不饱和脂肪酸对酿酒酵母生长及产香特性影响的研究进展
2020-08-22刘沛通郑晓卫段长青燕国梁
刘沛通,郑晓卫,段长青,陈 博,燕国梁,
(1.中粮营养健康研究院有限公司,老年营养食品研究北京市工程实验室,北京 102209;2.中国农业大学食品科学与营养工程学院,葡萄与葡萄酒研究中心,农业农村部葡萄酒加工重点实验室,北京 100083)
葡萄汁是一种非常复杂的基质,含有包括糖、有机酸等在内的数以千计的成分,它们的质量浓度水平从ng/L到g/L不等[1-2]。葡萄汁的成分受多种因素影响,如葡萄品种、产区、栽培管理和酿造工艺等[3-7]。葡萄汁的组成成分对葡萄酒的香气特征有决定性作用,各物质的微小变化甚至会直接影响酿酒酵母(Saccharomyces cerevisiae)的生长代谢,影响葡萄酒中挥发性组分的合成[8]。如柠檬酸可在发酵末期作为碳源被酵母代谢,其在葡萄汁中的含量影响酵母的生长[1];而向葡萄汁中适当添加铵盐、氨基酸等氮源则对酵母的生长繁殖有显著促进作用,同时增加酯类物质的合成[7,9]。近年来,葡萄汁中主要物质糖类(如葡萄糖和果糖)、羧酸类(如酒石酸和苹果酸)以及氨基酸(如丙氨酸和苯丙氨酸等)对酵母代谢影响的研究较多[1,7,9-11],但有关脂肪酸的研究仍不系统。
1 葡萄汁中的不饱和脂肪酸
葡萄果实中含有0.15%~0.24%(以湿质量计)的脂类化合物,以长链脂肪酸(C14~C18)含量最高,主要由亚油酸(C18:2)、油酸(C18:1)、亚麻酸(C18:3)、棕榈酸(C16:0)和硬脂酸(C18:0)组成[5,12]。发酵基质中的脂肪酸在酿造过程中起着关键作用。葡萄汁中约80%的脂肪酸在发酵中被酿酒酵母同化利用,为其生长代谢提供必需的营养成分,影响酵母的生理功能以及葡萄酒挥发性物质的合成,其中不饱和脂肪酸(unsaturated fatty acids,UFAs)的作用尤为重要[3,13-14]。
葡萄汁中UFAs的浓度及构成受多种因素影响,包括葡萄品种、产区、气候、栽培管理方式和酿造工艺等(表1)。研究表明,冷凉地区葡萄果实中的亚麻酸含量较高,而温暖地区葡萄果实中含有更多的油酸[15]。段亮亮[6]研究发现,转色结束时对葡萄果穗进行曝光处理可显著提高UFAs在总脂肪酸中的比例。实际生产中,常用的澄清工艺会显著减少葡萄汁中的UFAs含量,对葡萄酒品质有不良影响[16-17];与之相反,浸渍处理可提高葡萄汁中UFAs的含量,促进酵母的生长繁殖,改善葡萄酒香气品质[18]。

