造林密度对马尾松优良家系人工林生长及直径分布的影响
2020-08-08颜培栋零天旺杨章旗
陈 旋,颜培栋,零天旺,梁 机,杨章旗
(1.广西大学林学院,广西南宁 530004;2.广西壮族自治区林业科学研究院,广西南宁 530002;3.横县镇龙林场,广西横县 530327)
森林是自然界重要的碳源、基因库、资源库、蓄水库和能源库[1]。随着人口急剧增长,全球森林每年净损失量为3.3×106hm2[2]。第九次全国森林资源清查显示中国人工林保存面积为8.0×107hm2,居世界首位,但中、幼林面积所占比例为63.94%,结构严重失衡,人均森林面积和蓄积少,而中国每年的木材消耗量已达到8 亿m3,对进口的依赖度超过55%,大规格木材供需矛盾尤为突出。随着全球生态环境恶化,各国严管木材出口,加快木材的生产和调整木材结构是解决国家木材安全问题的重要任务[3]。通过对人工林的立地控制、遗传控制和密度控制实现人工林速生、稳定和定向培育,是提升人工林质量,保障国家木材战略储备的重要措施[4]。其中,林分密度是最易控制的因素。密度控制包括造林密度及密度调控,两者直接关系到培育目标的实现和经营者的经济收入,密度调控是人工林定向培育的关键问题和研究热点[5]。学者们对杂种落叶松(Hybrid Larch)[6]、杉木(Cunninghamia lanceolata)[7]和华山松(Pinus armandii)[8]等树种开展了相关研究。
马尾松(Pinus massoniana)分布范围广,林分面积大,蓄积量高,是中国特有的乡土树种。其具有耐干旱瘠薄、生长迅速、适应力强和经济价值高的特点,是我国长江流域以南广袤酸性土壤荒山造林的先锋树种和主要用材树种[9-10]。许多学者对马尾松开展了密度控制与林分直径结构的研究[11-13],但结果多源自临时样地或幼林阶段,缺乏长期连续的观测数据,林分长期生长的密度控制理论和大径材定向培育技术需继续完善。本研究基于22年生不同造林密度马尾松优良家系人工林的多年观测数据,分析造林密度对林分生长及其直径结构的影响,为马尾松优良家系人工林定向培育提供理论依据。
