早熟矮秆高粱不育系P03A生育期和株高性状的遗传分析
2020-08-15段有厚卢峰
段有厚,卢峰
早熟矮秆高粱不育系P03A生育期和株高性状的遗传分析
段有厚,卢峰
(辽宁省农业科学院高粱研究所,沈阳 110161)
研究早熟矮秆高粱不育系P03A生育期和株高遗传效应,探讨P03A早熟矮秆性状遗传规律,为早熟矮秆性状遗传改良提供理论依据。2016年以P03A、L025A、L080A、L081和P02A为母本,分别与恢复系L242、L2381、LNK1、L280、L237和L298采用不完全双列杂交方法进行杂交组配,获得F1杂交种子,并于2016年冬天在海南加代种植,套袋自交获得F2种子。2017—2018年通过配合力分析筛选出在生育期和株高性状上一般配合力负效应大的亲本早熟矮秆高粱不育系P03A和正效应大的中晚熟高秆恢复系L237,并以P03A和L237及其杂交后代F1、F2群体为研究对象,运用主基因+多基因混合遗传模型对生育期和株高的遗传进行4个世代联合分析。P03A/L237通过相互作用表现出较短的生育期和较矮的株高,与一般配合力较强的L237杂交组配,P03A表现出缩短生育期和降低株高的能力。与其他4个不育系相比,P03A与6个恢复系组配不同F1组合生育期和株高的超高亲值都是最小,即,另4个不育系与6个恢复系组配的F1生育期更长,株高更高,进一步验证了P03A具有缩短生育期和降低株高的遗传力。通过主基因与多基因的遗传分析方法对P03A/L237 4个世代的生育期与株高进行分析研究,发现生育期和株高性状均受2对加性-显性-上位性主基因和加性-显性多基因共同控制。生育期遗传效应分析发现加性互作效应对生育期性状的影响较大,上位性效应和显性效应真实存在。生育期主基因遗传率为81.13%,多基因遗传率为10.36%,主基因+多基因决定了生育期表型变异的91.49%,环境因素决定了生育期表型变异的8.51%;通过株高遗传效应分析,发现在控制株高的2对主效基因中,第1对主基因的加性和显性作用均大于第2对主基因,控制株高性状的2对主基因以显性效应为主,株高性状存在较大的加性互作效应。主基因遗传率为84.80%,多基因遗传率为6.89%。环境方差占表型方差的比例为8.31%。明确了早熟矮秆高粱不育系P03A生育期和株高的遗传力较高,受环境因素影响较小,在后代中遗传比较稳定的特性。在今后的亲本创造和新品种选育过程中,可充分利用P03A的遗传效应和特点挖掘早熟矮秆基因,创制适宜机械化新材料和新品种,适应未来机械化品种选育和轻简栽培要求。
早熟;矮秆;高粱;遗传分析
0 引言
【研究意义】中国高粱新品种选育较长时间内多以高秆大穗型为主,株高普遍在180—220 cm,耐密性差,不抗倒伏,难以适应目前规模化、机械化生产,制约了中国高粱产业机械化规模化的进一步发展。因此,选育适于机械化栽培矮秆高粱亲本系及杂交种成为促进高粱产业发展的关键。早熟矮秆不育系的选育是组配早熟矮秆杂交种的关键,而研究高粱早熟矮秆性状遗传规律对早熟矮秆不育系的选育具有重要的指导意义。【前人研究进展】作物的农艺经济性状多属于数量性状,早期人们认为数量性状均由微效多基因控制,后来研究表明数量性状也可能由主基因和多基因共同作用。目前,国内外对高粱农艺性状遗传分析多基于杂交F1配合力分析[1-7],这些研究将控制某一性状的多基因作为整体估计总体效应。盖钧镒等[8]、Zhang等[9]、Wang等[10]提出的主基因与多基因的遗传分析方法,把控制数量性状效应大的基因作为主基因,效应小的基因作为多基因,不仅可以鉴别主基因,而且可以检测多基因效应,并估计相应的遗传参数,更精确有效地分析遗传效应。该方法在小麦[11]、棉花[12]、水稻[13]、玉米[14]、大豆[15]等主要作物中已应用。利用主基因与多基因的遗传分析方法在高粱锤度[16-17]、光合作用[18-19]、株型[20]、穗型[21-22]、叶角[23]等数量性状遗传方面已有研究。【本研究切入点】与大作物相比,中国高粱遗传研究相对滞后,遗传改良程度相对较低。早熟矮秆耐密高粱雄性不育系P03A,是在广泛引进、收集各类资源基础上,应用高粱保持系群体改良技术,经多年选育而成的早熟矮秆高粱不育系。以其为母本所选育高粱新品种辽杂37、辽夏粱1号和辽杂54已在生产上推广应用。这些杂交种具有生育期短、植株矮、株型清秀、耐密植和适应性强等特性。但利用主基因与多基因的遗传分析方法对早熟矮秆高粱的生育期与株高的研究仍鲜见报道。【拟解决的关键问题】本研究通过配合力分析,筛选一般配合力差异大的亲本,构建组合及分离群体,对早熟矮秆P03A生育期和株高的遗传效应和遗传力进行分析。明确早熟矮秆不育系P03A生育期和株高遗传效应问题及遗传规律,为不育系P03A在高粱早熟矮秆育种中的利用及早熟矮秆性状遗传改良提供理论依据。