表1 葡萄汁中主要不饱和脂肪酸的质量浓度范围Table 1 Concentrations of main unsaturated fatty acids in grape must
2 UFAs对酿酒酵母生长代谢的影响
2.1 UFAs对酿酒酵母生长繁殖的影响
UFAs占葡萄汁中脂肪酸总量50%以上,是酿酒酵母生长代谢所必需的营养成分[16]。在厌氧条件下,酵母从发酵基质中摄取外源脂类物质(如麦角固醇或UFAs)来满足生长需求[23]。Redón等[24]研究发现,向发酵基质中添加脂肪酸(十二酸、棕榈油酸、亚麻酸)可显著增强酵母的活性,促进酵母繁殖,提高酵母细胞量。Beltran等[25]也发现酵母可通过吸收葡萄汁中的亚油酸来提高细胞活性。本实验室前期研究表明,高浓度的UFAs(油酸、亚油酸和亚麻酸)可上调酵母繁殖相关基因的表达量,诱导基因RAX2(编码N-糖基化蛋白)、SPS100(编码孢子壁成熟所需蛋白)、CLB1和CLB2(编码酵母细胞复制蛋白)表达,提高酵母的生长能力,显著提高酵母生物量[6]。UFAs还可在酵母细胞表面形成突起,促进酵母与环境之间的物质交换,增强酵母代谢活性,促进酵母的生长繁殖[26]。
发酵过程中,酿酒酵母不可避免地面临大量的胁迫因素,例如葡萄汁中极高的糖质量浓度(>200 g/L)导致的高渗透压胁迫,发酵过程中的营养饥饿、氧化胁迫,发酵后期的乙醇胁迫等,酵母对这些胁迫条件的耐受性直接影响其生长代谢,关系到葡萄酒的品质[27-30]。发酵基质中的UFAs可通过改善细胞抗逆性来提高酵母在胁迫环境中的生存能力。当基质中有多不饱和脂肪酸(polyunsaturated fatty acids,PUFAs)(如亚油酸或亚麻酸)存在时,酵母会将它们整合入细胞质膜,几乎取代质膜中原有的所有单不饱和脂肪酸(monounsaturated fatty acids,MUFAs),增强细胞膜流动性,提高酵母的抗逆能力[31-35]。如Landolfo等[36]通过向基质中添加UFAs(油酸和麦角固醇),增强酵母细胞对氧化胁迫的抵抗能力。Yazawa等[13]研究表明,产亚油酸和亚麻酸的菌株有较好的碱耐受性和较高的Li+、Na+、溶菌酶抵抗能力。Rodríguez-Vargas等[37]向酿酒酵母中转入Δ12去饱和酶促进亚油酸的合成,增强了细胞抗冻性。细胞膜中UFAs含量的增加也可提高酵母的耐热性[38],而UFAs(油酸)也可增加酵母的乙醇耐受性[39]。在葡萄酒发酵过程中,外源UFAs的供给可为酿酒酵母抵抗逆境、维持正常生长繁殖提供保障。
