军刘群龙
李秋煜,丁欣欣,贾莉芳,张小军,杨 笑,刘群龙
(山西农业大学 园艺学院,山西 太谷 030801)
【研究意义】核桃(JuglansregiaL.)为胡桃科(Juglandaceae)核桃属(Juglans)落叶乔木,是我国重要经济林树种之一[1]。根据核桃结实早晚和生物学特性的差异,将核桃分为早实和晚实两种类型[2]。光合作用是作物形成物质产量的基础,提高作物光合作用效能是增产的重要途径之一[3]。早实核桃和晚实核桃遗传特性的不同,导致两种类群核桃在生长特性和光合生理等方面存在较大差异[4-5]。【前人研究进展】目前,对同一核桃类群的不同品种间[6]、不同栽植密度和树形[7-8]以及不同肥水处理[9-10]光合特性影响的研究较多,关于早实核桃和晚实核桃类群的比较研究也多集中在生长特性、管理技术、树体调节[4]和叶绿素荧光[5]等方面,但在光合特性方面的研究较少[11]。【本研究切入点】以早实核桃‘农核1号’和晚实核桃‘晋龙2号’为试材,对其光合生理特性进行测定,并分析其净光合速率与主要环境因素间的关系。【拟解决的关键问题】比较早实核桃和晚实核桃光合特性的差异,为深入了解不同类群核桃生长机理、品种特性和促进核桃栽培技术发展提供参考依据。
1 材料与方法
1.1 试验材料
试验于2018年8月中旬在山西农业大学园艺学院试验站核桃种质资源圃进行。该站位于东经112°35',北纬37°25′ ,属温带季风气候,年降雨量450~500 mm,年平均温度9.8 ℃,无霜期175 d,年均日照时数2500~2600 h。选择早实核桃品种‘农核1号’和晚实核桃品种‘晋龙2号’,每个品种4株,树龄10年,株行距4 m×5 m,生长健壮,常规管理。
1.2 光合参数的测定
选择树体外围阳面生长一致的新梢中部复叶的顶叶下第2~4片功能叶,采用便携式Li-6400光合仪测定叶片净光合速率(Pn,μmol·m-2·s-1),从8:00-18:00,每小时测1次,每片叶读3次数,数据取平均值。同时记录蒸腾速率(Tr,mmol·m-2·s-1)、气孔导度(Gs,mmol·m-2·s-1)、胞间CO2浓度(Ci,μmol·mol-1)、光合有效辐射(PAR,μmol·m-2·s-1)、气温(Ta,℃)、空气相对湿度(RH, %)、空气CO2浓度(Ca,μmol·mol-1),并计算水分利用率(WUE,mmol·mol-1)和气孔限制值(Ls)等生理生态指标。
为比较环境因素作用大小,以净光合速率(Y)与光合有效辐射(X1)、空气温度(X2)、空气相对湿度(X3)和空气CO2浓度(X4)进行多元逐步回归,建立回归模型,并进行通径分析。
1.3 数据处理
采用Office 2019对数据进行整理,SPSS 23.0进行显著性、相关性和通径分析。
2 结果与分析
2.1 主要环境因素日变化规律