1 材料与方法
1.1 试验材料
试验材料为辽宁省农业科学院高粱研究所自主选育的农艺性状稳定高粱试材,包括5份早熟矮秆雄性不育系:P03A、L025A、L080A、L081和P02A;6份中晚熟中高秆恢复系:L242、L2381、LNK1、L280、L237和L298。2016年在辽宁省农业科学院试验田(沈阳)以P03A、L025A、L080A、L081和P02A为母本,分别与恢复系L242、L2381、LNK1、L280、L237和L298采用不完全双列杂交(North Carolina II-incomplete diallel cross,NCII)方法进行杂交组配,获得F1杂交种子,并于2016年冬天在海南加代种植,套袋自交获得F2种子。
1.2 试验设计
2017年,在辽宁省农业科学院试验田(沈阳)种植试验亲本及其F1,将组配成的30个杂交组合按照随机区组设计种植,3次重复。经过2017年筛选分析,于2018年种植P03A和L237亲本及其杂交组合后代F1和F2群体。亲本和F1每小区种植6行,F2种植20行,行长5 m,行距0.6 m。密度为13.5万株/hm2。试验于辽宁省农业科学院试验田(沈阳)进行,田间管理同大田生产。该地区气候类型为温带半湿润大陆性气候,雨热同季,光照充足,春季风大多干旱。常年平均气温为7.2℃,全年无霜期145—160 d。试验土壤类型为壤土,前茬为大豆。0—20 cm耕层土壤有机质含量为18.63 g·kg-1、全氮0.07 g·kg-1、速效钾0.12 g·kg-1、有效磷0.03 g·kg-1,pH6.2。
1.3 性状测定与统计分析
统计出苗期、成熟期,并于成熟期测量株高,统计标准参照中国高粱区域试验:出苗期指幼苗出土“露锥”(即子叶展开前)达75%的日期;成熟期指75%以上植株的穗背阴面下部第一枝梗籽粒达蜡状硬度的日期;生育期指从出苗期到成熟期的日数。每个亲本各测量10株,F1测量10株,F2测量385株。采用EXCEL和DPS及植物数量性状混合遗传模型主基因+多基因多世代联合分析软件进行模型分析和遗传参数估计[8]。
利用ZHANG等[9]联合世代主基因多基因混合遗传模型,对P1、P2、F1、F2高粱4个世代的生育期和株高进行联合世代遗传模型分析,通过比较0、1和2对主基因的C(无主基因)、A(1对主基因)、D(1对主基因+多基因)、B(2对主基因)、E(2对主基因+多基因)共24个遗传模型的(Akaike’s Information-Criterion)值以选最优模型,并且进行遗传模型的适合性检验(均匀性检验、Smirnov检验和Kolmogorov检验),共有5个统计量:21、22、23(均匀性检验)、2(Smirnov检验)和n(Kolmogorov检验)。在选择遗传模型时,综合考虑极大对数似然函数值、值和适合性检验,并根据模型估计主基因和多基因的效应值及其方差等遗传参数。计算公式为:表型方差=家系平均数的方差;主基因方差=表型方差-分布方差(2mg=2p-2);多基因方差=表型方差-主基因方差-平均数的误差方差(2pg=2p-2mg-2e)=分布方差-平均数的误差方差(2pg=2-2e);主基因遗传率=主基因方差/表型方差(2mg=2mg/2p);多基因遗传率=多基因方差/表型方差(2pg=2pg/2p)。遗传参数包括一阶参数和二阶参数两类。根据入选模型的全部一阶、二阶分布参数极大似然估计值,估计出一阶遗传参数,即主基因的加性效应值,显性效应值和上位性效应值。通过对群体表型方差的部分估计二阶遗传参数,即性状主基因遗传力及多基因遗传力[8-9]。
2 结果
2.1 生育期和株高性状的配合力方差分析