2.2 UFAs对酿酒酵母中脂肪酸合成与吸收的影响
酿酒酵母胞内的脂肪酸一部分从发酵基质中吸收同化,另一部分由酵母自身合成[31]。发酵基质中的UFAs会对酵母脂肪酸的吸收与合成产生重要影响。
酿酒酵母合成的饱和脂肪酸和UFAs主要是16碳及18碳的脂肪酸,包括棕榈酸、硬脂酸、油酸和棕榈油酸(C16:1)[24,31,40]。脂肪酸的合成及吸收如图1所示。糖代谢产生的乙酰CoA在乙酰CoA羧化酶(acetyl-CoA carboxylase,ACC)1的催化下生成丙二酰CoA,随后在脂肪酸合成酶(fatty acid synthase,FAS)1、FAS2和脂酰CoA延长酶(acyl elongase,ELO)1的作用下合成脂酰CoA,多次反应后合成棕榈酰CoA(C16:0-CoA)和硬脂酰CoA(C18:0-CoA)。在Δ9-脂肪酸去饱和酶OLE1催化下生成MUFAs[41]。由于缺少Δ12-脂肪酸去饱和酶和ω-3脂肪酸去饱和酶,酿酒酵母不能合成PUFAs,如亚油酸和亚麻酸[42-43]。
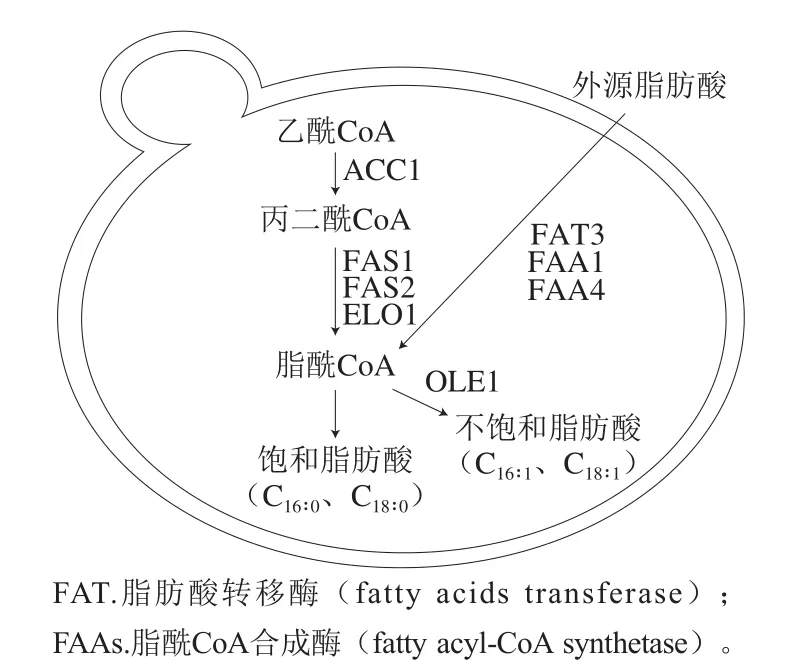
图1 酿酒酵母脂肪酸的合成及吸收[31]Fig.1 Synthesis and transportation of fatty acids in Saccharomyces cerevisiae[31]
厌氧条件下,酵母自身合成脂肪酸不足,为了维持正常的生长代谢,酵母必须从发酵基质中摄入脂肪酸,其中UFAs被优先吸收[23]。在FAT3及FAAs的作用下,发酵基质中的UFAs进入胞内,调节脂肪酸的合成代谢。酵母胞内的外源UFAs通过下调脂肪酸合成基因ACC1、FAS1和FAS2的表达量来抑制脂肪酸的内源合成[44-46]。另一方面,UFAs下调脂肪酸去饱和基因OLE1的表达[47],减少内源MUFAs的合成[45-46]。发酵前期,酵母细胞大量吸收外源UFAs,其中40%整合入细胞膜内,替代膜中原有的MUFAs[48],酵母细胞膜中的高UFAs含量可增强细胞膜的流动性,保持细胞膜完整性,保证酿酒酵母的正常生长代谢[35,49]。酵母对外源脂肪酸的吸收不仅可直接调控内源脂肪酸的合成,也会影响其自身的代谢活动。
2.3 UFAs对酿酒酵母中心碳代谢的影响