1 材料与方法
1.1 试验地概况
试验地位于广西壮族自治区横县镇龙林场(109°08'~109°19'E,23°02'~23°08'N),海拔450 m左右,多低山丘陵;属南亚热带季风气候,年均气温21.5 ℃,最低气温-1 ℃,最高气温39.2 ℃,年均降水量1 477.8 mm,年均蒸发量1 056.9 mm,年均日照时长1 758.9 h。造林地为杉木采伐迹地,土壤为细石英岩和泥质粉砂岩发育而成的红壤,土层厚度大于80 cm,坡度25°~33°,坡向西南,立地指数18。
1.2 试验材料及试验设计
于1997年5月造林,苗木为广西壮族自治区林业科学研究院提供的马尾松优良家系(桂MVF443)Ⅰ级营养杯苗(苗高20 cm 左右),采用块状整地,整地规格40 cm×30 cm×30 cm。试验采用随机区组设计,设置4 个不同的造林密度,A:2 500 株/hm2(2 m×2 m),B:3 300 株/hm2(1.5 m×2 m),C:4 500 株/hm2(1.5 m×1.5 m),D:6 000 株/hm2(1 m×1.67 m),3 次重复,共12 个小区,小区间设置2 列保护行,小区面积400 m2,各小区集中排布,立地条件基本相同。试验林未进行间伐,无人为干扰。
1.3 指标测定
自2006年(10年生)起,每年对林木进行每木检尺,将10~22年生连年观测数据进行统计整理并计算分析。其中林分平均胸径和树高为算术平均值;单株材积(V单)采用广西马尾松单株材积经验式计算,公式为V单= 0.714 265 437×10-4×Dg1.867008×H0.9014632,式中,Dg为林分平均断面积对应的直径,H为树高;将林木胸径按2 cm 为一个径阶划分并统计各径阶的林木株数,计算直径株数分布率和累积频率分布百分比。采用偏度(SK)、峰度(ST)及变异系数(CV)对林分直径分布曲线的形状和均质性进行分析,探索密度对林分直径结构的影响。偏度、峰度及变异系数的计算公式如下:
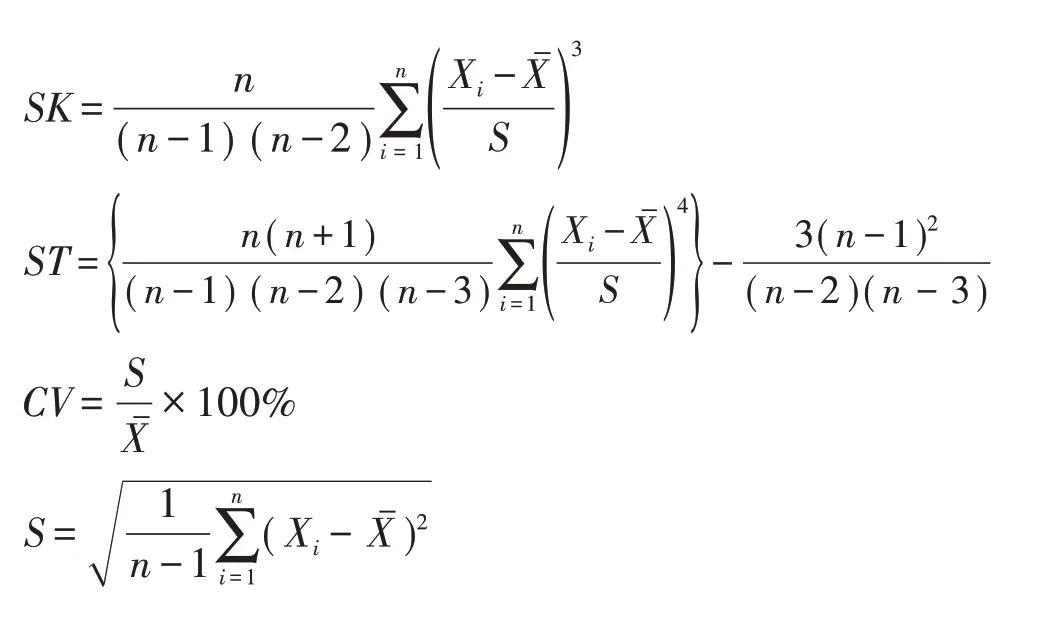
式中,n为林木株数,Xi为每木直径,Xˉ为林分平均直径,S为标准差。
1.4 数据处理
利用Excel 2010 软件整理制表,采用OriginPro 2017 作图,运用SPSS 23.0 中的One-way ANOVA 进行单因素方差分析、Duncan’s法进行多重比较。
2 结果与分析
2.1 造林密度对林分树高的影响
林分树高随林龄增大而增加,树高年均生长量则随林龄增大逐渐减小;造林密度越小,树高及树高年均生长量越大(图1)。方差分析表明,各造林密度间的树高差异不显著(表1)。
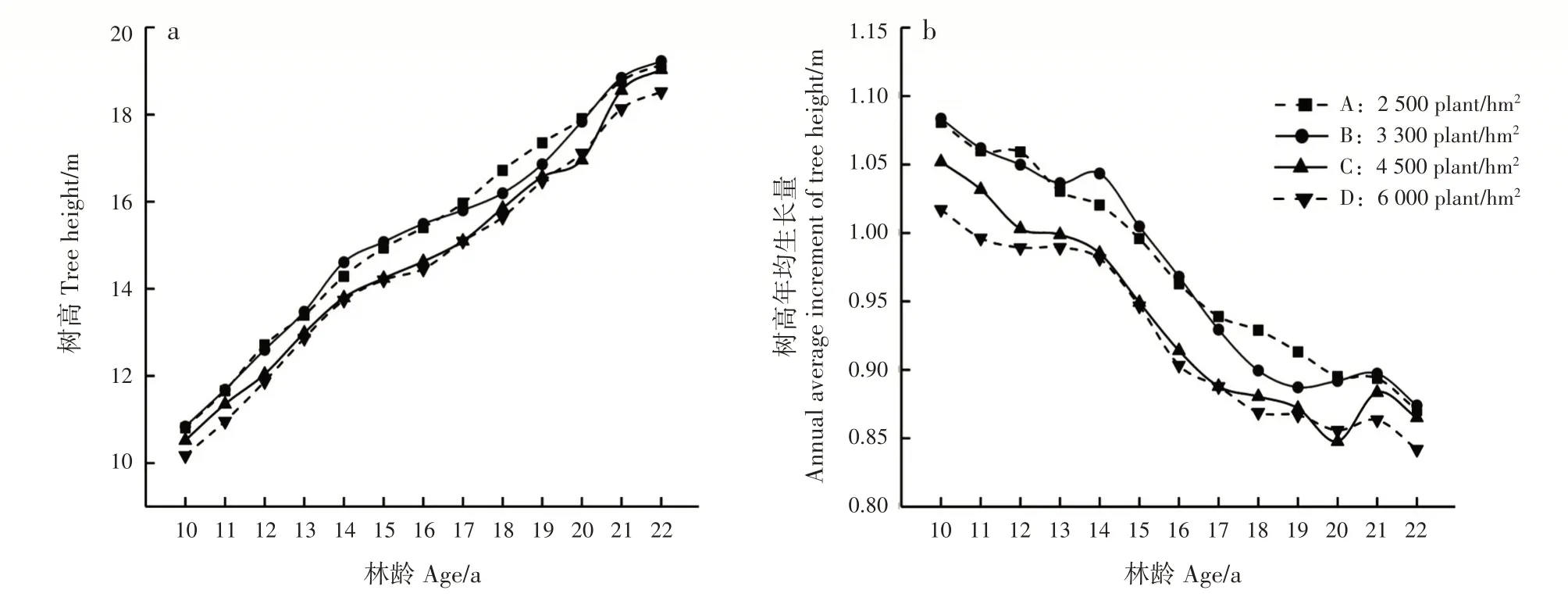
图1 不同造林密度马尾松林分不同林龄树高的变化Fig.1 Variation of tree height of P.massoniana stands with different ages in different planting densities
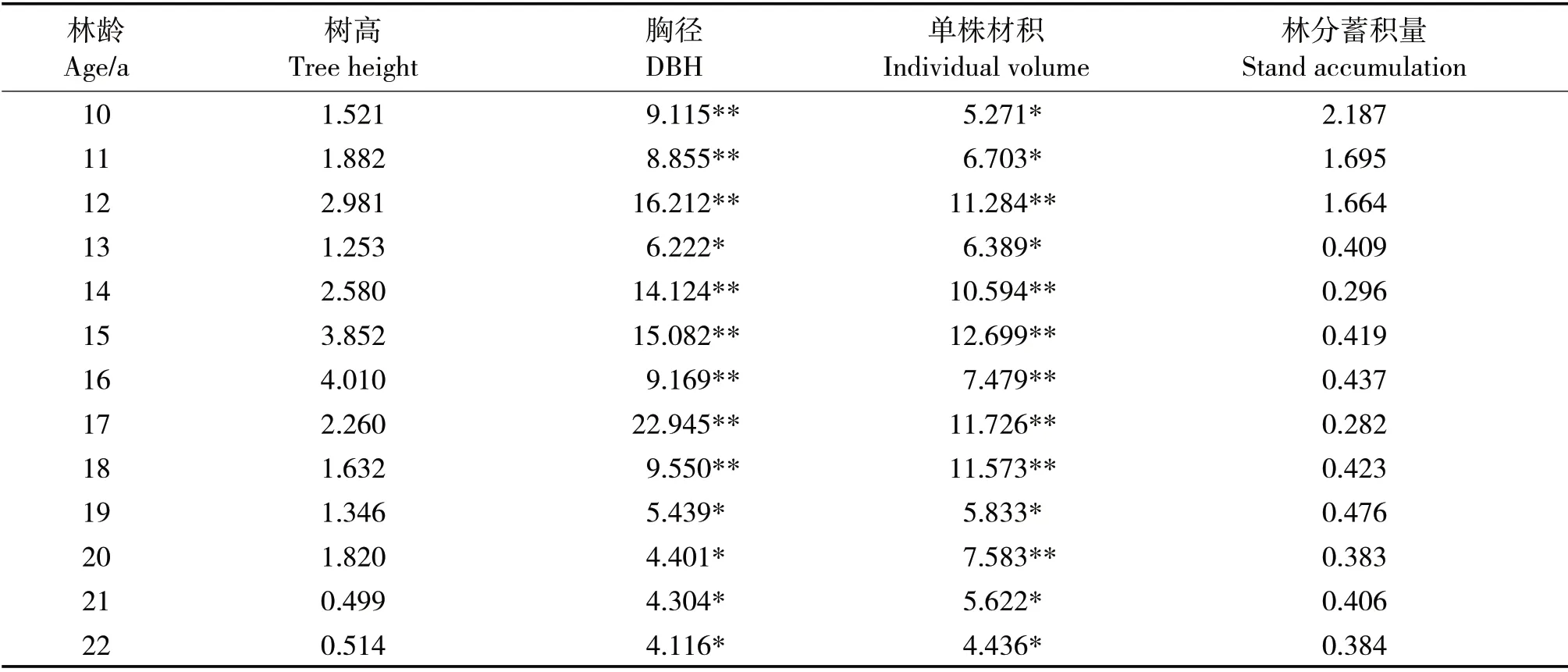
表1 不同造林密度树高、胸径、单株材积和林分蓄积量方差分析Tab.1 ANOVA of tree height,DBH,individual volume and stand accumulation in different planting densities
2.2 造林密度对林分胸径的影响
林分胸径及胸径年均生长量随造林密度增大而减小,胸径随林龄增加逐渐增大,不同造林密度的胸径年均生长量分别在21、19、16 和15年生时降至最低,而后缓慢增加(图2)。方差分析表明,13 和19~22年生时,各造林密度间胸径差异显著(P<0.05),其他林龄差异极显著(P<0.01);10~18年生时,胸径差异较大,18年后差异逐年减小(表1)。
2.3 造林密度对单株材积的影响