由表1可见,生育期和株高2个性状组合间差异均达极显著水平,说明各组合间存在真实的遗传差异。母本中生育期和株高性状的配合力方差均达到显著水平,父本中生育期和株高2个性状的配合力方差均达极显著水平,母本×父本的2个性状特殊配合力方差也达极显著水平,说明母本间配合力差异显著,母本间在配合力上差异小,父本间配合力极显著,父本间在配合力上差异大。据此可进一步分析其一般配合力和特殊配合力大小。

表1 各性状的方差和配合力方差分析
PH:株高;GP:生育期;**与*分别表示差异达极显著(1%)和显著(5%)水平。下同
PH: Plant height; GP: Growth period; **and *indicating significant differences (1%) and significant (5%) levels, respectively. The same as below
2.2 各亲本生育期和株高特性的一般配合力比较
通过比较各亲本生育期和株高的一般配合力(表2),5个早熟矮秆不育系,株高和生育期一般配合力效应值趋向负向,P03A的株高与生育期一般配合力分别为-5.82和-2.61,小于其他4个不育系,P03A在株高和生育期的一般配合力效应值均为负向最大,说明其具有较好地降低株高和缩短生育期的作用。以其为母本的杂交组合,可降低F1的株高并缩短其生育期。6个父本的株高和生育期一般配合力效应值趋向正向,其中,L237株高和生育期的一般配合力效应值均为正向最大,与P03A的加性效应恰恰相反,以L237组配的F1生育期会延长,株高升高。
2.3 组合的生育期和株高特性特殊配合力分析
通过30个组合的特殊配合力分析得出P03A/L237在生育期和株高性状上均表现出最小的特殊配合力(表3)。由此可知,P03A/L237通过相互作用表现出了较短的生育期和较矮的株高,与一般配合力较强的L237杂交组配,P03A表现出了缩短生育期和降低株高的能力。
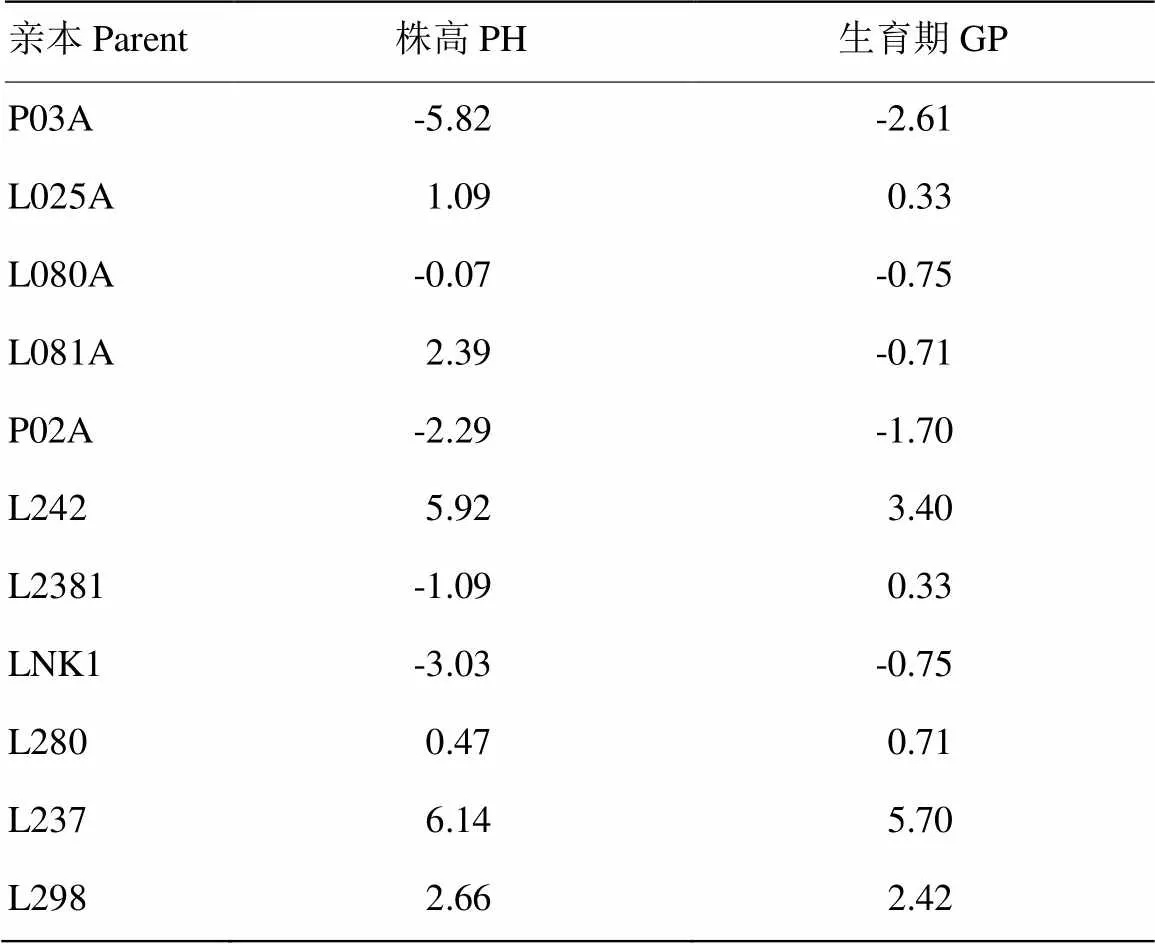
表2 各亲本的一般配合力的效应值

表3 特殊配合力极值及其组合名称
2.4 P03A为母本组配的F1组合生育期及株高杂种优势分析
由表4可知,与其他4个不育系相比,P03A与6个恢复系组配不同F1组合生育期和株高的超高亲值均是最小,即,另4个不育系与6个恢复系组配的F1生育期更长,株高更高,进一步验证了P03A具有缩短生育期和降低株高的遗传力。通过对以P03A为母本的6个杂交组合进行杂种优势分析(表5),各组合F1的生育期均比中亲值(midparent,MP)早,最少早0.5 d,最多早4 d。结果表明,由P03A组配的杂种一代生育期的表现倾向于早熟亲本,不育系P03A的生育期早熟表现为部分显性,而且其早熟显性程度受到不同遗传背景的恢复系的影响。
2.5 株高和生育期性状遗传分析
选取在株高和生育期上一般配合力负效应强的P03A与一般配合力正效应强的L237进行杂交,再利用4个世代遗传体系(P1、P2、F1和F2)应用主基因+多基因混合遗传模型对生育期和株高性状进行遗传分析。
2.5.1 P1、P2、F1和F24个世代生育期和株高频率分布 由分离世代性状统计分析与正态性检验结果(表6)可知,F2中生育期和株高性状偏度值>0,峰度值<0,偏度和峰度绝对值均<1.0,F2中生育期正态性检验<0.05,为偏正态分布,株高性状正态性检验≈0.05,为近似正态分布;生育期和株高性状表现连续的双峰和单峰分布(图1和图2),具有植物数量性状遗传的分布特征。推测可能有主基因或主基因+多基因混合遗传方式控制其生育期和株高性状。
由表7可见,P03A平均生育期天数为102 d,L237的平均生育期天数为118 d,F1平均生育期天数为106 d,F2平均生育期天数为106 d。亲本中亲值为110 d,因此,F1和F2均偏向于生育期早熟不育系。F2生育期出现5株超低亲遗传,没有超高亲遗传现象。P03A平均株高为95 cm,L237的平均株高为160 cm,F1平均株高为163 cm,F2平均株高为142 cm。亲本中亲值为128 cm,因此,F1和F2均偏向于高株恢复系。F2株高出现了60株超高亲遗传,6株超低亲遗传现象。