由图1可知,一天中光合有效辐射和空气温度均呈“单峰”曲线,光合有效辐射最高峰出现在12:00,峰值达2000.03 μmol·m-2·s-1;空气温度最高峰出现在14:00,峰值为34.00 ℃。空气相对湿度随空气温度的升高而降低,清晨达到最高,此后逐步减低,14:00达到谷值,呈V型变化趋势。空气CO2浓度在8:00-10:00明显降低,10:00之后变化较为平缓。
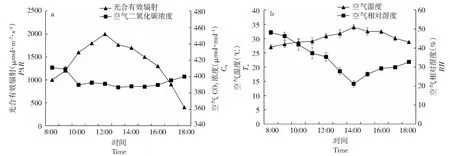
图1 环境因素的日变化过程Fig.1 Diurnal variations of the environmental factors
2.2 光合特征参数日变化规律比较
2.2.1 净光合速率和蒸腾速率日变化规律比较 由图2可知,‘晋龙2号’核桃叶片净光合速率日变化呈“双峰”曲线,‘农核1号’则呈单峰曲线,最高峰均出现在上午9:00,且‘农核1号’峰值较‘晋龙2号’显著提高19.58 %(P<0.05),日变化均值较‘晋龙2号’显著提高20.00 %(P<0.05);‘晋龙2号’于14:00降至低谷,出现光合“午休”现象。随空气温度升高,‘晋龙2号’和‘农核1号’叶片蒸腾速率日变化与净光合速率呈相同变化趋势,‘晋龙2号’蒸腾速率日变化均值显著高于‘农核1号’27.46 %(P<0.05)。‘晋龙2号’和‘农核1号’核桃叶片蒸腾速率均在14:00达到峰值,分别为4.44和3.56 mmol·m-2·s-1;且‘晋龙2号’较‘农核1号’显著提高24.82 %(P<0.05)。因此,‘农核1号’叶片净光合速率日变化显著高于‘晋龙2号’,蒸腾速率日变化与之相反。
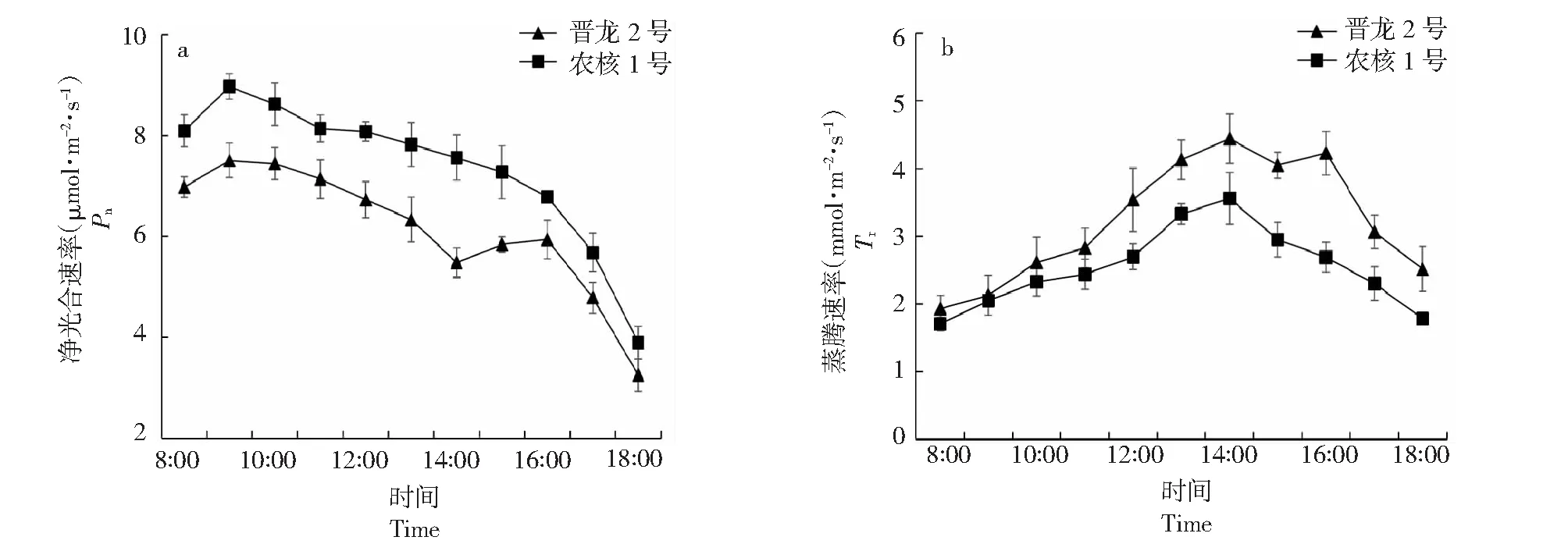
图2 ‘晋龙2号’和‘农核1号’核桃叶片净光合速率(a)、蒸腾速率(b)日变化Fig.2 Net photosynthetic rate (Pn) (a) and transpiration rate (Tr) (b) diurnal variation of ‘Jinlong 2’ and ‘Nonghe 1’
2.2.2 胞间CO2浓度和气孔导度日变化规律比较 由图3可知,‘晋龙2号’胞间CO2浓度日变化较平缓,‘农核1号’于12:00时后出现明显下降,17:00达到全天最低值213.52 μmol·mol-1。‘晋龙2号’胞间CO2浓度日变化均值较‘农核1号’显著提高17.78 %(P<0.05)。2品种叶片气孔导度日变化与净光合速率日变化趋势相同。‘晋龙2号’峰值出现在上午10:00,‘农核1号’出现在11:00,‘晋龙2号’叶片气孔导度日变化均值显著高于‘农核1号’(P<0.05)。因此,‘晋龙2号’与‘农核1号’叶片胞间CO2浓度和气孔导度变化规律存在差异,且‘晋龙2号’叶片胞间CO2浓度和气孔导度均显著高于‘农核1号’。
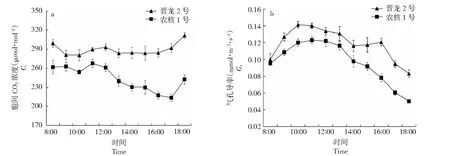