酵母的中心碳代谢(central carbon metabolism,CCM)包括糖酵解路径、戊糖磷酸路径和三羧酸循环[50-52]。发酵基质中的UFAs可通过调控CCM来影响甘油、乙醇、乙酸等代谢产物及乙酰CoA等关键代谢中间物质的合成,对葡萄酒品质产生重要影响。如图2所示,糖酵解过程中产生的中间产物磷酸二羟丙酮可经转化生成甘油-3-磷酸,去磷酸化后生成甘油。葡萄酒中的甘油质量浓度通常为5~14 g/L,为葡萄酒带来甘甜味和圆润感[53]。甘油是酵母细胞调节氧化还原平衡和渗透压的重要物质,为细胞供能(提供ATP),其产生与酵母对外界逆境的响应有关[53-55]。通常认为,甘油为磷脂的合成提供前体物质[56],发酵基质中的UFAs含量会影响酵母胞内磷脂浓度,进而通过影响磷脂的合成来调节甘油的代谢。
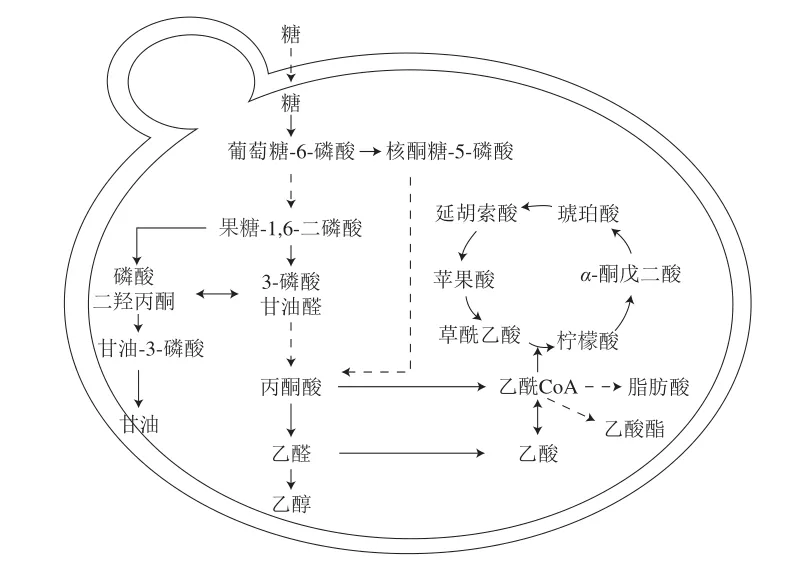
图2 酿酒酵母的中心碳代谢[50-52]Fig.2 Central carbon metabolism in Saccharomyces cerevisiae[50-52]
丙酮酸是CCM中的关键代谢物,由糖酵解路径产生[57]。在有氧条件下,酵母进行呼吸作用,丙酮酸在丙酮酸脱氢酶的作用下转化为乙酰CoA;而无氧条件下的丙酮酸参与发酵反应生成乙醇[56]。乙酰CoA有多条代谢路径,可参与三羧酸循环,也可生成乙酸,部分乙酰CoA进入脂肪酸合成路径,还有一些与高级醇发生酯化反应生成乙酸酯(图2)。发酵基质中的脂类物质会对乙酰CoA的代谢流向产生重要影响。Rollero等[58]研究发现,酵母可吸收发酵基质中的甾醇,减少乙酰CoA在脂类合成路径上的流量,通过调控胞内乙酰CoA库来影响三羧酸循环。三羧酸循环中的柠檬酸、琥珀酸和苹果酸是葡萄酒中重要的非挥发性酸。酵母在发酵初期产生柠檬酸,随着发酵的进行,基质中碳源含量下降,发酵末期柠檬酸作为碳源被酵母代谢[59]。UFAs对葡萄酒中柠檬酸的含量无显著影响[23,60]。琥珀酸与苹果酸的合成受环境因素调控,如pH值、糖浓度和氮源浓度等[53]。UFAs可通过影响酵母对发酵基质中糖、氮源的代谢[58,61-62],调控琥珀酸与苹果酸的生成。酵母胞内乙酰CoA流向的变化也会影响乙酸的合成。葡萄酒中的乙酸质量浓度一般为0.2~0.7 g/L,高于1.1 g/L时会给酒带来难闻的酸味[53]。乙酸的合成需要NAD+的参与,添加UFAs可增加酵母细胞量,提高胞内NADH水平,减少NAD+的浓度,抑制乙酸合成[23,63]。但也有研究表明,UFAs对乙酸浓度的影响随酵母菌株的不同而变化,添加油酸和亚麻酸能显著抑制酿酒酵母X5的乙酸合成,而亚油酸的添加能提高酿酒酵母EC1118发酵酒中的乙酸浓度,UFAs浓度及种类对乙酸合成的影响具有菌株特异性[22]。
