Ectoine高产菌株Halomonas sp. XH26的鉴定及紫外诱变选育
2020-08-19沈国平龙启福朱德锐
田 磊, 张 芳, 沈国平, 高 翔, 龙启福, 朱德锐
(青海大学 医学院 基础医学研究中心, 西宁 810016)
Ectoine(1, 4, 5, 6-tetrahydro-2-methyl-4-pyrimidnecarboxylic acid, C6H10N2O2)即1, 4, 5, 6-四氢-2甲基-4-嘧啶羧酸,是一种氨基酸衍生物,广泛存在于嗜盐古菌、嗜盐细菌、盐藻及真菌等胞内[1]。Ectoine是一种抵抗外界高盐环境的有机相容性溶质,具有增强酶活性、稳定DNA结构及保护蛋白质的功能[2]。国内外诸多文献报道,已将其广泛地应用于消化道疾病、干眼综合征、过敏性鼻炎、皮肤创伤与保健美容等生物医学领域[3-4],其研发产品涉及消化道药物、滴眼液、鼻喷剂、皮肤创伤药及新型生物化妆品等[5-6]。目前,Ectoine大规模生产主要以微生物发酵为主,模式菌株主要有盐单胞菌属(Halomonas)与色盐杆菌属(Chromohalobacter),如H.elongata(DSM 2581T、1A01717与BK-AG18)和C.salexigensDSM 3043T等[7-8]。相关研究主要集中于菌株优选和发酵工艺与条件的优化。Ectoine的发酵工艺主要包括实验室条件下的单批次发酵(试管液体培养、浅层液体培养、摇瓶培养与台式发酵罐等),中试或工业级大规模生产(分批发酵罐法、流加发酵罐法与细菌挤奶Bacterial milking)等,或采用不同发酵方式联用,以提高Ectoine产量[9]。
针对Ectoine积聚菌株的研究,主要集中在野生菌株的分离筛选、基因工程菌株的构建(生物合成Pathway基因和ectABC操纵子等)、模式菌株代谢流量控制发酵(Metabolic overflow,包括Carbon-limited fed-batch cultures、Metabolic flux analysis与Metabolic flux distribution)及优化[10-12]。