图3 ‘晋龙2号’和‘农核1号’核桃叶片胞间CO2浓度(a)和气孔导度(b)日变化Fig.3 Diurnal variation of CO2 concentration (Ci) (a) and stomatal conductance (Gs) (b) of ‘Jinlong 2’ and ‘Nonghe 1’
2.2.3 水分利用率和气孔限制值日变化规律比较 由图4可知,‘农核1号’和‘晋龙2号’叶片水分利用率变化规律一致,均在8:00-14:00逐渐下降,之后趋势平缓。‘农核1号’水分利用率日变化均值较‘晋龙2号’显著提高45.40 %(P<0.05)。2品种叶片气孔限制值均呈现“降-升-降”的变化规律,‘农核1号’谷值出现在上午11:00,‘晋龙2号’则出现在12:00,且‘农核1号’气孔限制值日变化均值显著高于‘晋龙2号’42.40 %(P<0.05)。因此,‘农核1号’叶片水分利用率和气孔限制值日变化均显著高于‘晋龙2号’。

图4 ‘晋龙2号’和‘农核1号’核桃叶片水分利用率(a)和气孔限制值(b)日变化Fig.4 Diurnal variation of leaf water use efficiency (WUE) (a) and stomatal limit value (Ls) (b) of ‘Jinlong 2’ and ‘Nonghe 1’
2.3 核桃光合参数与主要环境因素的相关性
由表1可知,‘晋龙2号’和‘农核1号’叶片净光合速率与光合有效辐射均呈显著正相关(r=0.631 0*,n=11;r=0.717 5*,n=11)。空气温度与空气相对湿度和空气CO2浓度呈极显著负相关(r=-0.913 8**,n=11,r=-0.797 4**,n=11)。空气相对湿度与空气CO2浓度呈极显著正相关(r=0.7728**,n=11)。
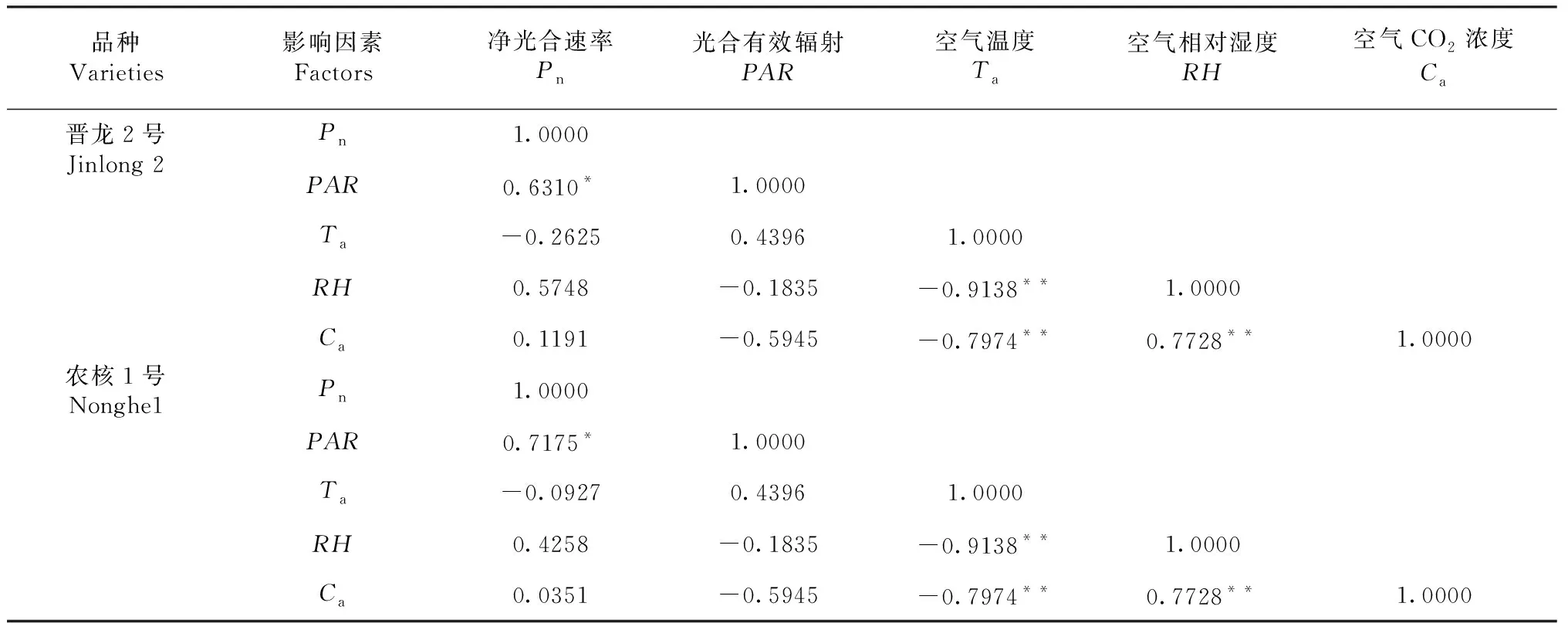
表1 净光合速率与主要环境因素的相关性
2.4 核桃光合参数与主要环境因素的通径分析
由表2可知,影响‘晋龙2号’叶片净光合速率的主要环境因素是光合有效辐射(X1)和空气相对湿度(X3)。通径分析(表3)表明,光合有效辐射和空气相对湿度对‘晋龙2号’叶片净光合速率的影响均表现为正效应。对各自变量的间接通径系数分析得出空气相对湿度通过光合有效辐射对净光合速率的影响较大。但由于光合有效辐射的直接通径系数最大,且与因变量的简单相关系数高达0.6310,同时光合有效辐射的决策系数(0.3809)>空气相对湿度(0.3108),所以环境因素对‘晋龙2号’叶片净光合速率影响的顺序为光合有效辐射>空气相对湿度。

表2 ‘晋龙2号’和‘农核1号’核桃叶片净光合速率日变化与主要环境因素的多元逐步回归分析