2.5.2 最适遗传模型的确定 利用4个世代联合分析方法对组合P03A/L237生育期和株高进行分析,获得5类24种遗传模型的极大似然函数值和值(表8)。E-1模型的值最低,为2 156.72,其次是E-2和E-0。根据盖钧镒世代模型分析方法,生育期性状选取值较小的E0、E1和E2作为备选模型;同理,株高性状选取值较小的E1、E3和E0作为备选模型。根据主基因+多基因遗传模型分析法可知,E0、E1、E2和E3属于2对主基因+多基因调控时,最适模型不一定是最小值,应根据其适合性检验结果来综合考虑利用,利用21、22、23(均匀性检验)、2(Smirnov检验)和n(Kolmogorov检验)对以上较小值模型的F2世代进行适合性检验(表9),最终选择统计量达到显著水平个数最少的模型作为最优模型。由表9得知,显著水平最少的均为E-1(MX2-ADI- AD)模型(显著水平个数均为0个),即2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型。该遗传模型可解释为性状表现受2对主基因和多基因混合遗传控制,主基因的基因作用方式表现为加性、显性、上位性作用,多基因表现为加性、显性作用。
由于试验所用恢复系的生育期、株高均比不育系长、高,故超高亲以F1与恢复系比较计算
Since the restorer lines used in the test are longer than the growth period of the sterile line and the plant height was higher, the F1was compared with the restorer line
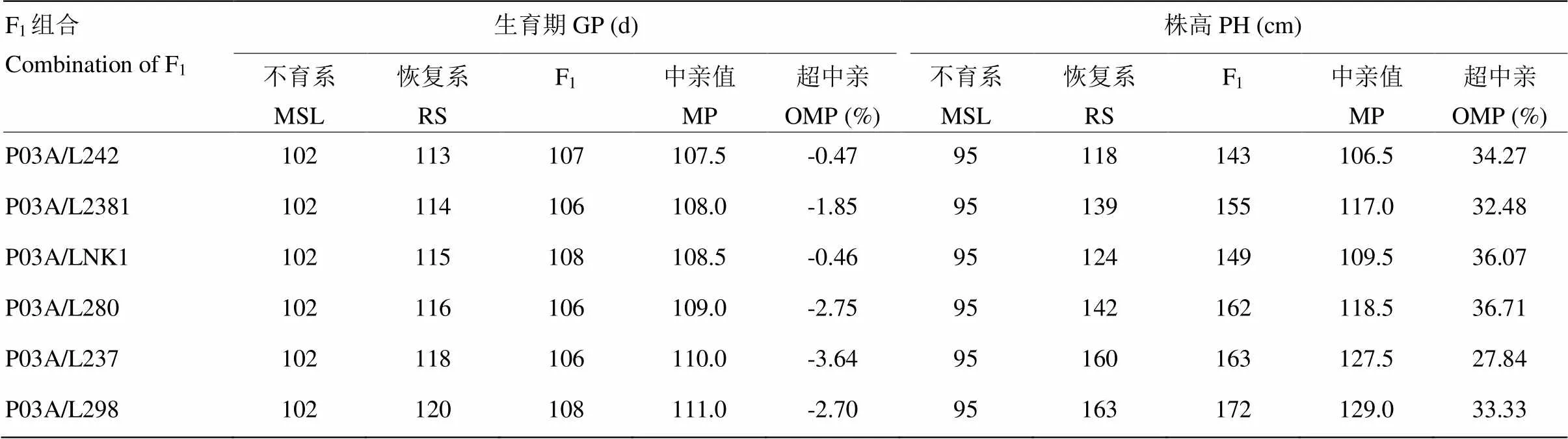
表5 P03A、不同类型(中晚熟、中高秆)恢复系及其杂交种F1的生育期和株高统计
MSL:不育系;RS:恢复系;MP:中亲值;OMP:超中亲 MSL: Male sterile line; RS: Restorer line; MP: mid-parent value; OMP: over mid-parent value
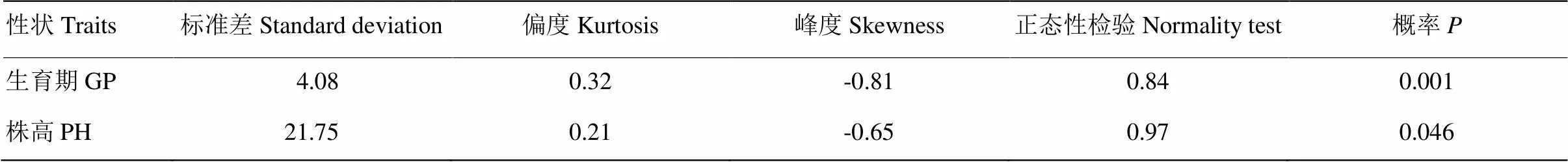
表6 F2植株生育期和株高性状统计分布和正态性检验
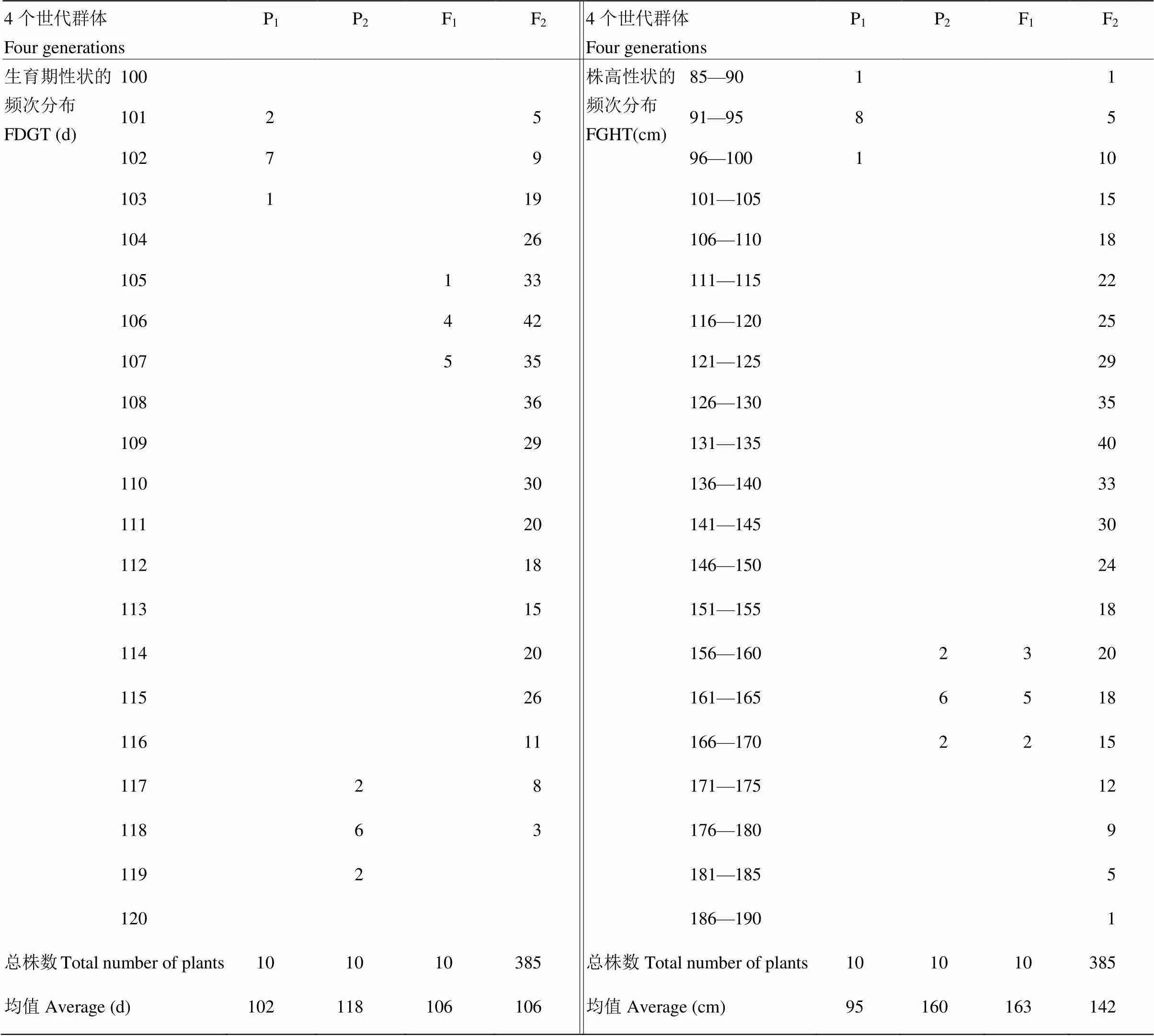
表7 P03A/L237组合P1、P2、F1和F2群体生育期与株高频次分布
FDGT:生育期性状的频次分布;FDHT:株高性状的频次分布
FDGT: Frequency distribution of growth traits; FDHT: Frequency distribution of plant height traits
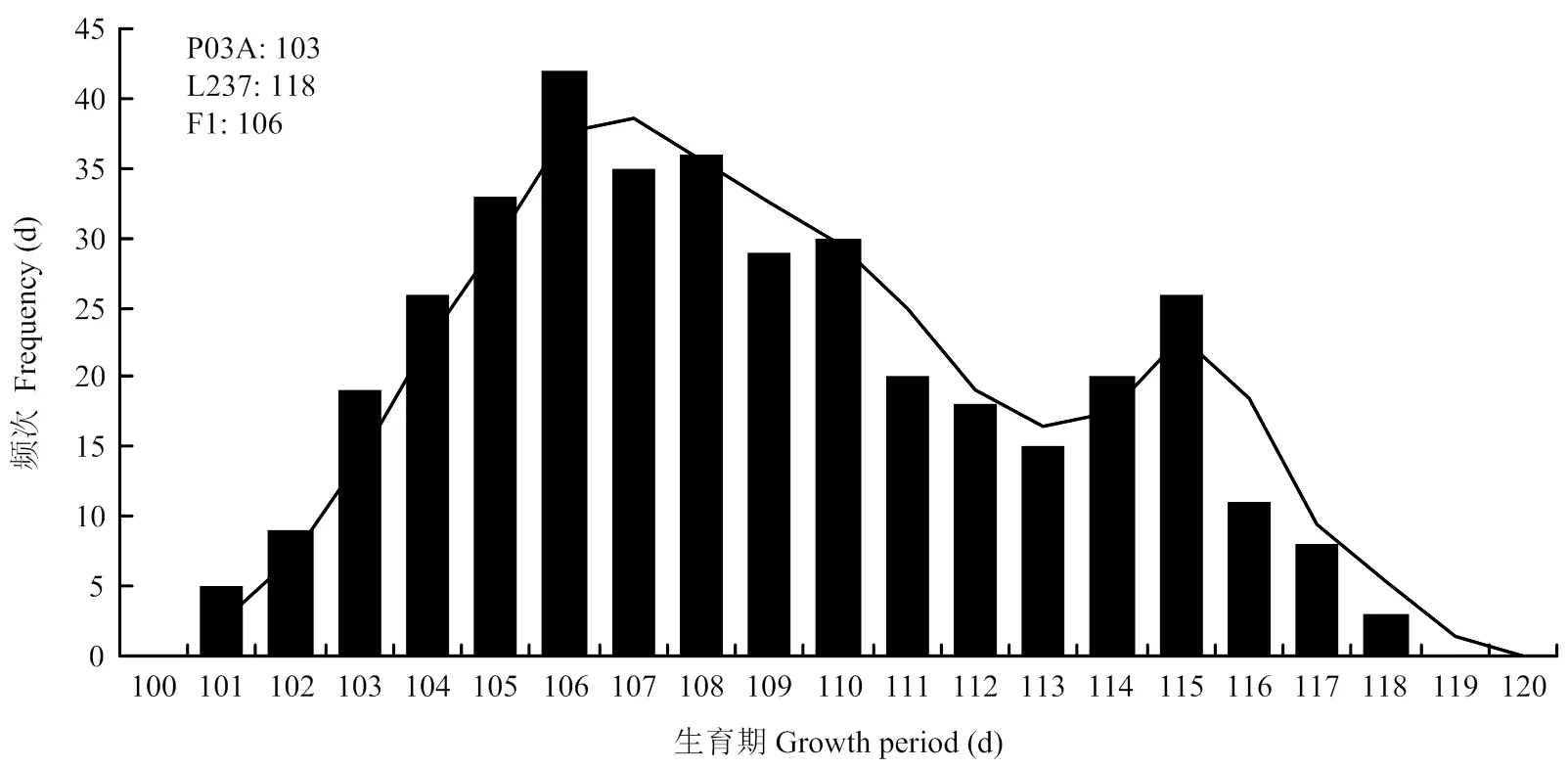
图1 P03A/L237的F2生育期株数频次分布图
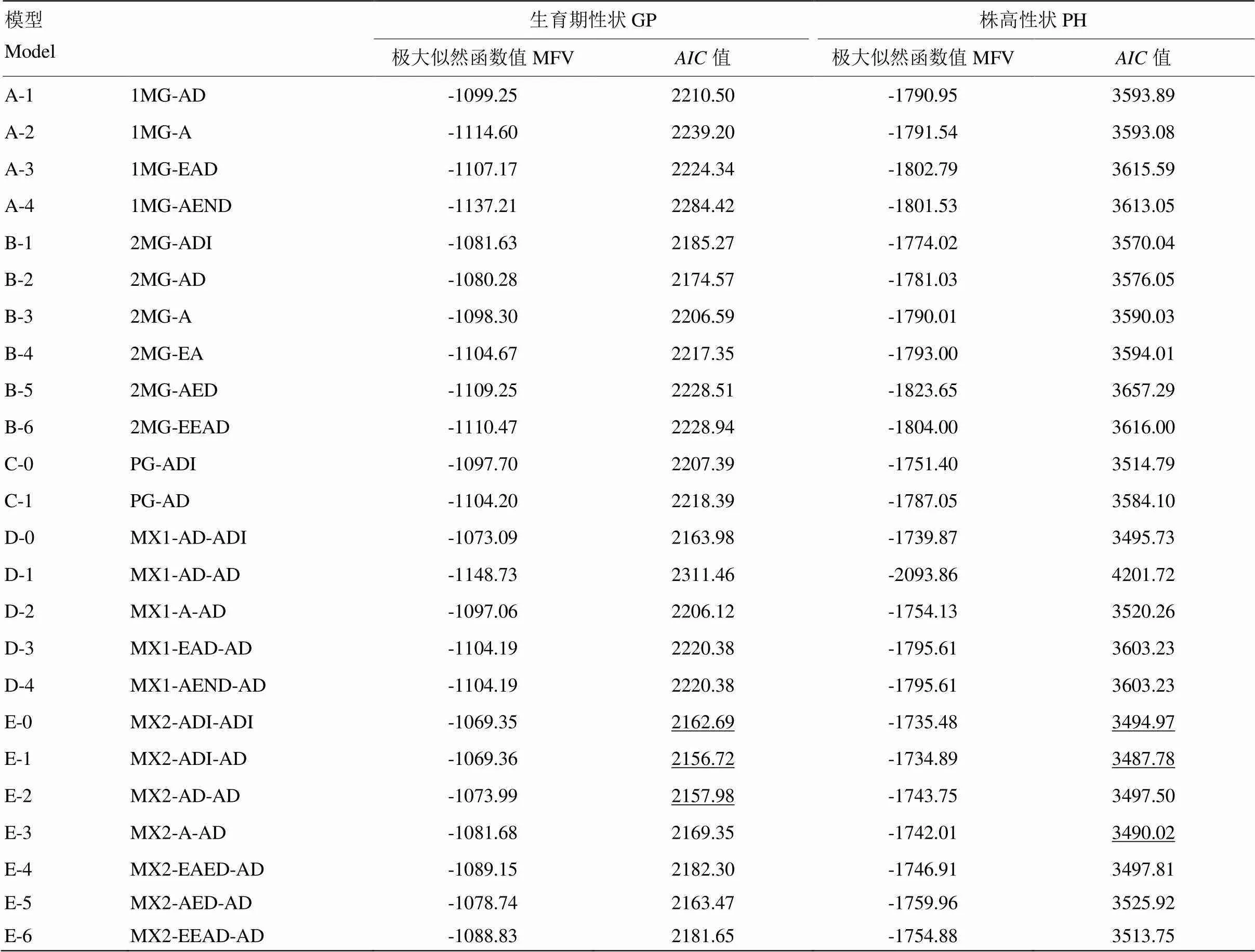
表8 P03A/L237 4个世代联合分析在不同遗传模型下的极大似然函数AIC值
MFV极大似然函数值;MG:主基因模型;MX:主基因+多基因混合模型;PG:多基因模型;A:加性效应;I:上位性效应;E:等位;C:完全;P:部分。例如:E-0模型MX2-ADI-ADI,表示2对加性-显性-上位性主基因+加性-显性-上位性多基因混合遗传模型。下划线为值较小的模型
MFV: Maximum likelihood function value; MG: major gene model; MX: mixed major gene and polygene model; PG: polygene model; A: Additive effect; I: interaction (epistasis); E: equal; C: completely; P: partly. Example: MX2-ADI-ADI represents 2 pairs of additive-dominant-epitope major gene + additive-dominant- epistatic multi- gene mixed genetic model.The model with a smallervalue is underlined
