反式肉桂醛对副溶血性弧菌毒力因子的抑制作用
2020-08-22邓海潮庸琪瑶赵鹏瑜邓瑞莎夏效东
王 硕,邓海潮,郭 都,庸琪瑶,赵鹏瑜,邓瑞莎,石 超,夏效东
(西北农林科技大学食品科学与工程学院,陕西 杨凌 712100)
副溶血性弧菌(Vibrio parahaemolyticus)是弧菌科弧菌属的一种无芽孢、无荚膜、有鞭毛、能运动的短杆或弧杆状的革兰氏阴性细菌[1]。副溶血性弧菌是一种常见的嗜盐菌[2],可在含盐量为0.5%~8.0%的环境中正常生长[3],广泛存在于海水及海产品中,其在夏季(7~9月份)海产品中的检出率高达50%以上[4],其中虾类、鱼类和贝类受副溶血性弧菌污染尤为严重[5]。食用被副溶血性弧菌污染的生的或未煮熟的海鲜后,可能会出现腹泻、头痛、呕吐、恶心、腹痛、低热等症状[6],重症患者还可能脱水、休克、昏迷甚至死亡[7]。有报道称,副溶血性弧菌已逐渐成为引起我国食源性食物中毒的首要致病菌,严重威胁着我国公众的健康安全[8]。并且在全球范围内,副溶血性弧菌也是导致肠胃炎等疾病频发的主要致病菌[9-10]。
副溶血性弧菌的致病性与其毒力因子密不可分。细菌的运动性在其侵染宿主细胞的过程中起着至关重要的作用[11],副溶血性弧菌能够通过口腔进入人体胃肠道,通过黏附肠上皮细胞、穿越肠道屏障,进而侵入血管破坏宿主体内环境,引发疾病[12]。因此,黏附和侵入人体肠上皮细胞是副溶血性弧菌引发感染的必经途径。并且副溶血性弧菌易在食品加工机械表面和食品包装上形成生物被膜,生物被膜对细菌起着保护作用,这使得细菌对环境的耐受能力大大增强[13-14]。副溶血性弧菌的这些毒力因子使得其具有感染率高、难以被清除的特点,因此,寻求一种安全有效的副溶血性弧菌控制方法对保障公众安全具有重大意义。
目前,控制副溶血性弧菌感染主要依赖于抗生素类物质[15]。但近些年来,抗生素的滥用使诸多问题也随之涌现,主要表现为细菌耐药菌株增加和耐药性增强[16],这给使用抗生素治疗疾病的方式带来了巨大的挑战。近年来,植物源活性物质因具有天然、安全和营养等优点而受到广泛关注[17],其抑菌功效也成为许多学者的研究热点[18]。肉桂醛是肉桂精油的主要活性成分,主要存在于肉桂树皮中,在自然界中的天然存在形式为反式肉桂醛(C9H8O)[19]。反式肉桂醛已被美国食品药品监督管理局列为公认安全的食品成分。研究表明,反式肉桂醛具有多种生物活性,可在抗癌、抗炎、治疗糖尿病等方面发挥功效[19],且对阪崎克罗诺肠杆菌、沙门氏菌、大肠杆菌O157:H7等有良好的抑制效果[20]。然而,反式肉桂醛对副溶血性弧菌毒力因子的抑制作用及其机制鲜有研究。
基于此,本研究将探究反式肉桂醛对副溶血性弧菌毒力因子的抑制作用。首先,通过测定反式肉桂醛对副溶血性弧菌的最小抑菌浓度(minimum inhibitory concentration,MIC),以评价其抑菌效果,并确定亚抑制浓度;在此基础上,探究在亚抑制浓度下,反式肉桂醛对菌体泳动运动性、生物被膜形成能力和黏附及侵入Caco-2细胞能力的影响,并探讨反式肉桂醛对副溶血性弧菌多种毒力因子的抑制作用,旨在为反式肉桂醛进一步应用于食品工业中控制食源性疾病提供理论依据。