然而,以上不同的研究方法,各具优劣,其总体产量提高30%~79%[13-14]。参考其他微生物代谢产物的菌种改良研究,原始菌株的生产能力可通过物理(紫外诱变)或化学(硫酸二乙酯和甲基磺酸乙酯等)突变方式大幅度提高。紫外诱变[15](UV mutagenesis)是一种经典微生物诱变方法,菌株对诱变因素极具敏感性,且正向突变率高,能有效提高生产菌株的产量,然而国内外暂无Ectoine菌株诱变研究的相关报道。至此,本研究以小柴旦盐湖盐单胞菌Halomonassp. XH26为研究对象,采用9轮紫外线循环诱变选育遗传稳定的Ectoine高产突变菌株,以期为Ectoine大规模发酵生产提供新的菌源,提高Ectoine的发酵生产效率。
1 材料与方法
1.1 材料
1.1.1 菌株来源与培养基
野生菌株Halomonassp. XH26来源于青海大学基础医学研究中心的保存菌株,该菌株分离于柴达木盆地小柴旦盐湖。OSM分离培养基[1](g/L):NaCl 50.0 g,MgSO4·7H2O 20.0 g,柠檬酸钠3.0 g,KCl 2.0 g,CaCl20.2 g,细菌蛋白胨10.0 g,酵母抽提物2.0 g,调pH 7.5,固体培养基另加琼脂15.0 g。发酵培养基(g/L):NaCl 60.0 g,MgSO4·7H2O 25.0 g,KCl 55.0 g,L-谷氨酸钠6.5 g和酶水解酪素7.5 g,调pH 8。
1.1.2 主要试剂与设备
NaCl、KCl、MgSO4·7H2O、柠檬酸钠、酵母、L-谷氨酸钠、酶水解酪素和NaOH等分析纯(天津永大化学试剂公司);乙腈(HPLC级, 美国Fisher scientific公司);Ectoine 标准品(HPLC级 > 95%, 德国Fluka Analytical公司);超净工作台(上海智城公司);第三代变速组织研磨器(OSEY50型,北京天根公司);低真空扫描电镜(JSM-561ULV型, 日本电子光学公司);高效液相色谱仪(HPLC1260型,美国安捷伦公司);色谱柱(Merck-SeQuant ZIC-HILIC,150×4.6 mm,5 μm);恒温摇床(ZD-85,上海精达公司);微孔过滤器(0.22 μm水系,天津亳津公司)。