林分单株材积及单株材积年均生长量随林龄增加而增大,随造林密度增大而减小,10年生时,密度A、B 和C 的单株材积分别为密度D 的162.7%、137.0% 和123.8%;22年 生 时,分 别 为 密 度D 的131.9%、120.2%和118.5%(图3)。方差分析表明,12、14~18 及20年生时,各造林密度间单株材积差异极显著(P<0.01),其余林龄差异显著(P<0.05)(表1)。这是由于单株材积的差异主要由胸径差异决定,高密度林分对被压木较大的自疏强度使得各林分的密度差异逐年减小,胸径差异降低,单株材积差异随之减小。
2.4 造林密度对林分蓄积量的影响
林分蓄积量随林龄增加逐渐增大,22年生时,密度C 的林分蓄积量为372.29 m3/hm2,比密度A、B和D 分别高22.9%、28.9%和13.3%,林分蓄积量不仅取决于林分单株材积,更受林分保存株数影响;密度A、B、C 和D 林分蓄积年均生长量分别在13、12、12 和11年生时达到峰值,而后降低(图4)。方差分析表明,各密度的林分蓄积量及蓄积年均生长量差异不显著(表1)。
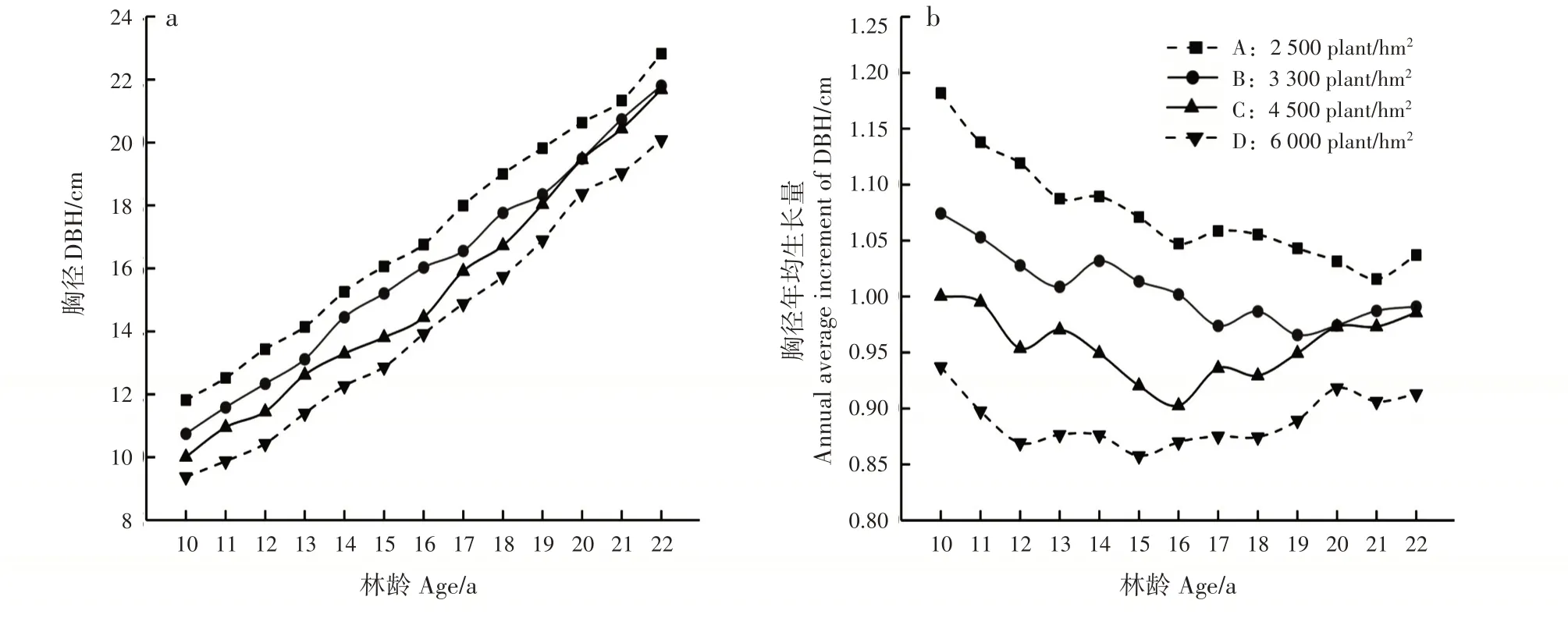
图2 不同造林密度马尾松林分不同林龄胸径生长变化Fig.2 Variation of DBH of P.massoniana stands with different ages in different planting densities
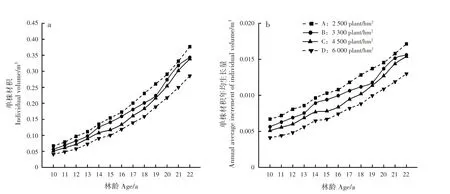
图3 不同造林密度马尾松林分不同林龄单株材积变化Fig.3 Variation of individual volume of P.massoniana stands with different ages in different planting densities
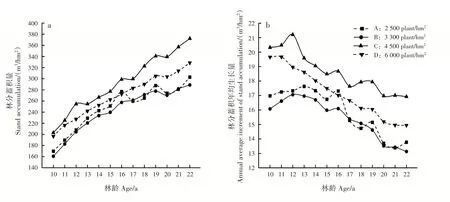
