苜蓿链霉菌内切β-N-乙酰氨基葡萄糖苷酶的克隆、表达及酶学性质
2020-08-05李玲聪胡少锋谷天燕吕晨茵刘妍池刘华顾金刚赵国刚
李玲聪,胡少锋,谷天燕,吕晨茵,刘妍池,刘华,顾金刚,赵国刚
1 河北农业大学 生命科学学院,河北 保定 071000
2 中国农业科学院农业资源与农业区划研究所,北京 100081
糖基化修饰是蛋白质翻译后的主要修饰方式之一,在蛋白质折叠形成正确的空间构象和发挥正常的生物功能中有着十分重要的作用[1]。根据糖基化位点的不同,糖基化修饰方式主要可以分为N-糖基化、O-糖基化和C-糖基化。N-糖基化是糖链与多肽链的天冬酰胺的自由氨基连接。O-连接的糖基化是糖链与多肽链上丝氨酸、苏氨酸等氨基酸的自由羟基共价连接。C-糖基化主要是糖链通过碳碳双键与多肽链上的色氨酸连接。在血浆等体液中的蛋白质大多发生N-糖基化,因此在以上几种糖基化修饰方式中,以N-糖基化的修饰方式最为普遍[2]。
在糖基化位点研究中常用的工具酶主要分为外切糖苷酶和内切糖苷酶两大类,外切糖苷酶沿着糖链末端进行切割,这种切割方式有助于分析寡糖链的连接顺序和糖基组成[3]。内切糖苷酶主要是水解糖链内部的寡糖链,从而释放完整的糖链[4]。内切糖苷酶提供了从糖肽去除完整的寡糖链的方法,长的寡糖链断裂成短的糖链,有利于对糖链的结构分析[5],因此内切糖苷酶在糖生物学中有着更为广泛的应用。而在不同种类的内切糖苷酶中,内切β-N-乙酰氨基葡萄糖苷酶(Endo-β-N- acetylglucosaminidase,ENGase) 在研究中的应用更加普遍,其主要作用于糖链上两个相邻的N-乙酰葡糖胺之间的β-1,4糖苷键[6]。ENGase酶处理糖蛋白后,在蛋白的糖基化位点上留下一个N-乙酰氨基葡萄糖胺,可以作为糖基化位点鉴定的一个标签。通过这个标签可以提高糖基化位点鉴定的正确性[7],而且经过ENGase酶处理后的糖蛋白仍具有生物学活性,可用于后续功能的研究[8]。同时,部分ENGase酶还具有转糖基作用。
ENGase在糖生物学研究和工业生产中具有重要应用价值。例如1998年,Yamamoto等在转糖活力的研究中运用Endo M酶在多肽T上连接上了唾液酸化的复合型糖链,提高了多肽T的稳定性[9]。2001年Haneda等利用Endo M酶以神经肽为底物,在其特定位置转移了唾液酸糖链,改善了神经肽的稳定性[10]。利妥昔单抗常用于白血病和淋巴瘤的治疗过程,Huang等首先利用Endo S的去糖基化作用将单抗上含有岩藻糖的非均一糖链切除,然后应用Endo D的突变体N322Q在糖基化位点连接上三甘露糖结构赋予了利妥昔单抗新的功能[11]。而最近几年ENGase更由于其去糖基化作用被用于食品工业中,人乳中含有大量的寡糖,目前已被证明能够有效地促进肠道中有益菌落的生长[12]。Garrido等利用从婴幼儿肠道中的双岐杆菌获取的内切糖苷酶Endo BI从人体中的乳铁蛋白和免疫球蛋白中获得寡糖链[13],该发现可用于处理奶制品加工过程中的副产物乳清,对于提高乳制品行业的经济效益有重要的研究意义[14]。
目前已从多种微生物中分离到内切β-N-乙酰氨基葡萄糖苷酶。1974年首次在褶皱链霉菌中分离纯化出Endo H[15],1985年Robert等从褶皱链霉菌中克隆并在大肠杆菌中表达了Endo H酶[16],以及脓链球菌来源的Endo S[17]、里氏木酶菌来源的Endo T[18]、金针菇在生长过程中分泌的Endo FV[19]、肺炎链球菌来源的Endo D[20]和冻土毛霉菌来源的Endo M[21]等越来越多的内切β-N-乙酰氨基葡萄糖苷酶家族新成员被发现,但是商品化的内切β-N-乙酰氨基葡萄糖苷酶种类非常有限,只由NEB等几家大型生物公司提供,但价格昂贵,并不利于大规模的应用。本文主要从苜蓿链霉菌中获取了一种新的内切β-N-乙酰氨基葡萄糖苷酶的基因,成功在大肠杆菌中实现高效表达,并进行了酶学性质的研究。本研究结果显示该酶有应用于生产的潜力。
1 材料与方法
1.1 材料
1.1.1 菌种和质粒
中国农业微生物菌种保藏中心提供苜蓿链霉菌Streptomyces alfalfaeACCC 40021。大肠杆菌Escherichia coliBL21 (DE3) 和Trans1-T1菌株购自北京全式金生物技术有限公司。