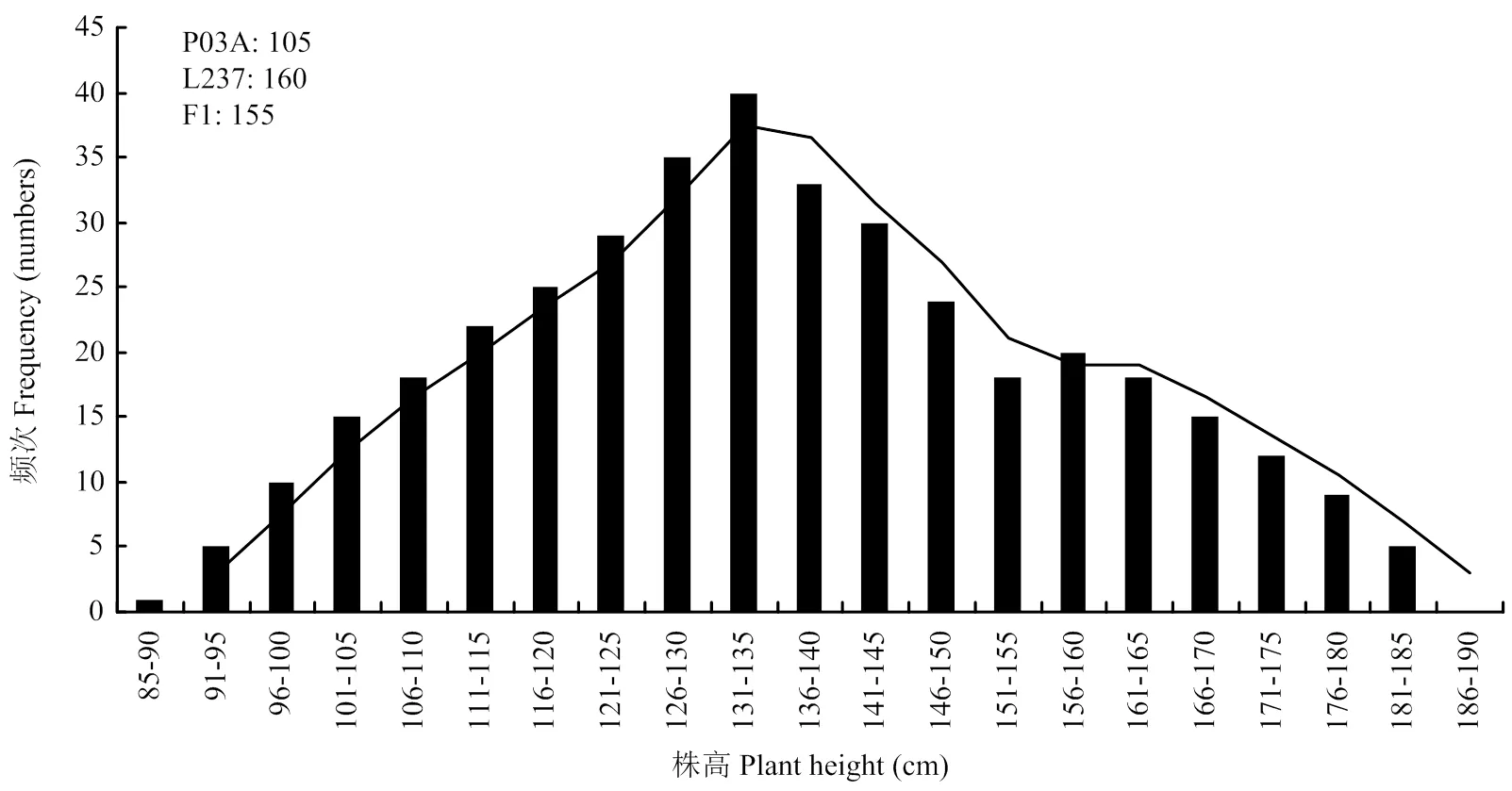
图2 P03A/L237的F2株高株数频次分布图
括号外数据是适合性检验值,括号内数据表示显著水平
The data outside the parentheses in the table is the fitness test value, and the data in parentheses indicates the significant level
2.5.3 遗传参数的估计 生育期性状:在控制生育期的2对主效基因中,第1对主基因的加性效应(a)、显性效应(a)和显性度(a/a)分别为4.5649、-0.76和-0.1682,第2对主基因的加性效应(b)、显性效应(b)和显性度(b/b)分别为-0.0286、0.6971和24.3741(表10)。其中|a|>|b|,说明第1对主基因的加性作用大于第2对主基因的加性作用;|a|>|b|,说明第1对主基因的显性作用大于第2对主基因的显性作用;二者的显性度分别小于1和大于1,第1对主基因的显性度为不完全显性,第2对主基因的显性度为超显性,说明控制生育期性状的2对主基因既有加性效应又有显性效应。从主基因间的互作分析结果来看,2对主基因加性×加性和显性×显性的互作效应绝对值均大于主基因显性效应,加性×加性的互作效应大于显性×显性的互作效应,说明加性互作效应对生育期性状的影响比较大。多基因加性效应值[]为-12.1463,多基因显性效应值[]为-4.0979。从生育期性状的二阶参数分析结果可知,主基因遗传率为81.13%,多基因遗传率为10.36%。环境方差占表型方差的比例为8.51%。
株高性状:在控制株高的2对主效基因中,第1对主基因的加性效应(a)、显性效应(a)和显性度(a/a)分别为13.8071、-14.6771和-1.0630,第2对主基因的加性效应(b)、显性效应(b)和显性度(b/b)分别为-1.2563、-13.136和10.4562(表10)。其中|a|>|b|,说明第1对主基因的加性作用大于第2对主基因的加性作用;|a|>|b|,说明第1对主基因的显性作用大于第2对主基因的显性作用;二者的显性度均大于1,说明控制株高性状的2对主基因以显性效应为主。从主基因间的互作分析结果来看,2对主基因加性×加性及显性×显性的互作效应绝对值均大于主基因加性效应和显性效应;加性×加性的互作效应大于显性×显性的互作效应,说明加性互作效应对株高性状的影响比较大。多基因加性效应值[]为-46.5507,多基因显性效应值[]为68.7717。从株高性状的二阶参数分析结果可知,主基因遗传率为84.80%,多基因遗传率为6.89%。主基因遗传率远大于多基因遗传率。环境方差占表型方差的比例为8.31%。
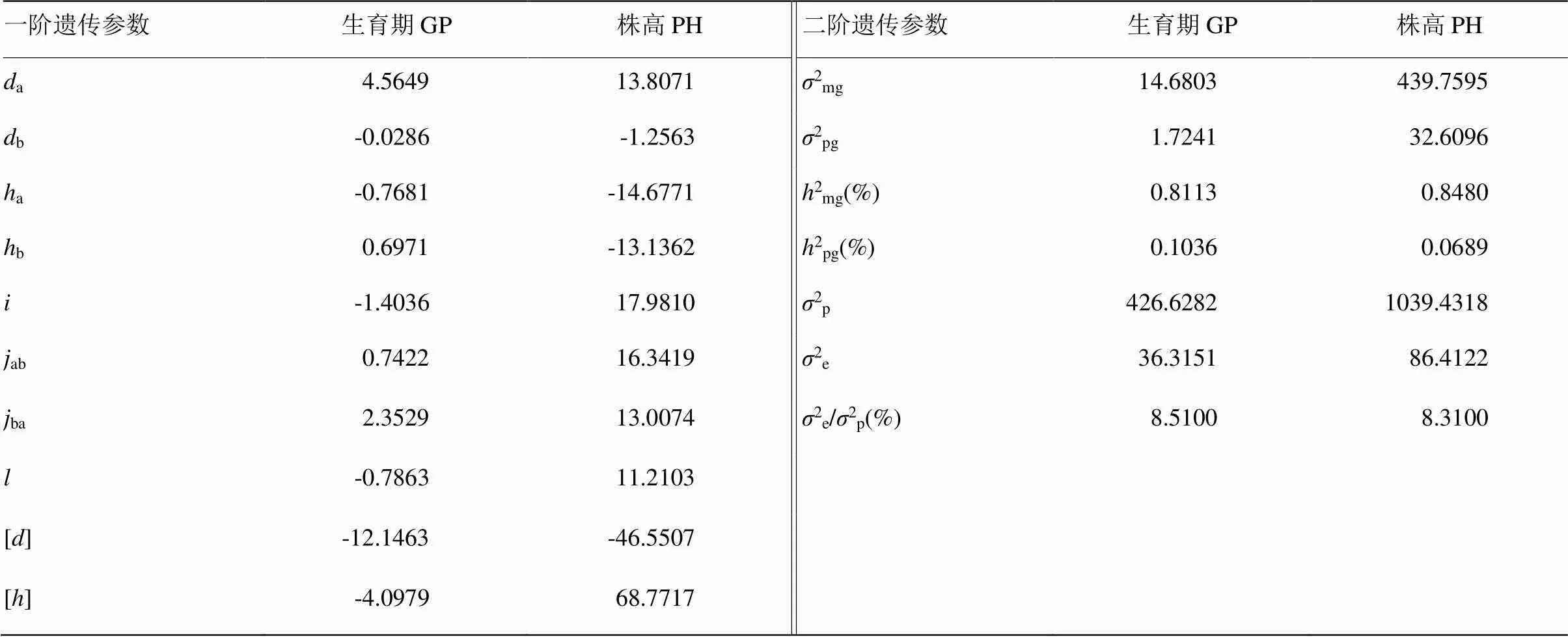
表10 生育期和株高性状E-1模型遗传参数的估计
a:主基因A的加性效应;b:主基因B的加性效应;a:主基因A的显性效应;b:主基因B的显性效应;:2个主基因的加×加效应;:2个主基因的显×显效应;ab:加性(a)显性(b)效应;ba:加性(b)×显性(a)效应;[]:多基因加性效应;[]:多基因显性效应;2mg:主基因方差;2pg:多基因方差;2mg(%):主基因遗传率;2pg(%):多基因遗传率;2p:表型方差;2e:环境方差
a: Additive effect of major gene A;b: Additive effect of major gene B;a: Dominant effect of major gene A;b: Dominant effect of major gene B;: Additive plus effect of two major genes;: Significant effect of two major genes;ab: Additive(a) ×dominant(b);ba: Additive(b) ×dominant(a); []: Additive effect of multi-gene; []: Dominant effect of multi-gene;2mg: Variance of major gene;2pg: Variance of multi-gene;2mg(%): Heritability of major gene;2pg(%): Heritability of multi-gene;2p: Variance of phenotype;2e: Variance of environment