1 材料与方法
1.1 菌株、材料与试剂
副溶血性弧菌(Vibrio parahaemolyticus)标准菌株ATCC 17802、ATCC 33847购于美国模式培养物集存库(American type culture collection,ATCC);副溶血性弧菌分离菌株202、209、210、253、258、259、260、261由香港理工大学食物安全及科技研究中心分离自鲜虾。实验所用的Caco-2细胞(人克隆结肠腺癌上皮细胞)购于武汉大学细胞保藏中心。
反式肉桂醛(纯度≥99%) 美国Sigma公司;质量分数3%氯化钠胰蛋白胨大豆琼脂(3% NaCl tryptone soya agar,3% NaCl TSA)、3%氯化钠胰蛋白胨大豆肉汤(3% NaCl tryptone soya broth,3% NaCl TSB)北京陆桥技术有限公司;DMEM细胞培养液 美国Gibco公司;胎牛血清(fetal bovine serum,FBS)以色列Biological Industries公司。
1.2 仪器与设备
微生物全自动生长曲线分析仪 芬兰Bioscreen公司;KB-900脱色摇床 海门市其林贝尔仪器制造有限公司;HF90 CO2恒温培养箱 上海力申科技仪器有限公司;Model 680酶标仪 美国Bio-Rad公司;S-4800场发射扫描电子显微镜 日本Hitachi公司;JS680B凝胶成像系统 上海培清科技有限公司。
1.3 方法
1.3.1 菌种活化及菌悬液制备
将冻存于-80 ℃的副溶血性弧菌采用划线法在3% NaCl TSA平板上活化,37 ℃培养12 h后挑取单菌落接种于30 mL 3% NaCl TSB中,将培养液置于37 ℃培养18 h,培养后的菌悬液经离心(5 000×g、5 min,4 ℃)去除上清液,使用pH 7.2磷酸盐缓冲溶液(phosphate buffered saline,PBS)洗涤两次后,使用3% NaCl TSB调整菌悬液在600 nm波长处的OD值为0.5,此时菌悬液浓度约为108CFU/mL。
1.3.2 反式肉桂醛对副溶血性弧菌MIC的测定
参照Shi Chao等[21]的方法,利用液体稀释法测定反式肉桂醛对副溶血性弧菌(2 株标准菌株和8 株分离菌株)MIC。将1.3.1节中所得菌悬液稀释至浓度为5×105CFU/mL,并将100 μL菌悬液加入96 孔酶标板中。向每孔加入反式肉桂醛溶液,使每孔中反式肉桂醛的终质量浓度为200、100、50、25、12.5 μg/mL,随后检测样品的OD600nm。将样品置于37 ℃培养箱中静置培养24 h后,再次检测OD600nm,比较样品培养前后的OD600nm。若两者相差小于0.05,则认为测试质量浓度的反式肉桂醛能够抑制细菌的生长,所对应的最低质量浓度即为反式肉桂醛对副溶血性弧菌的MIC。
1.3.3 反式肉桂醛对副溶血性弧菌ATCC 17802亚抑制浓度的测定