图4 不同造林密度马尾松林分不同林龄蓄积量变化Fig.4 Variation of P.massoniana stand accumulations with different ages in different planting densities
2.5 造林密度对林分直径结构的影响
偏度是描述数据总体取值分布对称性的统计量,SK>0表示数据分布右偏,此时数据的平均数>中位数>众数;SK<0则左偏,此时数据的众数>中位数>平均数。17年生前,造林密度越大的林分直径分布右偏越大,是由于高密度林分直径分化严重,极大径级林木拉高了林分平均胸径。密度A 和B 林分直径分布的非对称性随林龄增大而增大,密度C 和D 则随林龄增大而减小,是由于低密度林分可分配林地资源较多,林木个体竞争较小,多数小径级林木快速生长为中等径级林木,部分占据较优地位的中等径级林木生长至较大径级林木,导致直径分布右偏加重,而高密度林分自疏强度较大,被压木淘汰率高,使林分内趋于平均胸径的林木株数增多,直径分布右偏减小(图5a)。
峰度是描述数据分布形态陡缓程度的统计量。ST>0 表示数据分布为尖顶峰;ST<0 则为平顶峰。高密度的林分直径结构峰度随林龄增长而降低,低密度则升高,其中密度B 林分17年生后变为高狭峰(图5b)。直径变异系数反映林分直径分布的离散程度。各密度林分直径变异系数逐年增大,直径分化程度逐渐加重,分布范围逐渐离散(图5c)。低密度林分直径变异较小,利于特定材的定向培育,较高密度林分更适合多规格材种的培育需求。
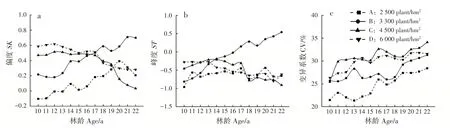
图5 不同造林密度马尾松林分不同林龄直径结构变化Fig.5 Variation of diameter structures of P.massoniana stands with different ages in different planting densities