乙醇是酵母发酵的重要代谢物,是葡萄酒中的关键物质。改变发酵基质中碳源、发酵温度,调节酵母胞内NADH水平等方式可影响葡萄酒中乙醇的含量[64-65]。Pham等[66]向发酵基质中添加脂肪酸和固醇,显著促进了乙醇的合成。Pinu等[22]却发现UFAs(油酸、亚麻酸)的添加降低了葡萄酒中的乙醇浓度。但也有研究表明,外界环境对乙醇的调控具有菌株特异性[67]。随着发酵的进行,葡萄酒中的乙醇含量升高,对酵母的胁迫也逐渐增大,近年来,如何提高菌株对乙醇胁迫的抵抗能力成为研究热点[68]。外源UFAs对酵母细胞膜的改善,能提高菌株的乙醇耐受性,有利于发酵的进行[39,69]。
2.4 UFAs对酿酒酵母发酵香气物质累积的影响
UFAs可以调控酿酒酵母香气物质的积累,影响葡萄酒的感官品质。主要香气物质的阈值及香气描述见表2。
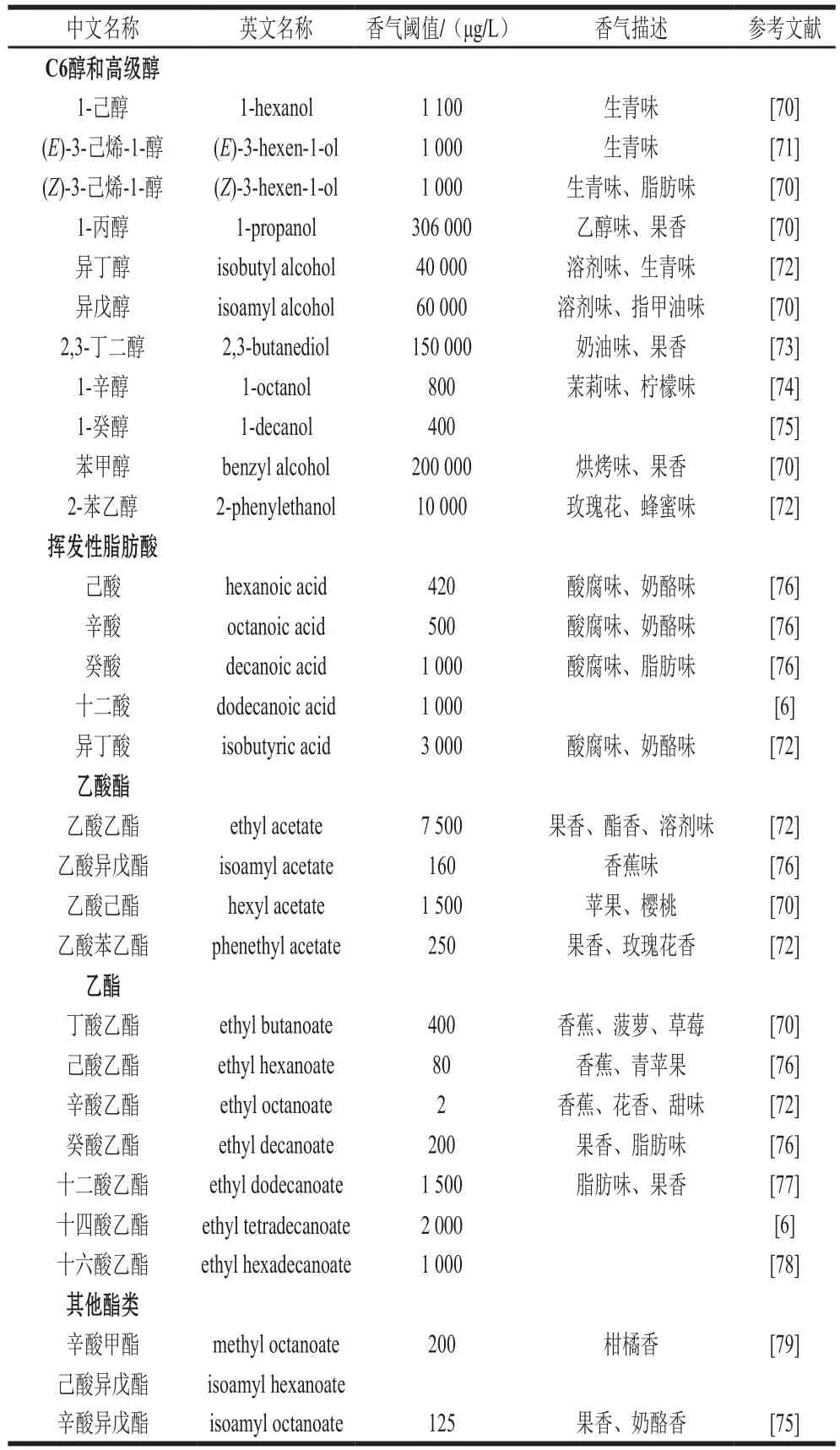
表2 葡萄酒中主要香气物质及其阈值和描述Table 2 Thresholds and aroma characteristics of main volatile compounds in wine
C6醇可由亚油酸和α-亚麻酸通过脂氧合酶/脂氢过氧化物裂解酶催化生成,为葡萄酒带来青草味,对葡萄酒香气有不良影响[3,80]。向发酵基质中添加UFAs(油酸、亚油酸和α-亚麻酸)会显著提高葡萄酒中C6醇的浓度[60]。
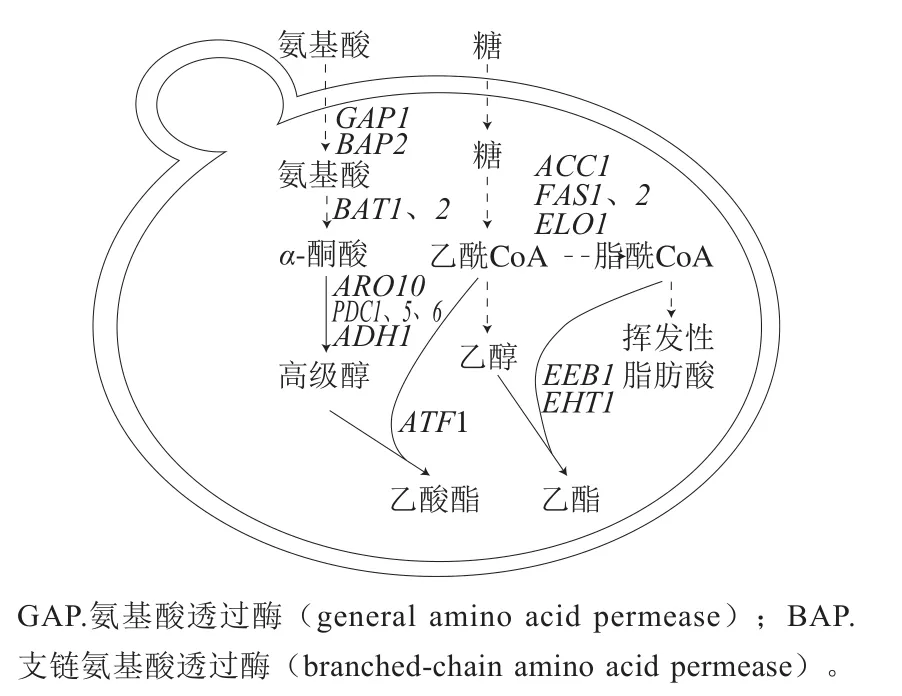
图3 酿酒酵母挥发性组分(高级醇、挥发性脂肪酸、乙酸酯和乙酯)的合成[53]Fig.3 Synthesis of volatile compounds (higher alcohols, volatile fatty acids, acetate esters and ethyl esters) by Saccharomyces cerevisiae[53]