参照Silva-Angulo等[22]的方法测定反式肉桂醛对副溶血性弧菌ATCC 17802的亚抑制浓度。将1.3.1节中最终得到的菌悬液再稀释100 倍,使菌悬液浓度约为106CFU/mL。向百孔蜂窝板中每孔加入125 µL的菌悬液及125 µL的反式肉桂醛溶液(使用3% NaCl TSB配制),使各孔中反式肉桂醛的终浓度分别为MIC、1/2 MIC、1/4 MIC、1/8 MIC、1/16 MIC、1/32 MIC和1/64 MIC。样品对照组添加125 µL菌悬液及125 µL 3% NaCl TSB,背景空白对照组添加250 µL 3% NaCl TSB。设置微生物全自动生长曲线分析仪的培养温度为37 ℃,每隔1 h检测每孔样品在波长600 nm处的OD值并绘制生长曲线,选择质量浓度低于MIC且对副溶血性弧菌的正常生长无明显抑制作用的最高的3 个质量浓度作为本研究的亚抑制浓度。
1.3.4 副溶血性弧菌ATCC 17802泳动能力的测定
反式肉桂醛对副溶血性弧菌ATCC 17802泳动运动性影响的测定参照Shi Chao等[21]的方法。称取LB肉汤培养基0.5 g、氯化钠0.5 g和琼脂0.06 g于20 mL蒸馏水中,加热煮沸至完全溶解,121 ℃高压灭菌,待温度降至45 ℃左右时,向其中加入反式肉桂醛,使其终质量浓度分别为0(对照)、1/64 MIC、1/32 MIC和1/16 MIC,混匀后倒入培养皿,制成泳动平板。冷却30 min后,吸取1.3.1节制备的菌悬液5 µL,接种至泳动平板表面,37 ℃培养7 h。随后使用凝胶成像系统进行拍照,并测量副溶血性弧菌向周围泳动区域(泳动圈)的半径,并计算出泳动圈面积。
1.3.5 副溶血性弧菌ATCC 17802生物被膜形成能力的分析
1.3.5.1 结晶紫染色法定量检测
参照Naves等[23]的方法,选取成膜能力较强的副溶血性弧菌ATCC 17802按照1.3.1节中的方法制备菌悬液,并将菌悬液浓度调整至OD600nm=1.0,配制质量浓度分别为1/32 MIC和1/16 MIC的反式肉桂醛溶液和菌液的混合液,以不含反式肉桂醛、3% NaCl TSB为空白对照,取各组混合液250 µL转移至96 孔酶标板后,分别置于10 ℃和25 ℃生化培养箱培养24、48 h和72 h。测定各时间点时所有孔溶液的OD630nm。随后,吸出菌液,每孔用无菌水漂洗一次,烘干20 min,再向每孔加入1 g/100 mL结晶紫溶液,室温下染色20 min,染色结束后吸出染料,每孔用无菌水漂洗3 次,烘干30 min,然后用冰乙酸溶解。最后使用Model 680酶标仪测定OD570nm。本实验副溶血性弧菌的生物被膜成膜能力用生物被膜形成(specific biofilm formation,SBF)指数表征,其按下式计算。
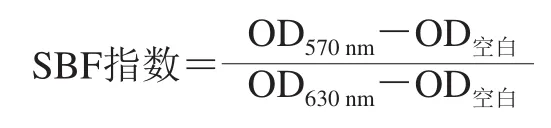
1.3.5.2 场发射扫描电子显微镜定性观察