各密度林分直径结构近似服从正态分布(图6a)。密度A、B、C 和D 直径最大分布频率对应的径阶分别为24、22、20 和18 cm,曲线峰值随造林密度增大而左移至较小径阶。径阶累积百分比相同时,对应的径阶随造林密度增大而降低,密度A、B、C 和D 在16 cm 径阶的累积百分比分别为12.7%、21.1%、21.9%和32.0%,表明密度越大的林分小径阶林木株数越多(图6b)。
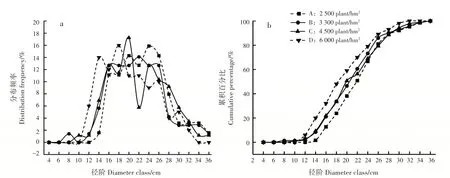
图6 22年生马尾松林分直径分布频率及径阶累积百分比变化Fig.6 Variation of diameter distribution frequency and cumulative percentage of diameter class of 22-year-old P.massoniana
株数最大分布率对应的径阶随林龄增加向高径阶方向进级,10~22年生之间,密度A、B、C和D林分直径的最大分布率所对应的径阶分别前进了5、7、6和5个径级。22年生时各密度林分大于20 cm径阶的株数占比分别为76.3%、66.2%、66.3%和52.0%,低密度林分直径生长更快,且大径级林木株数较多(表2)。
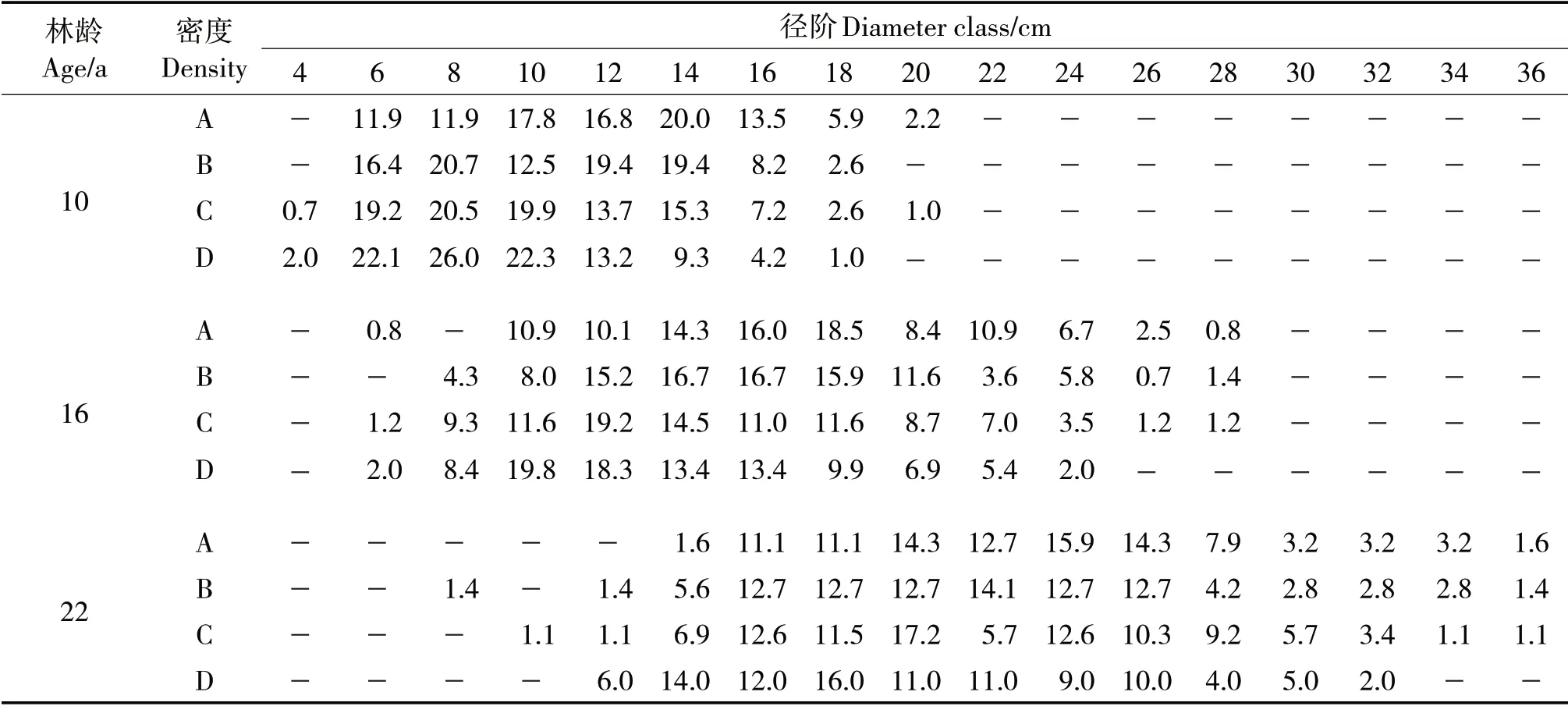
表2 不同林龄和造林密度下不同径阶的株数分布频率Tab.2 Distribution frequency of plants with different diameter classes at different ages and planting densities (%)
3 结论与讨论
本研究中,造林密度对树高和林分蓄积量的影响不显著,林分胸径及单株材积随造林密度增大而明显减小,这与温佐吾等[14]对马尾松和张阳峰等[15]对米老排(Mytilaria laosensis)的研究结果相同。这种规律在18年生后逐渐缩小,比谌红辉等[16]的研究结果延迟两年,原因是马尾松优良家系具有更长效的直径生长能力。