基于16S rRNA基因分析双斑东方鲀肠道微生物多样性
2020-07-31王松刚
雷 阳,王松刚,陈 钰,姜 尧,郑 毅,2
( 1.福建师范大学 生命科学学院,福建 福州 350117; 2.福建师范大学 南方海洋研究院,福建 福州 350117 )
河鲀鱼肉味鲜美,广受人们的喜爱,但因其体内含有河鲀毒素,因此河鲀的食用受到限制。早在1990年,我国在《水产品卫生管理办法》中写到:河鲀鱼有剧毒,不得流入市场。河鲀毒素是一种剧毒,其毒性是氰化物的1000余倍,不慎食入会口舌、肌肉麻痹,四肢无力,恶心呕吐,瘫痪,严重者会呼吸困难,窒息而亡,并且迄今为止尚无可以解除此毒的特效药物[1]。关于河鲀毒素的起源一直存在争议,主要有内因说和外因说,内因说主张河鲀毒素是由其自身产生,河鲀鱼体内存在某种机制可以将摄入的食物转化为毒素,1998年kendo[2]证实,胚胎可以产生河鲀毒素,但目前该假说由于证据不足,尚未得到研究者的普遍认可。外因说主要指食物链假说和微生物起源假说。王少蓉等[3]在黄鳍东方鲀(Takifuguxanthopterus)的卵巢中筛选出产毒能力较强的EL10菌株。邢磊[4]自横纹东方鲀(T.oblongus)肠道分离到菌株HDC13、HDC35、HDC37,将3株菌混合培养后其产物对老鼠有较强的致死效果,这为外因说提供有力的证据。人工养殖的河鲀通常低毒或无毒,这一差异与野生和养殖河鲀肠道微生物的菌群结构有很大联系。笔者拟通过16S rRNA基因V3~V4区的Miseq PE300测序,对野生和养殖双斑东方鲀(T.bimaculatus)肠道微生物进行多样性分析,以期揭示野生和养殖双斑东方鲀肠道微生物中优势菌群的组成,为探明菌群多样性的进化规律奠定基础。
1 材料与方法
1.1 样品采集
2018年11月进行样品采集,3尾野生双斑东方鲀采于厦门同安海域,3尾养殖双斑东方鲀采于漳州养殖场,6个样品规格相似。根据国家标准GB 5009.206—2016检测,野生双斑东方鲀肠道内河鲀毒素的含量小于1.5 MU/g,养殖双斑东方鲀肠道内河鲀毒素含量小于0.04 MU/g。
1.2 样品处理
用75%酒精擦拭河鲀的表面3次,然后在无菌的条件下,用灭过菌的解剖器取其肠道内容物于灭菌的PE管中,于-80 ℃保存备用。
1.3 DNA的提取和扩增
用E.Z.N.A. Stool DNA Kit试剂盒提取DNA。将提取的DNA样品用1%琼脂糖凝胶电泳检测,利用分光光度法(260 nm/280 nm光密度比)检测含量和纯度。检测后将样品保存于-20 ℃备后续试验使用。