参考Gowrishankar等[24]的方法,利用场发射扫描电子显微镜定性观察反式肉桂醛对副溶血性弧菌ATCC 17802生物被膜形成能力的影响。按照1.3.1节中的方法稍作修改制备菌悬液,将菌悬液调整至OD600nm=1.0。将反式肉桂醛溶液与菌液等体积混合,使反式肉桂醛终质量浓度分别为0(对照)、1/32 MIC和1/16 MIC,将玻璃片(直径6 mm)浸入混合液中,置于25 ℃生化培养箱培养72 h。取出玻璃片后用无菌水漂洗15 s,再用新的无菌水清洗5 s。随后加入体积分数2.5%戊二醛溶液,置于4 ℃冰箱12 h初步固定。使用PBS和无菌水分别清洗玻片约15 s,再加入没过玻璃片的适量体积分数1%锇酸固定5 h。随后用体积分数30%、50%、70%、80%、90%和100%的乙醇溶液进行梯度洗脱,每次约10 min,静置晾干后进行真空干燥及喷金处理,置于场发射扫描电子显微镜下观察。
1.3.5.3 副溶血性弧菌ATCC 17802黏附及侵入Caco-2细胞能力的测定
反式肉桂醛对副溶血性弧菌ATCC 17802黏附及侵入Caco-2细胞能力的测定参照Amalaradjou等[25]的方法。首先,将浓度为1×105cells/mL处于对数生长期的Caco-2细胞接种于24 孔细胞培养板中恒温培养18 h,随后,使用无菌PBS轻柔清洗3 次。取1.3.1节中菌悬液,添加反式肉桂醛溶液,使反式肉桂醛的质量浓度分别为0(对照)、1/64 MIC、1/32 MIC和1/16 MIC,将样品置于37 ℃恒温摇床(100 r/min)培养6 h。取出样品进行离心(5 000×g、5 min,4 ℃),使用含体积分数10% FBS的DMEM培养液稀释菌体浓度至106CFU/mL。向预先培养的细胞中每孔分别加入1 mL菌悬液后离心(600×g、5 min),随后将上清液放入37 ℃、5% CO2细胞培养箱中恒温培养1 h。
在黏附实验中,恒温培养1 h后取出样品并吸出孔板内液体,使用无菌PBS清洗细胞3 次。向每孔中加入1 mL体积分数0.1% Triton X-100,置于4 ℃下处理20 min后使细胞裂解,使用移液器取孔中液体来回吸打,使得细胞及其碎片充分悬浮于液体中,并将所有液体全部转移至新的离心管中。将离心管中液体进行10 倍梯度稀释后涂布于3% NaCl TSA上,于37 ℃培养24 h后读数,计算黏附Caco-2细胞的菌体数量。相对黏附率以处理组黏附菌量占对照组黏附菌量的比例表示。
在侵入实验中,恒温培养1 h后取出样品并吸出孔板内液体,使用无菌PBS清洗细胞1 次。向每孔中加入1 mL含有100 µL/mL庆大霉素和体积分数1% FBS的DMEM培养液,于37 ℃、体积分数5% CO2环境下培养30 min以杀灭Caco-2胞外细菌。随后离心(600×g、5 min)去除上清液并使用无菌PBS清洗3 次,向每孔中加入体积分数0.1% Triton X-100,并置于4 ℃下作用20 min以裂解细胞。最后,使用无菌PBS 10 倍梯度稀释进行涂布,37 ℃下培养24 h后,读数并计算侵入Caco-2细胞的菌体数量。相对侵入率以处理组侵入菌量与对照组侵入菌量的比值表示。
1.4 数据处理与分析
每个实验进行独立的3 次重复实验,采用Excel软件进行数据统计和作图,结果以平均值±标准差的形式表示,并通过SPSS 19.0软件对数据进行方差分析,采用T检验进行显著性检验,P<0.05表示差异显著,P<0.01表示差异极显著。
2 结果与分析
2.1 反式肉桂醛对副溶血性弧菌的MIC结果