1.1.2 主要试剂
PfuDNA聚合酶、限制性内切酶和T4 DNA连接酶购自北京全式金生物技术有限公司。RNase B、Endo H均购自NEB (英国)。异丙基β-D-硫代半乳糖苷 (IPTG)、硫酸卡那霉素、其他分析纯化学试剂购自生工生物工程 (上海) 股份有限公司。
1.2 序列分析
利用NCBI CD-search (https://www.ncbi.nlm.nih.gov/Structure/cdd/wrpsb.cgi) 分析蛋白结构域,SignalP 5.0 (http://www.cbs.dtu.dk/services/SignalP/)进行信号肽预测并利用在线软件ClustalW(https://clustal.org) 进行多重序列信息比对。
1.3 重组质粒pET-30a(+)Endo SA的构建
从NCBI中获取目的蛋白Endo SA的核苷酸序列 (Locus_tag: A7J05_03925) 和氨基酸序列信息 (RfSeq: WP_019545283.1),用SignalP 5.0 对其信号肽进行分析,得到成熟蛋白的序列信息,并设计引物:Endo SA-F (BamHⅠ) 5′-CCGGATCC A TGAGCATGAAGAGGCGCA-3′和Endo SA-R(Hind Ⅲ) 5′-TTAGAAGCTT TCAGCGGCGTACC GC-3′,引物中下划线为酶切位点。
以苜蓿链霉菌ACCC 40021基因组(GenBank 登录号:NZ_CP015588.1) 为模板,分别以Endo SA-F和Endo SA-R为上下游引物扩增目的基因片段。PCR反应体系为:0.5 μL链霉菌ACCC 40021基因组DNA,0.5 μL TransStart FastPfuFly DNA Polymerase,2 μL 2.5 mmol/L dNTPs,0.5 μL 10 μmol上游引物F和下游引物R,5 μL 5×TransStart FastPfuFly Buffer,加ddH2O补足至总体积25 μL。PCR扩增的程序为95 ℃3 min;95 ℃ 30 s,58 ℃ 30 s,72 ℃ 30s,共35个循环;72 ℃ 10 min。核酸琼脂糖电泳检测并胶内回收目的片段,用限制性内切酶BamHⅠ和HindⅢ分别处理目的片段和载体pET-30a(+),T4 DNA连接酶16 ℃连接过夜,并热击转化到大肠杆菌Trans1-T1感受态细胞,酶切筛选阳性克隆,并送华大基因测序验证,得到重组质粒pET-30a(+)-Endo SA。
1.4 目的蛋白诱导表达与纯化
将pET-30a(+)-Endo SA热击转化到大肠杆菌BL21(DE3) 中,并涂布于含有50 µg/mL卡那霉素的LB平板上,37 ℃培养过夜。挑取单克隆,接种在2 mL (含50 µg/mL硫酸卡那霉素) LB液体培养基的EP管中,在37 ℃、转速为220 r/min的条件下培养过夜。以1% (V/V) 的过夜培养物接种于200 mL LB液体培养基(含50 µg/mL硫酸卡那霉素)的摇瓶中,37 ℃培养3 h,当OD600=0.6时,加入终浓度为0.1 mmol/L的诱导剂IPTG,在20 ℃、转速为220 r/min的条件下诱导培养20 h。4 ℃、12 000 r/min离心10 min,离心收集菌体,加入10 mL裂解缓冲液 (50 mmol/L NaH2PO4,300 mmol/L NaCl和10 mmol/L 咪唑,pH 8.0) 重悬,超声破碎。4 ℃、12 000 r/min离心10 min收集上清粗酶液,粗酶液用Ni-NTA纯化,其纯化步骤如下所示:将粗酶液加到含有Ni柱填料的纯化管中,用洗涤缓冲液 (50 mmol/L NaH2PO4,300 mmol/L NaCl和50 mmol/L咪唑,pH 8.0) 去除杂蛋白,再用250 mmol/L洗脱缓冲液(50 mmol/L NaH2PO4,300 mmoL/L NaCl和250 mmol/L 咪唑,pH 8.0) 洗脱目的蛋白。利用聚丙烯酰胺凝胶电泳 (Sodium dodecyl sulfate polyacrylamidegel electrophoresis,SDS-PAGE) 检测目的蛋白的纯度。