3 讨论
本研究发现各组合F1株高均表现出较强的超中亲优势,结果表明,P03A组配的高粱杂交组合株高的遗传存在着杂种优势现象,在杂种一代中高秆对矮秆表现为显性或部分显性作用,因遗传背景不同而略有差异。利用最佳杂交亲本是杂交种组配成功的关键和基础[24],相比较而言,在生育期和株高方面,P03A/L237杂交组合表现出最小的超中亲优势,该组合是本试验需选择的最佳组合。
本研究利用P03A和L237构建P1、P2、F1和F24个世代群体,应用植物数量性状主基因+多基因混合遗传模型,对P03A生育期和株高进行了遗传分析研究,明确了基于早熟矮秆高粱P03A/L237杂交F2生育期和株高的最适遗传模型均为E-1模型,即2对加性-显性-上位性主基因+加性-显性多基因遗传模型。本试验在配合力分析的基础上选取了配合力差异大的P03A和L237,并连续世代分析遗传效应,克服了纯粹单个分离世代分析的局限性,分解出多基因的变异,并将单个分离世代及亲本、F1和F2纳入了统一的似然函数,从而综合地估计出遗传参数,减小了环境对试验结果的误差影响。
3.1 生育期性状的遗传效应
本研究表明,杂交F2生育期多数单株倾向早熟亲本,控制生育期的2对主效基因中,第1对主基因的加性作用大于第2对主基因的加性作用,第1对主基因的显性作用大于第2对主基因的显性作用。加性×加性的互作效应大于显性×显性的互作效应,说明加性互作效应对生育期性状的影响较大,通过上位性效应及多基因效应数值分析,上位性效应和多基因效应真实存在。主基因遗传率远大于多基因遗传率,主基因+多基因决定了生育期表型变异的91.49%,环境因素决定了生育期表型变异8.51%,生育期性状遗传表现主要受遗传因素的影响,受环境影响较小。这与杨伟光[25-26]和李振武[27]结果相近,虽然此研究较早,分析的世代亦有所不同(有F1、F2和F3),但研究结果表明高粱生育期遗传均存在加性效应、显性效应和上位性效应。F2生育期存在超低亲遗传现象,育种者可利用高粱早熟基因遗传力高的特点,可在杂交后代中进行早期选择偏早熟的后代。
3.2 植株性状的遗传效应
本研究表明,在控制株高的2对主效基因中,第1对主基因的加性作用大于第2对主基因的加性作用,第1对主基因的显性作用大于第2对主基因的显性作用,控制株高性状的2对主基因以显性效应为主,加性互作效应对株高性状的影响较大,通过上位性效应分析,上位性效应也真实存在。主基因遗传率远大于多基因遗传率,说明株高性状主要受2对主基因的作用。环境方差占表型方差的比例为8.31%,株高性状遗传表现主要受遗传因素的影响,受环境影响较小。这与杨伟光等[28-29]结果一致,与管延安等[17](株高F2遗传符合全显性主基因+加性-显性多基因混合遗传模型)和白晓倩等[30](株高F2遗传符合加性-显性-上位性的混合遗传模型)相近,但不一致,或许与试验材料的选取有关。
分析研究早熟矮秆高粱的遗传对于高粱亲本选育和品种改良具有重要意义。在原有优质高产的品种基础上,提早生育期可满足不同生态条件和生产季节的需要,有助于解决早熟与丰产难以兼顾的矛盾;目前在中国高粱生产中应用的A2型雄性不育系多是印度高粱或偏印度高粱。印度高粱与中国高粱杂交优势较强,株高优势也强,这不利于抗倒和机械收割。所以,在培育不育系时还要注意株高优势弱的材料选拔[31]。矮秆材料普遍存在抗倒伏能力强,适宜机械化栽培管理的优势,所以要重视矮秆性状的遗传研究。
4 结论
明确了P03A生育期和株高的遗传力较高、受环境因素影响较小、在后代中遗传比较稳定的特性。在今后的亲本创造和新品种选育过程中,可充分利用P03A遗传效应和特点挖掘早熟矮秆基因,创制适宜机械化新材料和新品种,适应未来机械化轻简栽培要求。
[1] 张晓娟, 张一中, 周福平. 高粱新选不育系主要农艺经济性状的配合力分析. 中国农学通报, 2012, 28(18): 71-75.