影响‘农核1号’净光合速率的主要环境因素是光合有效辐射(X1)和空气CO2浓度(X4)。通径分析表明(表3)光合有效辐射和空气CO2浓度对‘农核1号’净光合速率的影响均表现为正效应。对两个自变量的间接通径系数分析得出空气CO2浓度通过光合有效辐射对净光合速率的影响较大。但由于光合有效辐射在两者中的直接通径系数最大,并且与因变量的简单相关系数高达0.7175,同时光合有效辐射的决策系数(0.3345)>空气CO2浓度的决策系数(-0.4597),所以环境因素对‘农核1号’叶片净光合速率影响的顺序为光合有效辐射>空气CO2浓度。

表3 ‘晋龙2号’和‘农核1号’核桃叶片净光合速率日变化与主要环境因素的通径分析
3 讨 论
叶片光合日变化能够反映一天中植物生理代谢与物质积累的变化过程,同时也是分析植物生长和代谢受环境因素影响的重要手段[12],有助于了解植物光能利用情况。植物的光合作用是十分复杂的生理生化过程,在自然生长状态下,由于温度、光照、水分等外部环境以及品种特性、光合色素、叶龄叶位等自身发育状况,植物净光合速率日变化多呈现“单峰型”或“双峰型”曲线[13]。本试验中,早实核桃‘农核1号’叶片净光合速率日变化规律呈单峰曲线,晚实核桃‘晋龙2号’则为“双峰”曲线,这与王博[14]等、徐梦莎[15]等对早实核桃‘中嵩1号’和晚实核桃‘晋龙1号’、‘清香’在7月的研究结果相一致;而与张芸丽[11]对早实核桃品种‘香玲’和‘强特勒’在8月份净光合速率日变化的研究结果存在差异,这可能是地域环境及气候条件不同所导致。此外,早实核桃‘农核1号’的净光合速率日变化均值和最高峰值均显著高于晚实核桃‘晋龙2号’,说明早实核桃较晚实核桃具有更强的光合能力,这与全邵文等[5]对新疆地区早晚实核桃的研究结果相似。
根据Farquhar和Sharkey的研究[16],当净光合速率和胞间CO2浓度下降,且气孔限制值增大时,净光合速率的降低由气孔限制因素引起;若净光合速率下降,胞间CO2浓度上升,则由非气孔限制因素引起。本试验中,‘晋龙2号’在14:00时出现光合“午休”现象,期间净光合速率和气孔导度均下降,胞间CO2浓度上升,说明其光合“午休”是由非气孔限制因素造成的,这与辛洪河[17]等、王博[14]等的研究结果一致。
通过对净光合速率与主要环境因素进行相关性和通径分析,‘农核1号’和‘晋龙2号’叶片净光合速率均与光合有效辐射呈显著正相关,且光合有效辐射对两个核桃品种净光合速率的影响大于其它三个因素,是对净光合速率产生影响的主要决策因素,这与张俊佩[18]等研究结果相一致,因此生产上可采取加宽株行距、采用合理的树形以及进行适当的修剪来改善园内通风透光情况,从而提高产量。
4 结 论
早实核桃‘农核1号’与晚实核桃‘晋龙2号’的光合特性存在差异,早实核桃具有更强的光合能力。晚实核桃午间出现净光合速率下降是由高温、低湿和强光等非气孔限制因素所导致的。光合有效辐射是影响早实核桃和晚实核桃净光合速率的主要环境因素,生产中可采用较宽的株行距、合理修剪等措施促进通风透光,以提升叶片光合作用,从而促进树体生长和提高产量。