反式肉桂醛对副溶血性弧菌MIC的测定选取了2 种标准菌株和8 种分离菌株进行,结果见表1。反式肉桂醛对副溶血性弧菌标准菌株ATCC 17802的MIC为50 μg/mL,对ATCC 33847的MIC为100 μg/mL,对8 株分离菌的MIC均为100~200 μg/mL。由于副溶血性弧菌ATCC 17802具有泳动运动性、生物被膜形成能力、黏附及侵入Caco-2细胞能力等毒力表征[26],因此选择ATCC 17802作为后续研究对象。
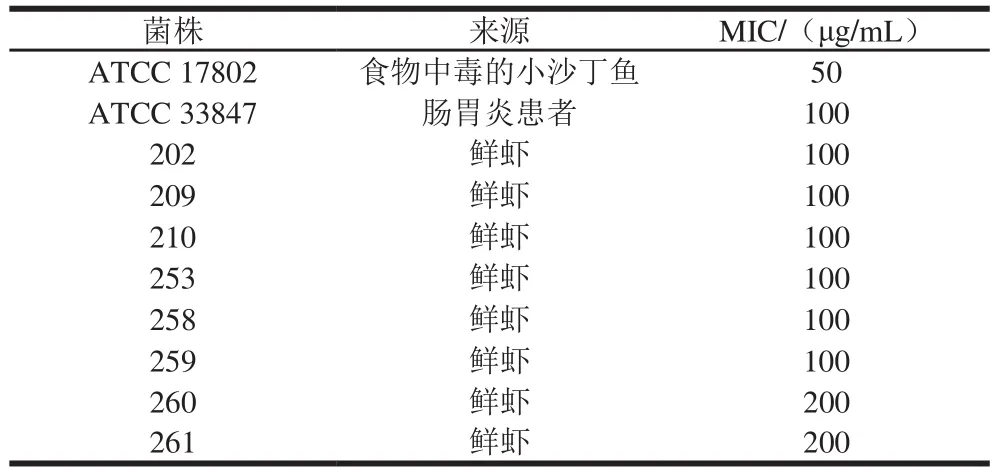
表1 反式肉桂醛对副溶血性弧菌的MICTable 1 MIC of trans- cinnamaldehyde against Vibrio parahaemolyticus
2.2 反式肉桂醛对副溶血性弧菌ATCC 17802的亚抑制浓度
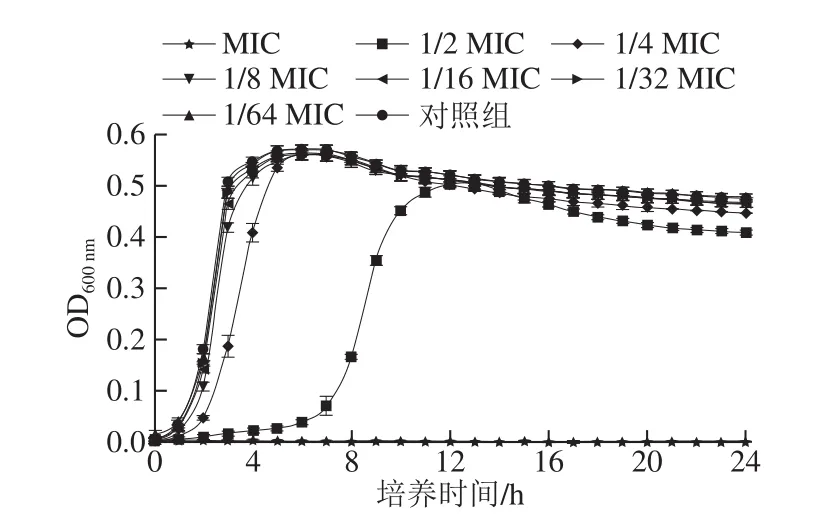
图1 反式肉桂醛对副溶血性弧菌ATCC 17802生长曲线的影响Fig.1 Effect of trans-cinnamaldehyde on the growth curve of Vibrio parahaemolyticus ATCC 17802
反式肉桂醛对副溶血性弧菌ATCC 17802的生长曲线的影响如图1所示。与对照组相比,当反式肉桂醛质量浓度为1/2 MIC时细菌的延滞期明显延长。当反式肉桂醛质量浓度为1/4 MIC时细菌的对数生长期与对照组相比有明显的差异。当反式肉桂醛质量浓度为1/16 MIC、1/32 MIC、1/64 MIC时,细菌的生长情况与对照组相比几乎无差异,细菌可正常生长繁殖,因此,本研究选择1/16 MIC、1/32 MIC、1/64 MIC为反式肉桂醛对副溶血性弧菌的亚抑制浓度。
2.3 反式肉桂醛对副溶血性弧菌ATCC 17802泳动运动性的影响
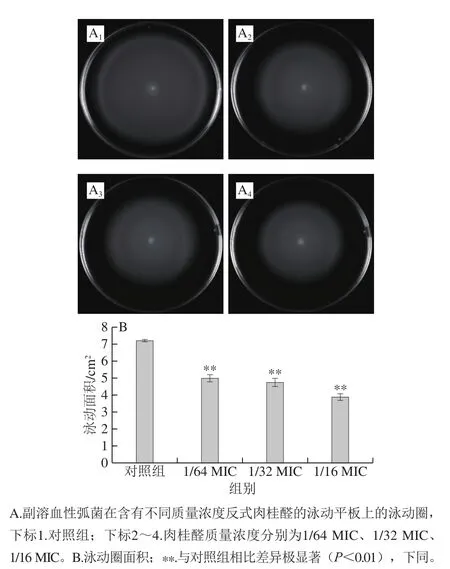
图2 反式肉桂醛对副溶血性弧菌ATCC 17802泳动能力的抑制作用Fig.2 Trans-cinnamaldehyde inhibited swimming motility of Vibrio parahaemolyticus ATCC 17802
反式肉桂醛对副溶血性弧菌ATCC 17802泳动运动性的影响如图2所示。经过反式肉桂醛处理的菌体泳动圈面积与对照组相比明显减小。对照组中副溶血性弧菌的泳动圈面积为(7.22±0.05)cm2。与对照组相比,经1/64 MIC的反式肉桂醛处理后,副溶血性弧菌ATCC 17802的泳动圈面积极显著降低至(5.01±0.18)cm2(P<0.01),1/32 MIC和1/16 MIC的反式肉桂醛处理后,泳动圈面积分别极显著降低至(4.76±0.22)cm2(P<0.01)和(3.89±0.19)cm2(P<0.01)。因此,亚抑制浓度的反式肉桂醛对副溶血性弧菌的泳动运动性有良好的抑制作用。
2.4 反式肉桂醛对副溶血性弧菌ATCC 17802生物被膜形成能力的影响
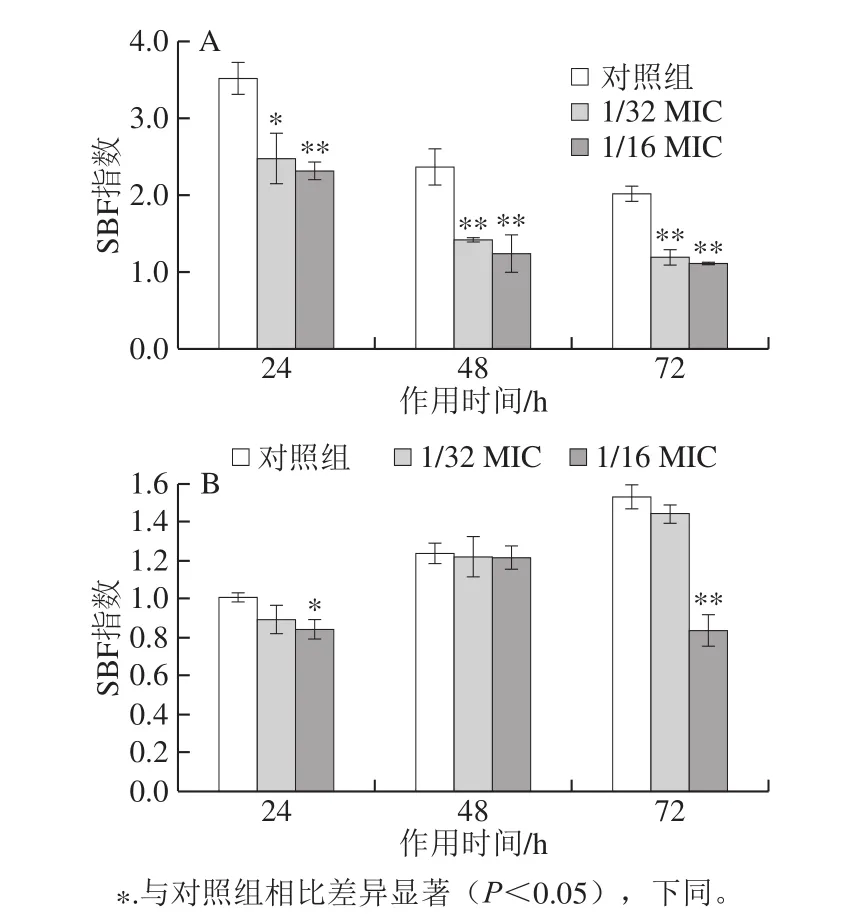