密度C 的林分蓄积量最大,各密度林分蓄积量随林龄增加未表现逐渐趋近的态势,这与Zhang 等[17]的研究结果有差别,原因是试验林分在17年生时遭受台风侵害,低密度林分由于保存密度较小,林分稳定性和抵抗性稍差,林木枯损较大。密度A、B、C 和D 的林分蓄积年均生长量在13、12、12 和11年生时达到峰值,较当前广西工业原料林主伐林龄(16年生)早,也比同等立地指数及造林密度下的马尾松人工林数量成熟林龄(20、20、19 和18年生)提前[18]。林分密度越大,数量成熟林龄越早,与桉树[19]和油松(Pinus tabuliformis)[20]的研究结论相同。马尾松优良家系林分的树高、胸径、单株材积及蓄积量较普通马尾松林分均有提高[16,21],说明马尾松优良家系具有较快的生长速度,能在一定程度上缩短经营周期。
林分直径结构是最重要和最基本的林分结构,林木直径的分配状态直接影响林木的树高、干形、材积、材种及树冠等因子。本研究中,低密度林分直径分布的偏度逐年增大,峰度随林龄增加也有增大的趋势,变异系数相对较低,有利于单一材种培育目标的实现;高密度林分直径分布偏度和峰度逐年减小,变异系数相对较高,适合多规格材种的培育。这在Boyden 等[22]和段爱国等[7]的研究中得到佐证。株数的径阶分布频率及累积百分比表明低密度林分直径生长更快,且大径级林木株数更多,与谌红辉等[16,23]的研究结果一致。
综上所述,马尾松优良家系人工林的数量成熟林龄提前,但根据林分生长和直径结构判断,数量成熟时各林分小径材居多,工艺性能较差,尚不能达到工业用材的需求,在此时应适当间伐,促进林木持续高速生长。密度C(4 500株/hm2)的林分蓄积量最大,适宜培育短周期工业用材林;密度A(2 500株/hm2)林分内,大径材数量多、直径分布相对集中且右偏大,更适宜定向培育大径材林。
本研究从林分生长和直径结构两方面探讨不同造林密度对马尾松优良家系林分的影响,后续研究应结合林分自然稀疏规律、木材工艺特性、经济收益和林地生产力维续状况等进行深入研究和分析,综合探讨马尾松优良家系人工林的定向培育理论技术和可持续经营水平。