高级醇是葡萄酒中重要的发酵香气物质,适宜质量浓度(<300 mg/L)的高级醇可增加葡萄酒香气复杂性,但质量浓度过高(>400 mg/L)时会给葡萄酒带来溶剂味和醇味[53]。高级醇主要由氨基酸通过Ehrlich途径代谢生成[9]。如图3所示,发酵基质中的氨基酸由GAP1、BAP2转运入酵母细胞内。胞内的氨基酸在转氨酶(BAT1和BAT2编码)的作用下转氨生成酮酸,再由脱羧酶(PDC1、PDC5、PDC6和ARO10编码)催化生成醛,最后经过多种醇脱氢酶(alcohol dehydrogenase,ADHs)的催化生成高级醇[9]。有研究表明,外源添加UFAs可通过调节膜蛋白活性及相关基因表达量,显著提高葡萄酒中的高级醇含量[23,81-82]。氨基酸透过酶的表达活性受细胞膜磷脂双分子层调控,细胞膜上UFAs(油酸、亚油酸、甾醇)比例的上升可提高膜连接的氨基酸转运蛋白的活性,促进胞外氨基酸向胞内转运[58,83-84]。段亮亮[6]研究发现,UFAs的添加可上调BAP基因表达量,利于酵母吸收发酵基质中的支链氨基酸,从而提高葡萄酒里异丁醇的含量,改善葡萄酒的品质[85]。Rollero等[62]研究发现氮源(氨基酸和铵盐)的代谢与发酵基质中植物甾醇的含量之间存在正相关关系,当甾醇含量增加,异丁醇和异戊醇的产量升高。本课题组前期研究也发现,UFAs的浓度对氨基酸代谢和高级醇的合成有显著影响,葡萄汁中低质量浓度(12 mg/L)的UFAs可促进支链氨基酸代谢,提高高级醇等物质的合成;但高质量浓度(120 mg/L)的UFAs会下调GAP1、ADH1等基因的表达量,减弱支链氨基酸对酵母生长的促进程度并抑制酵母合成高级醇及其酯类物质的能力[86]。
挥发性脂肪酸为葡萄酒贡献果香、奶酪味和脂肪味,低浓度的脂肪酸可改善葡萄酒的香气复杂性,但含量过高时会给葡萄酒带来不良风味[53]。葡萄酒中的挥发性脂肪酸主要通过酵母的同化作用合成(图3)。向发酵基质中添加UFAs(如棕榈油酸、α-亚麻酸)可造成脂肪酸合成基因(ACC1、FAS1、FAS2、ELO1)的表达量下调[87],抑制中短链脂酰-CoA的合成,降低葡萄酒中挥发性脂肪酸浓度[24,31,46,82]。同时,UFAs对酵母胞内乙酰CoA库的调节也会影响挥发性脂肪酸的合成,酵母从发酵基质中吸收的甾醇产生底物反馈抑制调节,能减少乙酰CoA在脂类合成路径上的流量,降低挥发性脂肪酸的含量[58]。