1.5 Endo SA的活力测定
在EP管中加入2 µL的RNase B、1 μL的糖蛋白变性缓冲液 (内含0.5% SDS,40 mmol/L DTT) 和4 µL的去离子水。将离心管置于金属浴中,100 ℃煮沸10 min,然后加入1 µL磷酸盐反应缓冲液 (pH 6.0) 和1 µL 10%的NP-40。最后加入1 µL的内切β-N-乙酰氨基葡萄糖苷酶,总反应体系共10 µL,37 ℃反应1 h。取3 μL的反应产物制样,进行SDS-PAGE (15%) 分析。酶活定义:1单位指10 μL的反应体系中,37 ℃条件下,在pH为6.0的磷酸盐缓冲液中,每1 h从10 μg变性RNase B中除去超过95%的碳水化合物所需要的酶量。
1.6 LC-MS/MS质谱分析
将Endo SA处理2 h的RNase B用15%的分离胶进行SDS-PAGE,切取去糖基化的RNase B的凝胶条带,送至中国科学院微生物研究所质谱平台鉴定。样品经胶内酶解处理,冷冻干燥后酶解肽段复溶于0.1% (V/V) 甲酸进行LC-MS/MS质谱分析(Orbitrap Fusion Tribrid mass spectrometer,Thermo Fisher Scientific)。肽段经预柱 (C18,粒径5 μm, 柱直径100 μm,柱长3 cm,Dr.Maisch GmbH) 除盐后经分析柱分离 (分析柱C18,粒径3 μm,柱直径75 μm,柱长15 cm,Dr.Maisch GmbH),流速400 nL/min。LC梯度分析条件:A相:0.1% (V/V) 甲酸,B相 (乙腈,0.1% (V/V) 甲酸)。3%–8% B相5 min,8%–20% B相40 min,20%–35% B相10 min,35%–80% B相3 min,80% B相2 min。所用质量分析器为Orbitrap和离子阱 (IT),碎裂方式HCD,碰撞能33。MS1扫描范围 (m/z350–1 550) 分辨率120 000;MS2分辨率30 000。
1.7 内切β-N-乙酰氨基葡萄糖苷酶的酶学性质测定
测定不同温度 (25 ℃、30 ℃、35 ℃、40 ℃、45 ℃、50 ℃、55 ℃、60 ℃、65 ℃) 和不同pH(50 mmol/L柠檬酸-柠檬酸钠缓冲液,pH 4–5;50 mmol/L磷酸盐缓冲液,pH 6–8;50 mmol/L甘氨酸-氢氧化钠溶液,pH 9)对Endo SA酶学活性的影响,以确定其最适温度和最适pH。
预先在不同温度 (25 ℃、30 ℃、35 ℃、40℃、45 ℃、50 ℃、55 ℃、60 ℃) 和不同pH (4.0、5.0、6.0、7.0、8.0、9.0) 的缓冲液预保温1 h,然后测定酶活,以未处理的重组酶在最适条件下测定酶活为100%,确定Endo SA的热稳定性和pH稳定性。
1.8 内切β-N-乙酰氨基葡萄糖苷酶对不同蛋白底物去糖基化分析
分别取酵母中表达的糖化酶、果胶酶和酸性蛋白酶作为反应底物,在10 µL的反应体系中,加入2 µL的底物、1 µL的糖蛋白变性缓冲液 (内含0.5% SDS,40 mmol/L DTT) 和5 µL的去离子水于1.5 mL的EP管中。将离心管置于金属浴中,100 ℃煮沸10 min。再加入1 µL的pH为6.0的磷酸盐缓冲液和1 µL 10%的NP-40。在管中加入1 μL的内切β-N-乙酰氨基葡萄糖苷酶,并加入等量缓冲液作为空白对照。在37 ℃的水浴锅中,水浴加热30 min。将反应产物进行SDS-PAGE (15%)分析。
2 结果与分析
2.1 Endo SA序列分析和氨基酸序列同源性比对