图3 不同质量浓度反式肉桂醛在10 ℃(A)和25 ℃(B)作用下对副溶血性弧菌ATCC 17802生物被膜形成能力的影响Fig.3 Effect of different concentrations of trans-cinnamaldehyde on the biofilm formation ability of Vibrio parahaemolyticus ATCC 17802 at 10 ℃ (A) and 25 ℃ (B)
如图3所示,副溶血性弧菌有较强的生物被膜形成能力,与对照组相比,在10 ℃下反式肉桂醛作用24 h后,1/32 MIC和1/16 MIC浓度条件使副溶血性弧菌ATCC 17802生物被膜形成量分别下降了29.59%和33.91%;作用48 h后分别极显著下降了37.89%和47.31%(P<0.01);作用72 h后分别极显著下降了40.86%和44.08%(P<0.01)(图3A)。与对照组相比,不同质量浓度的反式肉桂醛对副溶血性弧菌ATCC 17802生物被膜的形成量均有抑制作用,并且呈浓度依赖性。
如图3B所示,1/16 MIC的反式肉桂醛在25 ℃下作用24 h后对副溶血性弧菌的生物被膜形成能力有显著的抑制作用(P<0.05);反式肉桂醛作用48 h后对副溶血性弧菌的生物被膜形成能力没有显著的抑制作用(P>0.05);在72 h时,经1/32 MIC反式肉桂醛处理的副溶血性弧菌ATCC 17802生物被膜形成量下降了5.68%,而1/16 MIC浓度使生物被膜形成量极显著下降了45.44%(P<0.01)。
2.5 副溶血性弧菌ATCC 17802形成的生物被膜微观结构
如图4所示,对照组中副溶血性弧菌的生物被膜致密均匀,分布紧凑,布满整个视野(图4A);而在1/32 MIC反式肉桂醛作用下,菌体形成的生物被膜稀疏、松散、分布不均(图4B);经1/16 MIC反式肉桂醛处理后,菌体形成生物被膜时,生物被膜中的细菌量明显减少,生物被膜结构更加松散(图4C)。结果表明,反式肉桂醛能够抑制副溶血性弧菌ATCC 17802生物被膜的形成能力,且呈质量浓度依赖性。

图4 副溶血性弧菌ATCC 17802形成的生物被膜扫描电子显微镜图Fig.4 Scanning electron micrographs of biofilm formed by Vibrio parahaemolyticus ATCC 17802
2.6 反式肉桂醛对副溶血性弧菌ATCC 17802黏附及侵入Caco-2细胞能力的影响
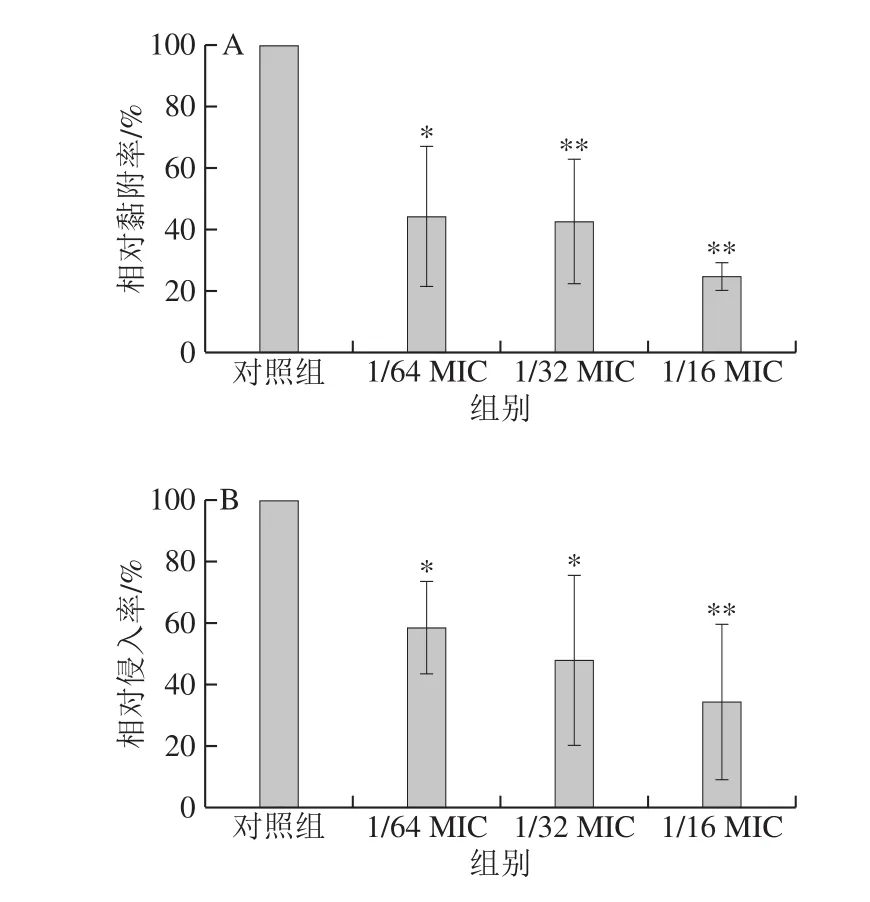
图5 反式肉桂醛对副溶血性弧菌ATCC 17802黏附(A)及侵入(B)Caco-2细胞的影响Fig.5 Effect of trans-cinnamaldehyde on the ability of Vibrio parahaemolyticus ATCC 17802 to adhere to (A) and invade (B) Caco-2 cells