酯类物质包括乙酸酯和乙酯,为葡萄酒提供果香和花香等香气特征[88]。目前的研究认为,酯类物质的合成是酵母胞内长链脂肪酸的脱毒过程,同时也是胞内平衡乙酰CoA/CoA比率的代谢过程[89-90]。乙酸酯由乙酰CoA和高级醇在醇酰基转移酶(acetyltransferase,ATF)1的催化下缩合而成(图3),主要包括乙酸异丁酯、乙酸异戊酯、2-乙酸苯乙酯等[53]。敲除酿酒酵母的醇酰基转移酶基因ATF1能显著降低乙酸乙酯、乙酸异戊酯等乙酸酯的含量,而过表达该基因时,葡萄酒中的乙酸酯类物质浓度增加10~200 倍不等[88]。发酵基质中的UFAs可降低乙酸酯合成关键基因ATF1的转录和活性,抑制高级醇和乙酰CoA的酰化反应,减少乙酸酯的含量[21,46,89,91]。乙酯由脂酰CoA和乙醇在乙酯合成酶(ethyl ester biosynthesis,Eeb1p)和醇己酰基转移酶(ethanol hexanoyl transferase,Eht1p)作用下缩合而成(图3),常见乙酯包括丁酸乙酯、己酸乙酯、辛酸乙酯、癸酸乙酯和十二酸乙酯[88]。EHT1编码乙醇己酰转移酶,催化乙醇和己酰CoA缩合生成己酸乙酯,在中链脂肪酸乙酯合成中的作用较小[92]。EEB1编码乙醇酰基转移酶,是乙酯类物质合成路径中的关键基因。EEB1也参与短链酯类的水解反应,与脂类代谢和脱毒有关[88]。已有研究认为,乙酯的合成由底物浓度决定,与基因表达水平无关[93-94]。过表达EEB1和EHT1并不能显著提高葡萄酒中乙酯类物质的浓度[93]。但也有研究表明,底物浓度与乙酯含量之间无对应关系,EEB1等关键基因是调控乙酯合成的关键因素[6,95]。向发酵基质中复合添加UFAs(油酸、亚油酸和α-亚麻酸)能显著提高葡萄酒中己酸乙酯、辛酸乙酯和癸酸乙酯的含量[60],但也有研究发现,提高UFAs的含量会减少短链脂酰-CoA的生成,抑制酵母合成乙酯[21,93]。目前关于乙酯合成的研究并未得到一致的结果,UFAs可通过综合调控底物水平(通过乙酰CoA库调控乙酯前体物脂酰CoA的浓度)和基因水平(影响EEB1及EHT1的表达量)来影响乙酯类物质的合成,但具体机制仍需进一步研究。
除了浓度之外,发酵基质中UFAs的种类也影响酿酒酵母挥发性组分的合成。当发酵基质中油酸和麦角固醇的含量升高时,酵母可产生较多的高级醇和乙酸酯[81],但增加亚油酸的浓度反而抑制酯类物质的合成[96];Varela等[23]向发酵基质中添加吐温-80(含体积分数70%油酸、30%硬脂酸与棕榈酸)却提高了葡萄酒中酯类、高级醇和挥发性脂肪酸的含量。有研究表明,PUFAs(如亚油酸和α-亚麻酸)对细胞膜流动性的改善大于MUFAs(如油酸),更有利于酵母细胞在胁迫环境中生存[82]。这可能是因为PUFAs提高了酵母细胞膜的流动性,在胁迫条件下,有利于保持ATP酶及氨基酸透过酶等膜连接蛋白的活性[83,97]。