来源于Streptomyces alfalfaeACCC 40021的Endo SA基因全长942个碱基,共编码313个氨基酸。NCBI CD-search分析表明Endo SA属于糖苷水解酶第18家族 (GH18),只存在一个GH18催化结构域。SignalP 5.0显示其有长为32个氨基酸的信号肽。成熟蛋白质的等电点为5.74,理论分子量约为30.20 kDa。序列比对分析如图1所示,Endo SA与里氏木霉来源的Endo T (GenBank登录号:CAZ16624.1)、金针菇来源的Endo FV(GenBank登录号:ACY01407.1)、褶皱链霉菌来源的Endo H (GenBank登录号:AAA26738.1)、黄色脑膜炎黄杆菌来源的Endo F1 (GenBank登录号:KWR53347.1) 和化脓性链球菌来源的Endo S(GenBank ANC26024.1) 同源性分别为38.21%、38.19%、20.83%、9.88%和6.26%。以上结果表明Endo SA为一种新的内切β-N-乙酰氨基葡萄糖苷酶。

图1 GH18家族的ENGase酶的氨基酸序列比对Fig.1 Amino acid sequence alignment of GH18 ENGases.
2.2 苜蓿链霉菌Endo SA基因的克隆和表达
以苜蓿链霉菌基因组为模板,PCR扩增出目的基因,结果如图2所示,目的基因大小为863 bp,与预期大小一致。并成功构建了pET30a(+)-Endo SA表达载体。
2.3 苜蓿链霉菌EndoSA的表达与纯化
将构建重组质粒转化至大肠杆菌感受态细胞中,20 ℃诱导表达,Ni2+柱纯化,结果如图3所示,诱导菌株产生分子量为34.24 kDa的蛋白条带,与带N端His标签的Endo SA分子量一致,表明Endo SA成功可溶性表达。
2.4 质谱分析
对通过Orbitrap Fusion Tribrid检测得到的响应较高质谱峰进行Mascot谱库检索分析,确认34N*LTKDR39肽段的m/z值为948.487 6 (图4)。此肽段理论相对分子质量为745.82。该结果表明:经Endo SA处理后,RNase B唯一的糖基化位点第34位氨基酸天门冬酰胺上存在1个N-乙酰葡萄糖胺。Endo SA是内切β-N-乙酰氨基葡萄糖苷酶。
图2 Endo SA基因的PCR扩增结果Fig.2 PCR amplified product of Endo SA gene.Lane M: 1 kb plus DNA ladder; lane 1: the target gene fragment of Endo SA.

图3 目的蛋白SDS-PAGE分析Fig.3 SDS-PAGE analysis of target protein. Lane M:protein marker; lane 1: the crude supernatant; lane 2: the purified enzyme.
2.5 Endo SA的去糖基化活力分析
Endo SA对RNase B 的去糖基化活力经SDS-PAGE分析,如图5所示,在1 h内0.01 µg Endo SA可以将10 µg变性的RNase B(17 kDa)完全去糖基化。以RNase B为底物,Endo SA的比活力为1.0×106U/mg,显著高于商品化的Endo H(1.0×105U/mg)。
2.6 不同温度和pH对Endo SA活性的影响
在不同温度下,以RNase B为底物,测定Endo SA活性,如图6所示,该酶在25–45 ℃的范围内活力较高,当温度为35 ℃时,酶活力达到最高。酶活力随着温度上升而逐步下降。温度高于45℃,酶活性迅速下降。该酶的最适反应温度为35℃。在不同pH条件下测定Endo SA活性,如图7所示,该酶在pH 4–9的范围内均保持较高的活力,在pH 6的磷酸盐缓冲液中,该酶的活力最高。据此,该酶的最适pH为6。
2.7 Endo SA稳定性分析
以RNase B为反应底物,Endo SA的热稳定性结果如图8所示,该酶在30–45 ℃的范围内保温1 h,保持100%活性;当温度逐渐上升到50 ℃以上,酶活快速丧失。结果表明该酶在25–45 ℃这个温度范围内有良好的热稳定性。Endo SA的pH稳定性结果如图9所示,该内切酶在pH为4.0–8.0的条件下保温1h,活力均大于90%,具有很好的pH稳定性。

图4 Endo SA处理的RNase B的质谱分析Fig.4 MS/MS spectra analysis of RNase B treated by Endo SA.