如图5A所示,反式肉桂醛能够显著降低副溶血性弧菌黏附Caco-2细胞的能力,并且呈浓度依赖性,当反式肉桂醛质量浓度为1/64 MIC时,细菌的黏附率降低至对照组的44.2%;当反式肉桂醛质量浓度为1/32 MIC和1/16 MIC时,细菌的相对黏附率分别降至42.9%和24.9%,与对照组呈现极显著差异(P<0.01)。
由图5B可知,反式肉桂醛对副溶血性弧菌ATCC 17802侵入Caco-2能力具有明显的抑制作用,并且随反式肉桂醛质量浓度的增加,相对侵入率逐渐下降。与对照组相比,当反式肉桂醛质量浓度为1/64 MIC和1/32 MIC时,细菌的相对侵入率分别降至对照组的58.2%和48.3%,与对照组呈显著性差异(P<0.05);当反式肉桂醛质量浓度为1/16 MIC时,细菌的相对侵入率降至对照组的34.5%,与对照组呈极显著差异(P<0.01)。
3 讨 论
本研究首先测定了反式肉桂醛对副溶血性弧菌的MIC,以评价反式肉桂醛对副溶血性弧菌的抑菌功效。结果表明,反式肉桂醛对副溶血性弧菌的MIC为50~200 μg/mL,抑菌效果良好。选择1/16 MIC、1/32 MIC、1/64 MIC为反式肉桂醛对副溶血性弧菌的亚抑制浓度,在亚抑制浓度下,细菌的生长及存活不会受到影响,因此实验可排除反式肉桂醛对细菌存活及生长的影响。在Li Guanghui等[27]研究安石榴苷对沙门氏菌泳动运动能力和侵入HT29细胞能力的影响时,选用了1/16 MIC和1/32 MIC为亚抑制浓度;Sivaranjani等[28]发现,亚抑制浓度(1/4 MIC、1/8 MIC、1/16 MIC)的桑色素可对单增李斯特菌的毒力因子产生有效的抑制作用;本实验研究了亚抑制浓度(1/16 MIC、1/32 MIC、1/64 MIC)反式肉桂醛对副溶血性弧菌运动性、生物被膜形成能力、黏附及侵入Caco-2细胞能力的影响,旨在共同阐述反式肉桂醛对副溶血性弧菌毒力因子的抑制作用。
副溶血性弧菌的运动性与其毒力密切相关,并且影响着其对宿主细胞的黏附及侵入能力[26]。细菌依靠鞭毛的定向运动是病原菌作用于宿主细胞并引发感染的第一步,因此抑制病原菌的运动性可有效降低其感染能力[29]。在本研究中,与对照相比,经亚抑制浓度反式肉桂醛作用后的副溶血性弧菌的泳动圈直径极显著下降,表明反式肉桂醛能够有效抑制副溶血性弧菌的运动能力。Upadhay等[30]证明了亚抑制浓度的反式肉桂醛能将空肠弯曲杆菌的泳动运动能力降低70%以上;Packiavathy等[31]发现100 μg/mL的姜黄素可使副溶血性弧菌的泳动运动能力显著降低;Bai Jinrong等[32]证明0.312 5、0.625 mg/mL和1.250 mg/mL的莽草酸可抑制金黄色葡萄球菌的运动能力。本研究中,亚抑制浓度的肉桂醛可显著降低副溶血性弧菌的运动性,从而降低了副溶血性弧菌侵染宿主引起感染的可能性。Guo Du等[26]研究发现,百里醌可通过降低鞭毛合成基因flaA、flaM和flaL的相对转录量来抑制副溶血性弧菌的运动性,而反式肉桂醛是否是通过降低flaA、flaM和flaL基因相对转录量来阻碍副溶血性弧菌鞭毛的合成,从而抑制副溶血性弧菌的运动能力,还有待进一步探讨。