1.2 方法
1.2.1 菌种活化与菌悬液的制备
菌株活化。将Halomonassp. XH26菌株接种于5 mL OSM培养基中,160 r/min,35 ℃恒温振荡培12 h活化,测定OD600值,大量接种备用。菌悬液的制备:取上述对数生长期的菌液5 mL,3000 r/min 离心10 min,弃清液,加入0.9%的无菌生理盐水振荡洗涤3次,后重悬于生理盐水中,调整菌悬液浓度约为108CFU/mL[16]。
1.2.2 产Ectoine菌种的鉴定
将菌株接种于OSM固体培养基上,35 ℃培养24 h,观察菌落形态,革兰染色,在光学显微镜下观察菌体形态,参照《微生物学实验》《常见细菌系统鉴定手册》及《伯杰氏鉴定细菌学手册》对菌株进行生长特性与生化鉴定[17]。16S rRNA基因测序分析由上海生工生物工程公司完成,运用BLAST搜索同源序列,通过软件Mega v 7.0构建系统发育树[18]。
1.2.3 细菌电镜样品的制备
细菌处理和固定。细菌过夜培养后,取2 mL菌液(OD600约1.20),8000 r/min离心3 min,弃上清;加入500 μL PBS洗2~3次,3000 r/min离心3 min,弃上清;加入2.5%戊二醛,混匀,4 ℃过夜固定。乙醇梯度脱水:固定处理液,8000 r/min离心1 min,弃上清,PBS溶液清洗3次,按30%~90%浓度梯度脱水;每次加入乙醇脱水后静置15 min,8000 r/min离心1 min后100%乙醇洗涤2次,条件同前;最后用乙酸异戊酯置换2次,每次20 min,最后一次3000 r/min离心3 min,弃上清;沉淀物经-20 ℃、-40 ℃与-80 ℃分别冷冻6 h后,在-65 ℃低温干燥箱中冷冻干燥12 h,粘样电镜检测[19-20]。