图5 Endo SA去糖基化结果分析Fig.5 Deglycosylation results of Endo SA.Lane M:protein marker; lane 1: add 0.001 μg enzyme; lane 2: add 0.005 μg enzyme; lane 3: add 0.01 μg enzyme; lane 4:add 0.015 μg enzyme; lane 5: add 0.02 μg enzyme; lane 6:negative control; lane 7: RNase B digested by 1.5 μg Endo H.
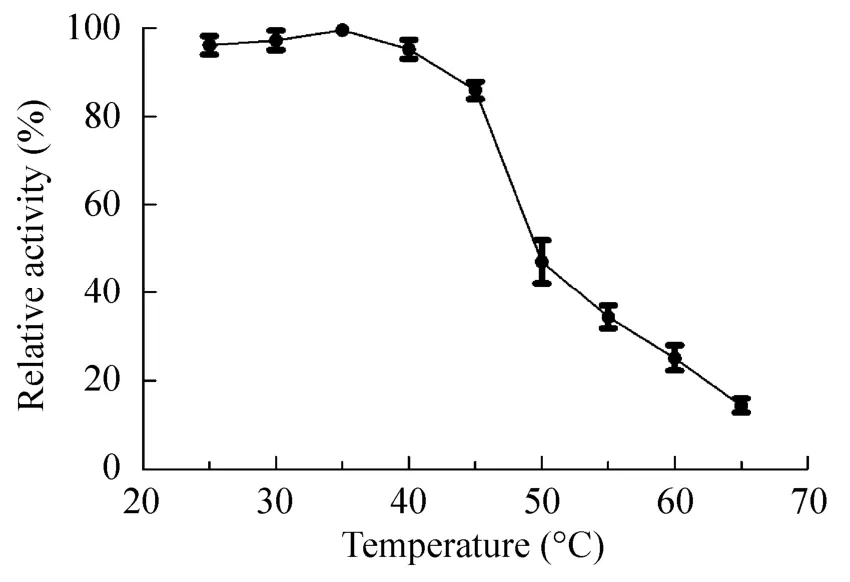
图6 温度对Endo SA酶活影响Fig.6 Effect of temperatures on the activity of Endo SA.
2.8 Endo SA对不同糖基化蛋白底物活力分析
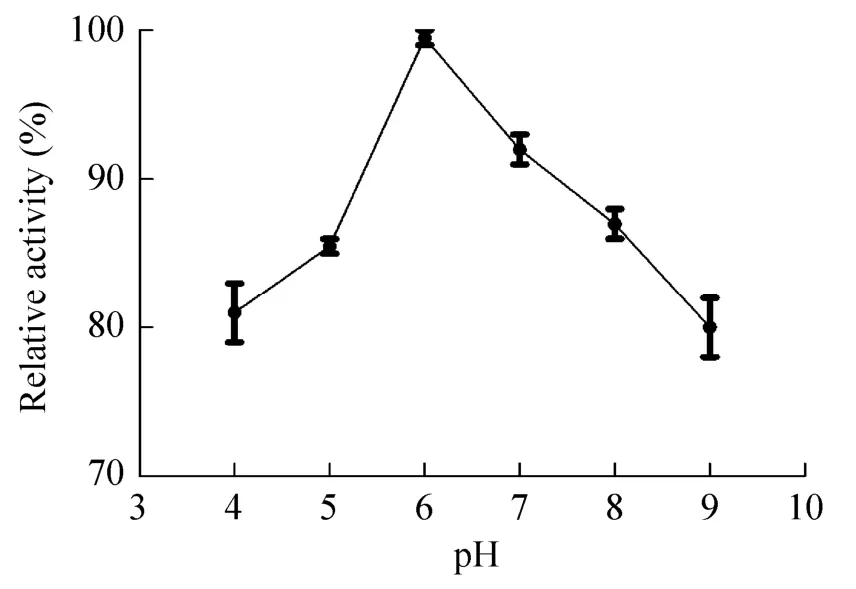
图7 pH对Endo SA酶活影响Fig.7 Effect of pH on the activity of Endo SA.