细菌的生物被膜可使细菌产生对抗生素的抗性[33],并且生物被膜形成的封闭基质还能够增加细菌对环境压力的耐受性[34]。在该研究中,采用结晶紫染色法将结晶紫与胞外多糖结合,检测洗脱液的SBF指数,进行生物被膜定量检测,并通过场发射扫描电子显微镜对样品进行电子照射,定性地观察由副溶血性弧菌形成的生物被膜。实验结果表明,随着反式肉桂醛质量浓度的增加,副溶血性弧菌生物被膜的形成量相比于对照组下降明显,并且生物被膜结构松散。证明了反式肉桂醛对副溶血性弧菌的生物被膜形成能力具有良好的抑制作用。Kang Jiamu等[35]利用结晶紫染色法发现,0.25~2.00 mg/mL的没食子酸可显著抑制福氏志贺菌生物被膜的生成量;Fan Qiuxia等[36]利用场发射扫描电子显微镜观察发现,1/16 MIC和1/32 MIC的辅酶Q0可以使单增李斯特菌形成的生物被膜变薄、结构变松散;张芳等[37]用1、5、10 μg/mL的白藜芦醇处理单增李斯特菌后,其生物被膜形成量分别减少10.40%、21.03%和39.10%。在本研究中,反式肉桂醛能够有效抑制副溶血性弧菌的生物被膜生成量,这会使得副溶血性弧菌对环境压力的抵抗能力降低,从而降低其危害性。群体感应是影响细菌生物被膜形成的主要因素之一[38],细菌间的群体感应由细菌合成、分泌的AI-2信号分子所调控,而由luxS基因编码合成的LuxS蛋白是AI-2分子合成的关键酶[39]。反式肉桂醛是否能通过抑制luxS基因的相对转录量来抑制其生物被膜的形成,还需要进行进一步探究。
黏附及侵入宿主肠上皮细胞是食源性致病菌感染宿主的必要途径[40]。有研究表明,病原菌的黏附作用可使细菌黏附在宿主细胞表面,并且黏附作用为致病菌侵入宿主细胞进而引发宿主的感染提供了有利条件[41]。副溶血性弧菌能够通过口腔进入人体胃肠道,进而黏附及侵入肠上皮细胞引发感染。因此,抑制副溶血性弧菌黏附及侵入肠上皮细胞的能力是降低其毒力的有效方法。本研究结果表明,反式肉桂醛能够降低副溶血性弧菌黏附及侵入Caco-2细胞的能力。在先前的研究中,Shi Chao等[21]证明了1/16 MIC、1/32 MIC、1/64 MIC柠檬醛可有效降低阪崎克罗诺肠杆菌黏附及侵入Caco-2细胞的能力;Inamuco等[11]研究表明,虽然香芹酚不会影响沙门氏菌的黏附能力,但会使其侵入肠上皮细胞的能力降低。本研究结果表明,反式肉桂醛可降低副溶血性弧菌侵染宿主细胞的能力,降低副溶血性弧菌对人体的感染能力。Sun Yi等[2]研究发现,副溶血性弧菌黏附及侵入宿主细胞的能力与副溶血性弧菌的运动能力有关,因此推测,副溶血性弧菌黏附及侵入Caco-2细胞能力的减弱是由于反式肉桂醛阻碍了副溶血性弧菌鞭毛的合成,导致其运动性减弱。
综上所述,反式肉桂醛对副溶血性弧菌有良好的抑制效果,其对副溶血性弧菌的MIC为50~200 μg/mL;并且亚抑制浓度的反式肉桂醛可抑制副溶血性弧菌的多种毒力因子,包括副溶血性弧菌的泳动运动能力、生物被膜形成能力、黏附及侵入Caco-2细胞能力。以上研究结果表明,反式肉桂醛具有降低副溶血性弧菌对人体的感染能力的潜力,这为反式肉桂醛的进一步开发应用于控制副溶血性弧菌引起的食源性疾病提供了理论依据。