1.2.4 紫外诱变
将108CFU/mL菌悬液稀释为10-3、10-4、10-5与10-6浓度,各取5 mL稀释液置于90 mm的培养皿中,放入无菌磁力搅拌子,后置于磁力搅拌器上,在紫外灯下(功率25 W,照射距离30 cm,提前预热30 min)分别照射10~70 s(间隔10 s)。取照射后的菌悬液0.1 mL均匀涂布平板(每梯度3~5个),以未经照射的菌悬液为对照,后将平板置于35 ℃培养箱,培养48 h计数致死率和正突变率。紫外诱变后选取菌落较大、长势良好、菌落边缘黏稠且不易拉丝的突变菌株,接种于发酵培养基中培养36 h,测定Ectoine产量。采用9轮循环紫外照射进行诱变,每轮诱变后连续培养3次,筛选Ectoine产量最高菌株进行下一轮诱变。正突变率(%)=正突变株数/诱变后菌落数×100%;致死率(%)=(对照CFU-处理CFU)/对照CFU×100%[21-23]。
1.2.5 细胞干重测定(CDW g/g)
将100 mL发酵液4 ℃ 12 000 r/min离心5 min,弃上清,再加10 mL的生理盐水快速洗涤,8000 r/min 离心5 min,弃上清,将沉淀放于70 ℃烘干箱至恒重,干燥后称量,计算细胞干重(CDW g/g)。
1.2.6 Ectoine的提取
菌株发酵培养36 h后测OD600,12 000 r/min离心5 min,弃上清;加入99%的乙醇10 mL,用组织研磨器高速研磨3~4 min,12 000 r/min离心5 min,取出上清至10 mL烧杯中,放入60 ℃烘干箱中烘干4 h;再加入2 mL超纯水,重溶Ectoine后经0.22 μm水系微孔过滤器过滤,即为Ectoine提取液。
1.2.7 高效液相色谱(HPLC)检测Ectoine含量
配制1.0 mg/mL的Ectoine标准品母液(间隔0.2 mg/mL),通过高效液相色谱(HPLC)检测Ectoine含量,采用高效峰面积和浓度制作标准曲线。HPLC检测条件:乙腈为流动相,检测波长210 nm,流速为1.0 mL/min,柱压3.486~4.761 MPa,柱温30 ℃,胞内Ectoine抽提液上样量10 μL[24-25]。