ZHANG X J, ZHANG Y Z, ZHOU F P. Analysis on the combining ability of main agronomic and economic traits for new sorghum male sterility lines., 2012, 28(18): 71-75. (in Chinese)
[2] 尹学伟, 王培华, 张晓春. 14个糯高粱亲本主要农艺性状配合力及遗传力分析. 西南农业学报, 2014, 27(4): 1363-1367.
YIN X W, WANG P H, ZHANG X C. Analysis of 14 parents glutinous sorghum’s main agronomic characteristics combining ability and heritability., 2014, 27(4): 1363-1367. (in Chinese)
[3] 吕鑫, 平俊爱, 张福耀, 杜志宏, 李慧明, 杨婷婷, 牛皓, 姚琳. 新选饲草高粱恢复系农艺性状配合力效应分析. 草业科学, 2016, 33(7): 1361-1366.
Lü X, PING J A, ZHANG F Y, DU Z H, LI H M, YANG T T, NIU H, YAO L. Effect analysis on the combining ability of main agronomic traits for new breeding restorer lines derived from forage sorghum., 2016, 33(7): 1361-1366. (in Chinese)
[4] 李金梅, 赵威军, 张福耀. 甜高粱抗倒伏性相关性状的配合力和遗传参数分析. 作物杂志, 2014(2): 56-60.
LI J M, ZHAO W J, ZHANG F Y. Analysis on combining ability and genetic parameters of traits related to lodging resistance in sweet sorghum., 2014(2): 56-60. (in Chinese)
[5] 高海燕, 程庆军, 田承华. 新选高粱亲本系的配合力及遗传力分析. 农学学报, 2016, 6(5): 6-10.
GAO H Y, CHENG Q J, TIAN C H. Combining ability and heritability of new sorghum parental lines., 2016, 6(5): 6-10. (in Chinese)
[6] NI X L, ZHAO G L, LIU T P. Analysis on the combining ability and heritability of main agronomic traits of hybrid glutinous sorghum., 2012, 13(10): 2104-2109.
[7] Geleta N, Mohammed H, Zelleke H. Genetic variability, heritability and genetic advance in sorghum [(L.) Moench] germplasm., 2005, 30(3): 439-445.
[8] 盖钧镒, 章元明, 王建康. 植物数量性状遗传体系. 北京: 科学出版社, 2003.
GAI J Y, ZHANG Y M, WANG J K.. Beijing: Science Press, 2003. (in Chinese)
[9] ZHANG Y M, Gai J Y, YANG Y. The ElM algorithm in the joint segregation analysis of quantitative traits., 2003, 81(2): 157-163.
[10] WANG J, Fodlieh D W, Cooper M, Delacy I H. Power of the joint segregation analysis method for testing mixed major-gene and polygene inheritance models of quantitative traits., 2001, 103: 804-816.
[11] 温明星, 李东升, 胡芳芳. 宁麦9号×镇麦168小麦F2群体产量相关性状的遗传模型分析. 麦类作物学报, 2018, 38(4): 386-394.
WEN M X, LI D S, HU F F. Genetic model analysis on yield-related traits in wheat f2population of Ningmai 9×Zhenmai 168., 2018, 38(4): 386-394. (in Chinese)
[12] 赵树琪, 庞朝友, 魏恒玲, 王寒涛, 李黎贝, 宿俊吉, 范术丽, 喻树迅. 陆地棉早熟性状多世代联合遗传分析. 棉花学报, 2017, 29(2): 119-127.
ZHAO S Q, PANG C Y, WEI H L, WANG H T, LI L B, SU J J, FAN S L, YU S X. Genetic inheritance of earliness traits in upland cotton (L.) inferredby joint analysis of multiple generations., 2017, 29(2): 119-127. (in Chinese)
[13] 刘金波, 徐波, 李建红, 李健, 刘艳, 周振玲, 杨波, 迟铭, 宋兆强, 卢百关, 方兆伟. 水稻株高和每穗颖花数的6个世代联合遗传分析. 华北农学报, 2017, 32(S1): 88-94.
LIU J B, XU B, LI J H, LI J, LIU Y, ZHOU Z L, YANG B, CHI M, SONG Z Q, LU B G, FANG Z W. Joint genetic analysis on plant height and spikelets per panicle by using six generations of two crosses inrice.,2017, 32(S1): 88-94. (in Chinese)
[14] 进茜宁, 张怀胜, 王铁固, 吴向远, 陈士林. 玉米单穗粒质量的遗传模型分析. 河南科技学院学报(自然科学版), 2018, 46(5): 17-21.
JIN Q N, ZHANG H S, WANG T G, WU X Y, CHEN S L. Genetic model analysis of maize single spike kernel weight.(), 2018, 46(5): 17-21. (in Chinese)
[15] 赵桂云, 王继安, 李文滨, 滕卫丽, 韩英鹏. 大豆抗食心虫主基因+多基因混合遗传模型的五世代联合分析. 大豆科学, 2014, 33(3): 301-304.
ZHAO G Y, WANG J A, LI W B, TENG W L, HAN Y P. Genetic analysis on resistance to soybean pod borer by using five generations joint analysis of mixed inheritance model of major gene and polygene., 2014, 33(3): 301-304. (in Chinese)
[16] 卢峰, 邹剑秋, 段有厚. 甜高粱茎秆含糖量相关性状的遗传分析. 中国农业大学学报, 2012, 17(6): 111-116.
LU F, ZOU J Q, DUAN Y H. Genetic analysis of stalk sugar content related traits in sweet sorghum (L. Moench)., 2012, 17(6): 111-116. (in Chinese)
[17] 管延安, 张华文, 樊庆琪, 杨延兵. 普通高粱与甜高粱杂交组合株高、糖度的主基因多基因模型遗传效应分析. 核农学报, 2012, 26(1): 36-42.