以提取DNA为模板,细菌16S rDNA V3~V4扩增引物为338 F(5′-ACTCCTACGGGAGGCAG CAG-3′)和806 R(5′-GGACTACNNGGGTATCT AAT-3′)进行PCR。PCR反应体系:12.5 μL 2×Taq PCR MasterMix,3 μL BSA(2 ng/μL),2 μL引物(5 μmol/L),2 μL模板DNA和5.5 μL ddH2O。反应参数:95 ℃预变性5 min, 95 ℃变性45 s,55 ℃退火50 s,72 ℃延伸45 s,32个循环,72 ℃延伸10 min。PCR产物经检测后按等量混合,进行Illumina测序。
1.4 数据分析处理
PCR扩增产物采用Illumina MiSeq平台进行双端测序,测序原始序列上传至美国国立生物技术信息中心的SRA数据库,下机数据去除接头序列和引物并拼接,进一步去除嵌合体、短序列后得到优质序列。根据接头序列对各个样本进行相似度为97%的运算分类单元(OTU)聚类,将得到的运算分类单元与Silva数据库(128)进行对比,得到每个运算分类单元对应的物种分类信息。
样本的Alpha多样性通过Chao-1、observed-species、PD_whole_ tree、香农指数表示,其中observed-species、Chao-1、PD_whole_ tree用来评估样本物种丰度,而香农指数用来评估样本多样性。
Chao1:即菌种丰富度指数,用以估计群落中的运算分类单元数目。
Schao1=Sobs+n1(n1-1)/2(n2+1)
式中,Schao1为估计的运算分类单元数,Sobs为观测到的运算分类单元数,n1为只有一条序列的运算分类单元数目,n2为只有两条序列的运算分类单元数目。
observed_species:随测序深度的增加,实际观测到运算分类单元的个数。
goods_coverage:观测深度。
C=1-n1/N
式中,n1为只有含一条序列的运算分类单元数目,N为抽样中出现的总的序列数目。
香农指数:用来估算样品中微生物多样性指数之一。
H=-∑PilnPi
式中,H为香农指数,Pi为样品中属于第i种的个体的比例。
利用QIIME软件(版本号1.8)估计样本多样性并制作韦恩图。
使用R语言vegan包、vegdist和hclust进行距离计算和聚类分析,构建基于未加权距离矩阵,UPGMA方法聚类建树,并将聚类结果与各样品在门、纲、目、科、属水平上的物种相对丰度整合展示。
2 结果与分析
2.1 高通量测序结果和基本数据统计
通过Illumina Miseq PE300高通量测序,在去除接头序列和引物并拼接后,经进一步去除嵌合体、短序列后,6份样品共得到优质序列99 500条,平均序列条数为16 583条,其中优质序列最多为27 781条,优质序列最少为7182条,对优质序列长度统计发现优质序列多集中在400~420 bp和420~440 bp,然后将优质序列进行97%相似度的运算分类单元聚类分析。