1.2.8 遗传稳定性
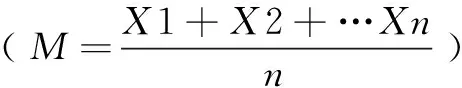
2 结果与分析
2.1 Halomonas sp.XH26菌体形态特征与同源性比对
Halomonassp.XH26菌株在OSM培养基上,菌落形态呈圆形(图1-A),大小适中,易粘连,乳白色,隆起,湿润状,边缘规则,不透明,革兰氏阳性菌(图1-B)。电镜下菌体形态呈长杆状(图1-C),略带弯曲,菌体的长度为(3~5)μm×(0.5~0.8)μm,两端圆润,无鞭毛。生理生化试验结果如表1所示。利用NCBI Blast分析比对XH26的16S rRNA基因序列,构建系统发育树(图2)。分析表明:菌株XH26位于盐单胞菌属进化分支单元,与嗜碱盐单胞菌H.alkaliphila2B8N(JF710970)和美丽盐单胞菌H.venustaDSM 4743(NR042069)类聚,进化趋近同源;XH26与H.alkaliphilaMAT-16(KC354706)、H.venustaB19b(JF710983)、H.alkaliphilaR4(KX495192)的相似性均为99%,进化高度同源,但在系统发育方向上,3者之间具有明显的差异。由图2可知,菌株XH26与HalomonascampaniensisLS21(CP007757)亲缘关系近,结合上述的生理生化鉴定结果,可将菌株XH26鉴定为Halomonascampaniensis。
2.2 紫外诱变最佳诱变时间的确定
紫外线照射时间对菌株的致死率和正向突变率的关系如图3所示。当紫外照射时间为50 s后菌株致死率达到80%以上,70 s以后菌株致死率达到了100%。随紫外照射时间的延长,菌株的正突变率也在增加。由于短时间照射不能达到较稳定的突变状态[24],按照诱变剂量选择的一般原则[15],致死率为70%~80%的剂量,菌株突变性可以保持在较稳定的水平。当紫外线处理50 s时致死率为75%,至此本研究选取50 s为最佳诱变时间。