GUAN Y A, ZHANG H W, FAN Q Q, YANG Y B. Genetic analysis of plant height and brix value by using major gene and polygene inheritance model in across between common sorghum and sweet sorghum.s, 2012, 26(1): 36-42. (in Chinese)
[18] 卢华雨, 李延玲, 罗峰. 粒用高粱4个主要光合性状数量遗传分析. 江苏农业科学, 2018, 46(17): 68-72.
LU H Y, LI Y L, LUO F. Quantitative genetic analysis of four main photosynthetic traits in grain sorghum., 2018, 46(17): 68-72. (in Chinese)
[19] Fernandez M G S, Strand K, Hamblin M T,Mark W, Emily H, Stephen K. Genetic analysis and phenotypic characterization of leaf photosynthetic capacity in a sorghum (spp.) diversity panel., 2015, 62(6): 939-950.
[20] 李延玲, 白晓倩, 于澎湃, 高建明, 裴忠有, 罗峰, 孙守钧. 高粱株型性状数量遗传分析. 华北农学报, 2018, 33(1): 143-149.
LI Y L, BAI X Q, YU P P, GAO J M, PEI Z Y, LUO F, SUN S J. Quantitative genetic analysis of sorghum plant type characters., 2018, 33(1): 143-149.(in Chinese)
[21] 卢华雨, 白晓倩, 于澎湃, 罗峰. 饲用高粱4个主要株型性状的遗传分析. 贵州农业科学, 2019, 47(1): 5-9, 13.
Lu H Y, BAI X Q, YU P P, LUO F. Genetic analysis of four main plant type traits in forage sorghum.,019, 47(1): 5-9, 13. (in Chinese)
[22] 邵健丰, 翟国伟, 王华. 高粱穗型相关性状的遗传研究. 科技通报, 2019, 35(2): 46-48.
SHAO J F, ZHAI G W, WANG H. Study on genetic feature of sorghum panicle type traits., 2019, 35(2): 46-48. (in Chinese)
[23] 周紫阳, 赵雪梅, 李光华, 石贵山, 王江红, 马英慧. 高粱叶角遗传研究. 杂粮作物, 2006(6): 392-394.
ZHOU Z Y, ZHAO X M, LI G H, SHI G S, WANG J H, MA Y H. Studies on the leaf angle of sorghum.,2006(6): 392-394. (in Chinese)
[24] 卢庆善. 高粱杂交种亲本遗传多样性及其改良. 园艺与种苗, 2012(1): 1-4, 27.
Lu Q S. Genetic diversity and improvement of hybrid parents in sorghum.,2012(1): 1-4, 27. (in Chinese)
[25] 杨伟光. 高粱生育期的遗传分析. 中国农业科学, 1989(5): 19-24.
YANG W G. Genetic Analysis of Growth period of Sorghum., 1989(5): 19-24. (in Chinese)
[26] 杨伟光. 高粱杂交二代生育期遗传特性的研究. 吉林农业科学, 1989(4): 39-43.
YANG W G. Study on the hereditary character of growth period in sorghum F2generation the second of hybrid., 1989(4): 39-43. (in Chinese)
[27] 李振武. 高粱F3生育期遗传表现. 辽宁农业科学, 1984(4): 1-4.
LI Z W. Reproductive genetic performance of sorghum F3., 1984(4): 1-4. (in Chinese)
[28] 杨伟光. 高粱主要农艺性状基因效应的研究. 中国农业科学, 1991(4): 26-31.
YANG W G. Study on gene effect of major agronomic characters in sorghum.,1991(4): 26-31. (in Chinese)
[29] 杨伟光, 顾德峰, 牟金明. 中国高粱地方品种株高的遗传研究. 吉林农业大学学报, 1993(4): 28-31, 105-106.
YANG W G, GU D F, MU J M. Genetic study on plant height of local variety of chinese sorghum., 1993(4): 28-31, 105-106. (in Chinese)
[30] 白晓倩, 于澎湃, 李延玲, 高建明, 裴忠有, 罗峰, 孙守钧. 粒用高粱F2群体农艺性状数量遗传分析. 华北农学报, 2019, 34(1): 107-114.
BAI X Q, YU P P, LI Y L, GAO J M, PEI Z Y, LUO F, SUN S Y. Genetic analysis of agronomic characters in F2population of sorghum bicolor.,2019, 34(1): 107-114. (in Chinese)
[31] 高士杰, 陈冰嬬, 李继洪, 贾俊英, 侯玉波. 中国高粱春播早熟区雄性不育系存在的问题探讨. 吉林农业科学, 2012, 37(5): 9-11.
GAO S J, CHEN B X, LI J H, JIA J Y, HOU Y B. Discussions on problems in male sterile line of sorghum in spring seeding early-maturing region in China., 2012, 37(5): 9-11. (in Chinese)
Genetic Analysis on Growth Period and Plant Height Traits of Early-maturing Dwarf Sorghum Male-sterile Line P03A
DUAN Youhou, LU Feng
(Sorghum Research Institute, Liaoning Academy of Agricultural Sciences, Shenyang 110161)
【】In order to identify the genetic effect of early-maturing and dwarf traits on sorghum male-sterile line P03A, here we, provide a theoretical basis for sorghum early-maturing and dwarf breeding improvement by genetic analysis on growth period and plant height. 【】In 2016, sorghum male-sterile line P03A,L025A, L080A, L081 and P02A were used as female parent lines, and restoring line L242, L2381, LNK1, L280, L237, and L298 were used as male parent lines the F1hybrid seeds were obtained by NCⅡ crossing method. And in the winter of 2016, F2seeds were harvested from F1selfing plantlets in Hainan province. During 2017-2018, Combining ability analysis on growth period and plant height traits were performed, together with 4 generation conjoint analysis from hybrid F1and F2population using mixed major gene plus poly-gene inheritance model. P03A and L237 were selected as female and male parent respectively. 【】Hybrid combination P03A/L237 show the characters of short growth period and short plant height through interaction of the 2 parent lines. P03A contribute to shortening the growth period and plant height in hybrid. Compared with the rest 4 sterile lines, the hybrid combinations with P03A show shorter growth period and plant height, and heritability on the 2 traits was identified. 4-generation analysis of P03A/L237 on growth period and plant height were performed using major gene plus poly-gene inheritance model, which indicate that growth period and plant height traits were both controlled by two major genes with additive-dominate-epistatic effects and poly-genes. Analysis result of growth period suggest that additive effect is higher than epistatic and dominate effects. Heritability of major genes is 81.13%, and heritability of poly-genes is 10.36% respectively. 91.49% phenotypic variation is conducted by major genes plus poly-genes, and 8.51% phenotypic variation is conducted by environmental factors. Analysis of plant height indicate that additive effects and dominant effects of the first major gene are all stronger than the second major gene, and the dominant effect is much more important. Heritability of major genes and poly-genes is 84.80%, and 6.89% respectively. 8.31% phenotypic variation is conducted by environmental factors. 【】Genetic effects of growth period and plant height of sorghum sterile line P03A were analyzed in this study. It was identified that the heritability of the two traits mentioned above are relatively high, not easily affected by environmental factors, and with stable hereditary characters. Thus, P03A can be utilized due to its early maturing and dwarf genes in sorghum hybrid breeding, which can meet the requirement of sorghum mechanized production.
early maturity; dwarf; sorghum; genetic analysis

10.3864/j.issn.0578-1752.2020.14.007
2019-07-31;
2019-10-24
国家谷子高粱产业技术体系项目(CARS-06-13.5-A11,CARS-06-13.5-A22)、中央引导地方项目(2018416023)
段有厚,E-mail:duanyouhou@163.com。通信作者卢峰,E-mail:lufeng740202023@163.com
(责任编辑 李莉)