图8 Endo SA热稳定性分析Fig.8 Thermo-stability profile of Endo SA.

图9 Endo SA pH稳定性分析Fig.9 pH stability profile of Endo SA.
本实验选择酵母中表达的糖化酶(理论分子量65.8 kDa)、果胶酶(理论分子量35.4 kDa)和酸性蛋白酶(理论分子量40.3 kDa)为底物,根据反应前后蛋白分子质量的变化来验证该酶对不同蛋白底物的去糖基化活力,结果如图10所示,该内切酶对于不同的蛋白底物能够完全地去糖基化,可以作为糖生物学糖基化位点研究中的工具酶。

图10 Endo SA对不同蛋白底物的去糖基化分析Fig.10 The deglycosylation of Endo SA towards different protein substrates.Lane M: protein marker; lane 1: saccharification enzyme; lane 2: glucoamylase treated by Endo SA; lane 3: pectinase; lane 4: pectinase treated by Endo SA; lane 5: acid protease; lane 6: acid protease treated by Endo SA.
3 讨论
随着糖生物学和蛋白组学的发展,对于多肽链上连接的糖链的研究越来越受到科学家的关注,而糖苷酶在糖链的组成分析中发挥着重要作用[4]。
本研究从苜蓿链霉菌ACCC 40021中发现了一个新的GH18家族基因,通过对其进行克隆表达和活性测定,发现该酶不具有几丁质酶活性,以RNase B为底物对该酶进行去糖基化功能分析及质谱鉴定结果表明,Endo SA是一种内切β-N-乙酰氨基葡萄糖苷酶。通过序列比对发现,该酶氨基酸序列与GH18家族其他内切糖苷酶同源性低,由此可以确定本文发现了一种新的内切-β-N-乙酰氨基葡萄糖苷酶。
内切-β-N-乙酰氨基葡萄糖苷酶均能水解糖蛋白N-糖链中几丁二糖核心结构的2个N-乙酰氨基葡萄糖之间的β-(1,4) 糖苷键。根据水解N-糖链的组分和类型不同,目前商品化的β-N-乙酰氨基葡萄糖苷内切酶分为:水解高甘露糖和一些杂糖组成的N-糖链的Endo H;专一性切除天然免疫球蛋白IgG重链的N-糖链的Endo S;切割paucimannose组成的N-糖链的Endo D。本研究中,Endo SA不能移除IgG重链的N-糖链(结果未显示),而是高效地去除糖蛋白 (RNase B) 的高甘露糖和一些杂糖组成的N-糖链,因此,Endo SA是一种新型的Endo H类内切-β-N-乙酰氨基葡萄糖苷酶。
NEB等公司提供商品化的Endo H,是目前蛋白糖基化研究的主要工具酶,其比活力为1.0×105U/mg;而阿维链霉菌来源的ENGase比活为90 000 U/mg[22],粪肠球菌来源的内切-β-N-乙酰氨基葡萄糖苷酶的比活力为1.0×104U/mg[23]。本研究中,Endo SA的比活力达到1.0×106U/mg,远高于已报道的和商品化的所有内切-β-N-乙酰氨基葡萄糖苷酶。Endo SA在35 ℃的酶活力最高,并且热稳定性高,在25–45 ℃的条件下酶活力保持不变,能够适用于蛋白糖基化研究和酶法工业生产。而阿维链霉菌来源Endo H在30 ℃条件下酶活力只能达到最适反应温度下的65%。该酶在pH范围为4–8保持稳定性,最适pH为6,与商品化Endo H、已报道的阿维链霉菌和脑膜脓毒性黄杆菌来源的ENGase的最适pH相近[22];但该酶在pH 4.0–8.0保持95%以上的活力,在pH 9的条件下保持1 h后,仍保持有近80%的活性。表明该酶具有很好的pH适应性和稳定性。
随着对糖苷酶研究的深入,越来越多的应用不断地被挖掘,例如在食品工业利用奶制品加工的副产物生产寡糖链。在医药工业中,通过去除糖链,更好地发挥药效以及合成新的药物。而本研究新发现的糖苷酶以其本身所具有活力高、稳定性强的特性,可作为糖生物学研究中的工具酶和工业化生产的生物催化剂。