表1 Halomonas sp. XH26的生理生化特征
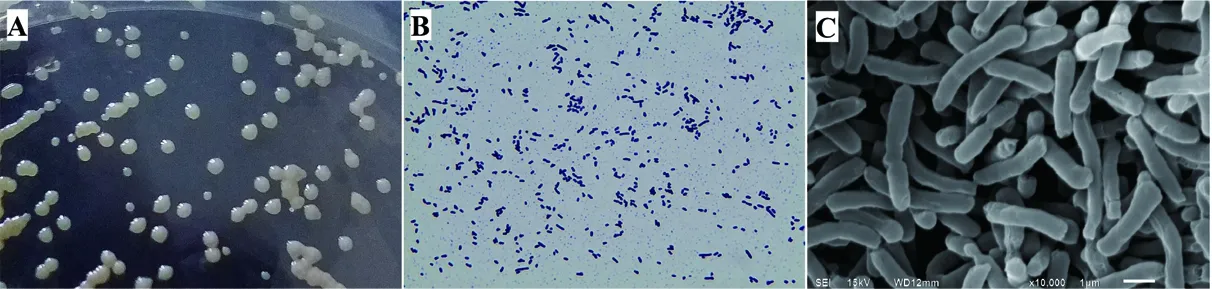
A:菌落形态;B:革兰氏染色油镜(×100);C: 扫描电镜(×10 000),电压15 kV,工作距离WD12 mm,Bar 1μm)
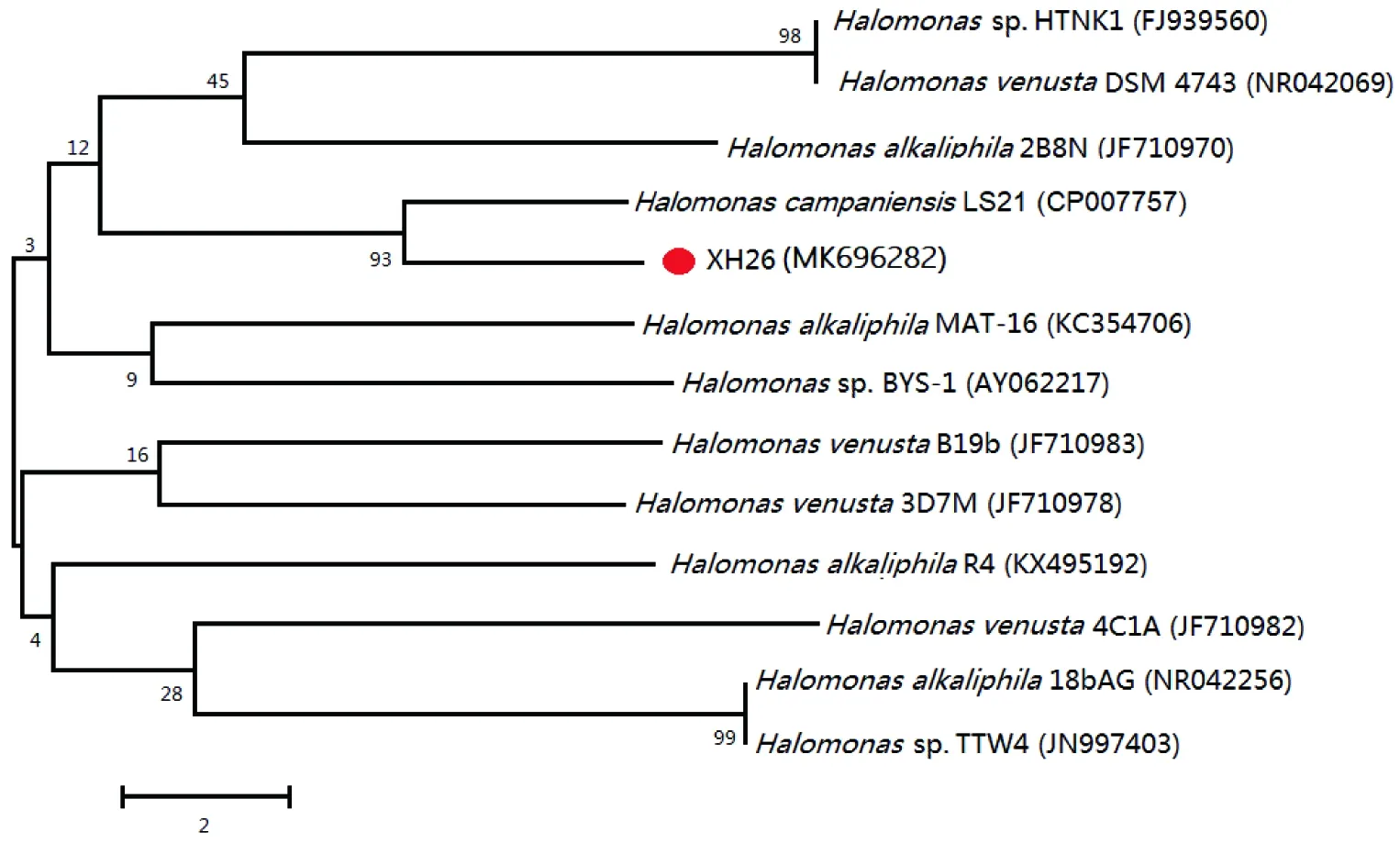
图2 菌株XH26的系统发育树
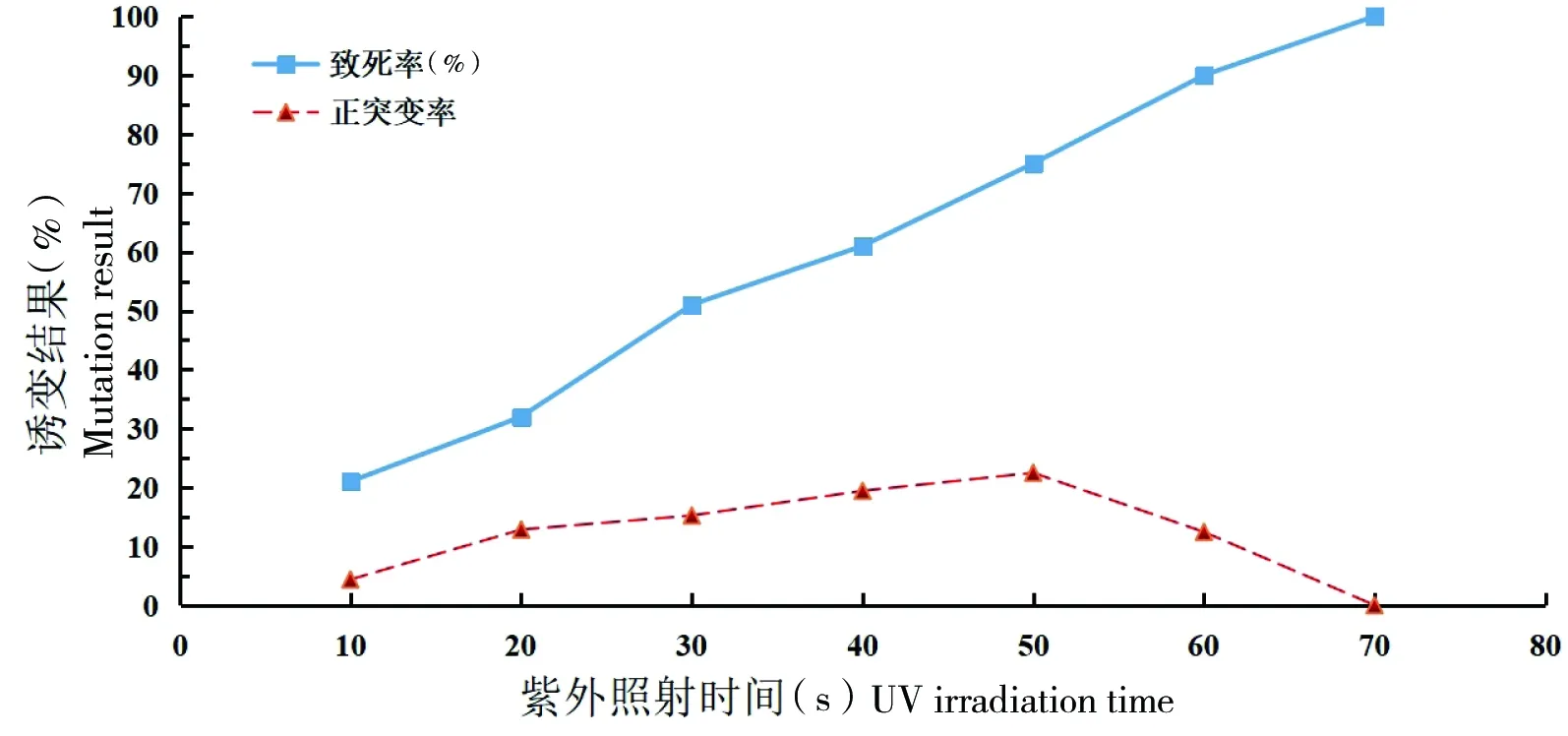
图3 紫外线照射时间对菌株的致死率和正突变率的影响
2.3 产物鉴定结果
采用高效液相色谱法(HPLC)绘制标准曲线(图4-A)对突变菌株产物进行分析。Ectoine标准品检测结果如图4-B所示,选取突变菌株HU09-32提取Ectoine(图4-C)。通过高效液相色谱图比对,表明利用乙醇抽提突变菌株胞内产物和Ectoine标准品的校正保留时间均为10 min,完全一致,应为同一物质,确定突变菌株能够积累Ectoine。
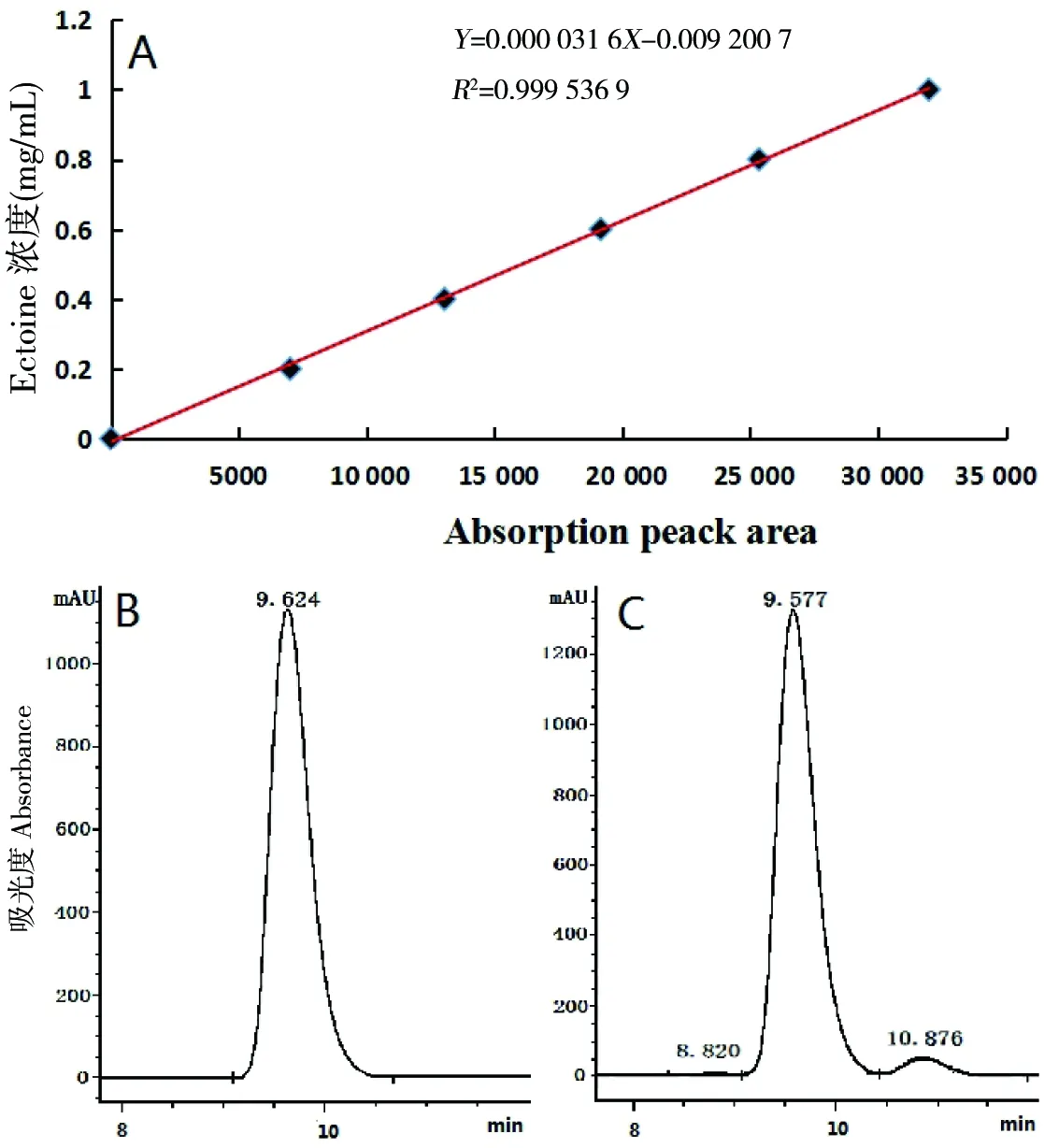
A:Ectoine标准曲线;B:标准品吸收峰型;C:突变菌株HU09-32提取Ectoine吸收峰型
2.4 紫外诱变突变菌株筛选
经过9轮循环诱变,得到272株突变菌,最终筛选出7株Ectoine产量较高的突变菌株(表2)。由表2可知,诱变菌株HU09-6、HU09-12及HU09-71的Ectoine产量相当,分别为原始菌株的2.1、2.4及2.1倍,HU09-21、HU09-61与HU09-62菌株Ectoine产量分别为1154.29、1165.16与1202.98 mg/L,其中HU09-32菌株Ectoine产量最高,达到1351.09 mg/L,为原始菌株的2.9倍,具有显著差异(P< 0.05)。因此,选取HU09-32菌株作为后期研究的目标菌株。
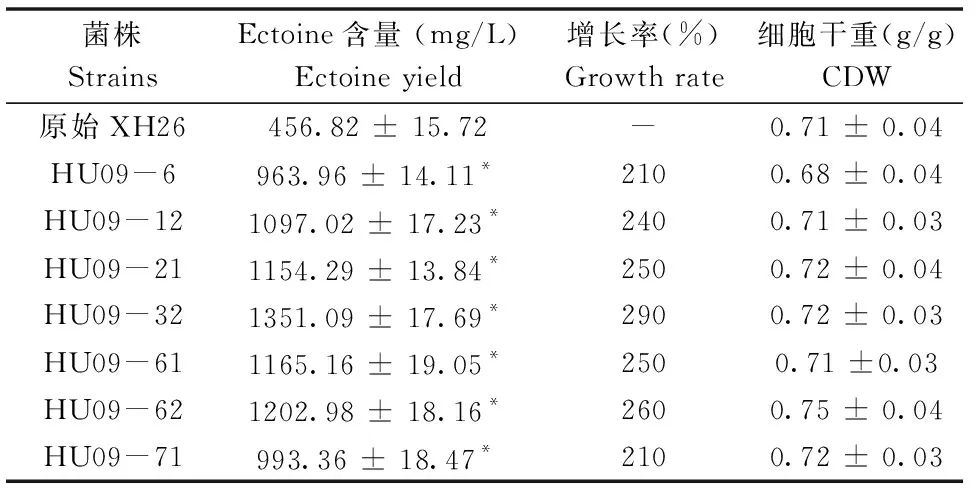