不同种类UFAs对酵母生理特性的影响不同,这直接导致在发酵过程中酵母的不同代谢响应,影响挥发性组分的合成,造成葡萄酒中不同的香气特性。为了进一步探讨UFAs的种类对酵母产香能力的影响,本课题组分别研究对比了油酸、亚油酸和亚麻酸对酿酒酵母生长代谢的调控[98],结果发现,UFAs的种类会影响酵母的生长代谢:添加亚油酸和油酸更有利于酵母的生长和发酵,可通过上调FAT3、ACC1、FAS1、FAS2和ATF1的表达量,促进中链脂肪酸(辛酸和癸酸)和乙酸酯(乙酸异戊酯)的合成;而添加α-亚麻酸可上调BAT1、BAT2、PDC1、PDC5和PDC6的表达量,显著提高了C6醇(1-己醇)和高级醇(异丁醇和2,3-丁二醇)的含量。UFAs的种类是影响酵母生长、发酵特性及挥发性组分合成的重要因素。
3 结 语
UFAs对葡萄酒酿造过程中酵母的生长代谢、香气物质的合成有重要影响,不同浓度、种类的UFAs会造成酵母不同的发酵表现,给葡萄酒带来不同的香气特性。综合前人研究,UFAs调控酿酒酵母产香特性的机制有以下几方面:1)UFAs通过提高酿酒酵母的代谢活力促进酵母的生长繁殖;2)UFAs通过促进氨基酸转运、上调高级醇合成基因增加高级醇的合成,但该促进作用与UFAs的浓度等因素有关;3)UFAs可影响中链脂肪酸合成基因的表达量,调控中链脂肪酸的合成;4)UFAs通过下调醇酰基转移酶基因(ATF1)的表达量抑制乙酸酯的合成,但从底物水平和基因水平综合调控影响乙酯类物质的合成,且该调控根据发酵菌株、发酵基质等条件的不同而变化;5)UFAs的种类会影响其对挥发性组分合成的调控,葡萄汁中不同种类的UFAs会给葡萄酒带来不同的香气特性。
因此,在实际生产中,可针对性地选择不同的葡萄栽培方式、葡萄酒酿造工艺,根据葡萄汁中UFAs的不同结构有目的性地酿造具有不同香气特性的葡萄酒。如通过采取田间曝光处理,提高葡萄汁中油酸的浓度,从而增加葡萄酒中乙酸酯的含量,赋予葡萄酒更多的花香特性等。现阶段关于UFAs调控酿酒酵母生长和产香特性机制的研究仍不全面,UFAs对乙酯类物质合成的调控机制仍需深入探究。同时,UFAs对酿酒酵母生长代谢的调控随发酵基质、发酵菌株等的变化而不同,后续可通过应用现代多组学技术(微生物组学、基因组学、蛋白组学、代谢组学等)揭示UFAs的调控机理,为葡萄种植过程中栽培处理的合理实施及发酵过程中发酵工艺的合理选择、进一步提升酵母的发酵性能和葡萄酒的品质提供理论依据。