表2 紫外诱变突变菌株Ectoine产量
2.5 突变菌株稳定性检测
选取Ectoine产量在950 mg/L以上的7株突变型菌株:HU09-6、HU09-12、HU09-21、HU09-32、HU09-61、HU09-62与HU09-71进行连续培养,HPLC检测Ectoine产量。结果分析(表3)表明:通过40 d连续培养(接种2 d/次)后,7株突变型菌株Ectoine产量均未出现明显降低,每组实验结果无明显差异,表现出一定的遗传稳定性,且均明显优于原始菌株。HU09-12、HU09-21与HU09-61稳定性较好,Ectoine产量保持在1000 mg/L左右。HU09-6和HU09-71菌株Ectoine产量较稳定,但是在30 d后出现轻度降低的趋势。HU09-32和HU09-62的稳定性最好,Ectoine产量均未低于1200 mg/L,具有发酵生产应用潜力。
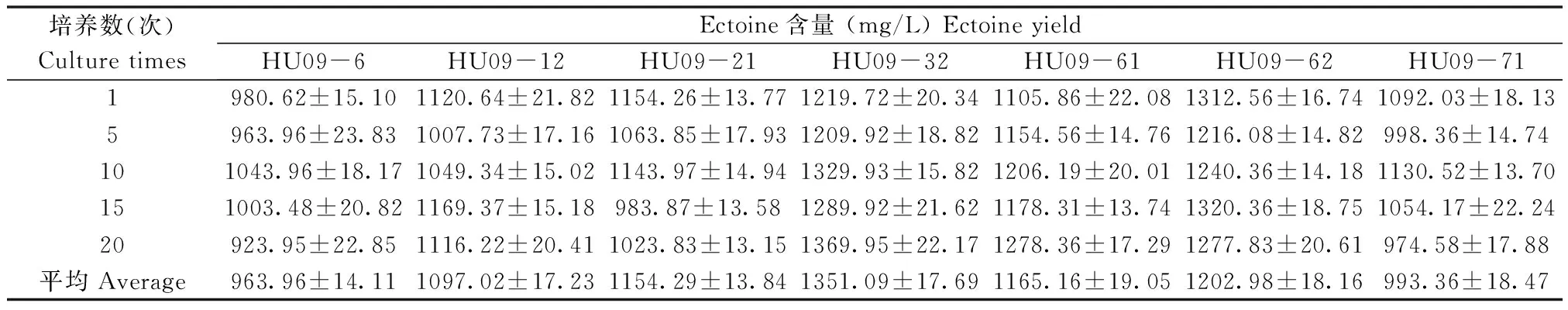
表3 突变株遗传稳定性的测定
3 讨论与结论
Ectoine工业生产的关键问题在于菌株的分离与选育,目前研究主要集中在野生菌株的分离筛选、发酵条件的优化及构建基因工程菌株[10-11]。周月慧等[27]单批次摇瓶发酵HalomonasvenustaSL 21(48 h),诱导生成的Ectoine产量为285.1 mg/L。张晓梅等[28]实验室摇瓶发酵Halorubrumaidingense菌株,显示Ectoine的最大产量为321.28 mg/L。Becker等[29]将伸长盐单胞菌(H.elongata)的Ectoine合成基因簇ectABC整合到谷氨酸棒状杆菌(C.glutamicum)的基因组,构建基因工程菌,经分批补料发酵Ectoine的产率为6.7 g/L·d。对比以上研究,Ectoine产量均在不同程度上获得提升,但是增长有限。目前,国内外暂无对产Ectoine菌株进行物理或化学方法诱变处理,来获得高产Ectoine突变菌株的研究报道。
基于紫外诱变是获得细菌突变体的经典方法,其诱变效应为引起细菌DNA结构或碱基序列的变化,导致基因发生突变[30-31]。目前,该方法广泛用于不同微生物发酵产物的研究中。张云野等[32]对休哈塔假丝酵母(Candidaalbicans)HDYXHT-01进行紫外诱变,获得的突变株乙醇产量提高3.03%~12.88%。刘俊梅等[21]采用紫外线和硫酸二乙酯相结合的诱变方法对P.koreensisPK3菌株进行复合诱变,获得一株高产PHB突变菌株,PHB产量达到15.94 g/L,较原始菌株提高了2.61倍。田连生和陈秀清[22]通过紫外线和甲基磺酸乙酯(EMs)对野生菌株XN-3进行复合诱变,选育突变菌株UH-2-4,其絮凝率达到96.8%,比出发菌株提高了25.1%。本研究通过比较紫外线处理时间对Halomonassp. XH26菌株与Ectoine产量的影响,发现紫外线处理50 s,能有效提高Ectoine的产量;同时,可以保持突变株胞内Ectoine产量的稳定。经9轮循环紫外诱变筛选,获得7株遗传稳定性较好的突变株,Ectoine产量均达到出发菌株2倍以上,其中HU09-32 Ectoine产量最高,为1351.09 mg/L±17.69 mg/L,是原始菌株Ectoine产量(Halomonassp. XH26发酵条件优化后,456.82 mg/L±15.72 mg/L,CDW 0.71 g/g±0.04 g/g)的2.9倍。
本次研究采取多轮紫外循环诱变,筛选获得Ectoine高产突变菌株(200%以上)。经过连续培养(40 d),其Ectoine的产量表现出良好的稳定性,显示出很好的生产和应用前景。但实验中发现,随着诱变代数的增加,诱变菌株亦表现出生长速度降低、Ectoine产量减少及菌落形态改变等现象。在本实验中,获得7株突变株的Ectoine产量稳定,没有回复亲本菌株的迹象,明显优于原始菌株。后续我们将会在分子水平上对突变菌株与原始菌株产Ectoine基因进行探究,为进一步提升菌株性能提供可靠的实验依据与实验菌株。