由图1~2可见,当测序量在2000以下时,运算分类单元的数目在迅速增加,香农指数趋于平缓,当测序量在6000多时,所有样品的稀释曲线仍在上升而香农指数已经达到饱和,尽管增加更多的数据量会产生新的运算分类单元,但目前的测序量已经能够反映样品中细菌的多样性。
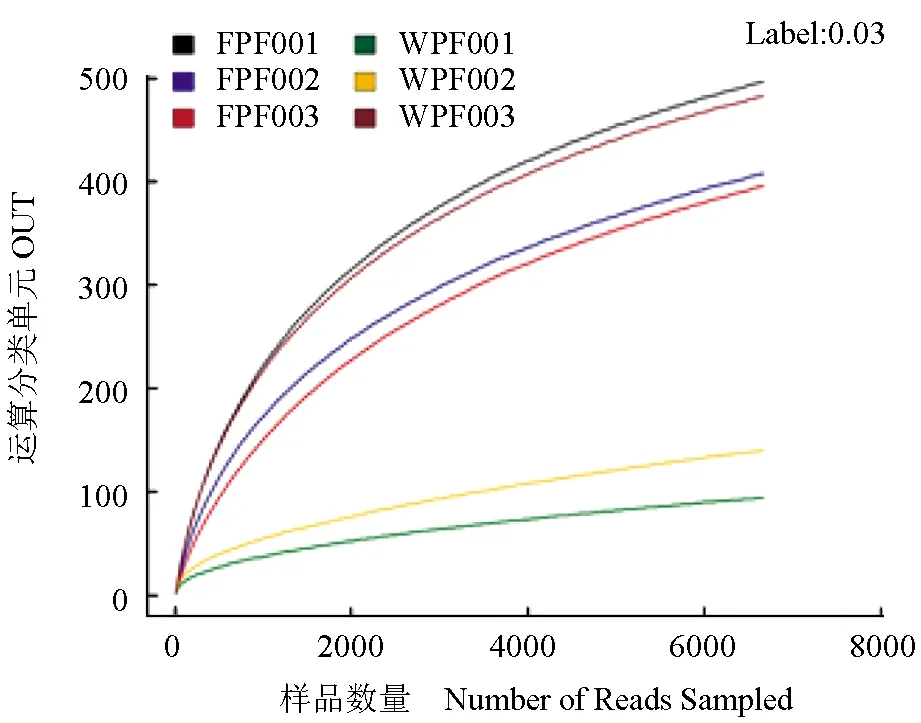
图1 样品稀释曲线
2.2 群落结构分析
对样品中的微生物群落进行分析,由于序列长度以及数据库的原因,运算分类单元注释只获得了门、纲、目、科、属几个水平,微生物种类和相对丰富度无法具体到种水平的分类。
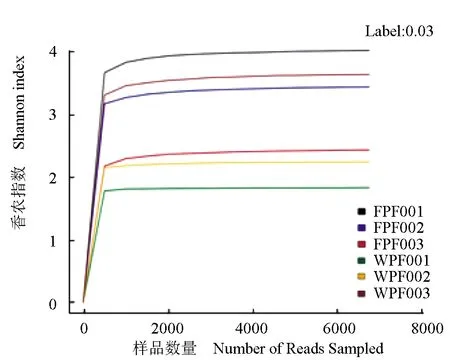
图2 细菌香农指数
2.2.1 细菌在门水平上进行分类及比较
测得序列经过鉴定,在门水平主要为变形菌门、拟杆菌门、厚壁菌门、螺旋菌门、硝化螺旋菌门、梭杆菌门、芽单胞菌门、放线菌门、绿弯菌门,从门水平的分析可以看出:变形菌门在野生双斑东方鲀和养殖双斑东方鲀中都占有很大的比例,为优势菌,梭杆菌门在野生双斑东方鲀中占的比例相对较大,分别为13.13%、7.65%和0.09%,而梭杆菌门在养殖双斑东方鲀中占的比例相对较少,分别为0.02%、0.02%和0.03%。螺旋菌门和拟杆菌门在养殖双斑东方鲀中明显比在野生双斑东方鲀中所占比例高(图3)。
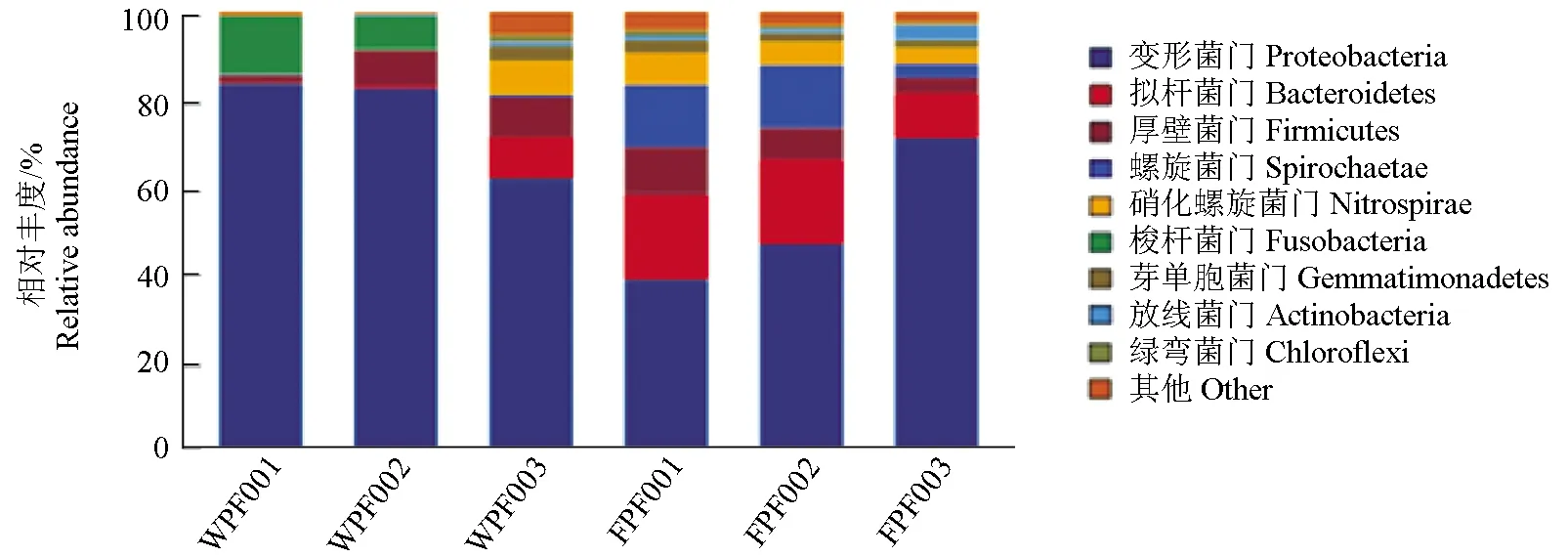
图3 细菌门水平的菌群分布
2.2.2 细菌在属水平上进行分类及比较
从属的水平进行分析得知,养殖双斑东方鲀中相对丰度大于1%的共有8个属,分别为弓形杆菌属(Arcobacter,39.63%)、短螺旋体属(Brevinema,10.44%)、理研菌属(Rikenella,8.25%)、弧菌属(Vibrio,1.14%)、硝化螺菌属(Nitrospira,5.26%)、乳杆菌属(Lactobacillus,1.31%)、反硝化细菌(Denitratisoma,1.1%)、拟杆菌属(Bacteroides,1.07%),未鉴定的细菌属占总细菌属的16.5%。野生双斑东方鲀中相对丰度超过1%的有弓形杆菌属(35.97%)、内皮单胞菌属(Endozoicomonas,17.56%)、弧菌属(7.21%)、硝化螺菌属(2.6%)、希瓦氏菌属(Shewanella,4.2%)、纤毛菌属(Propionigenium,3.52%)、鲸杆菌属(Cetobacterium,3.44%)、Romboutsia(1.01%),未鉴定的细菌属占总细菌属的12.31%。希瓦氏菌属、纤毛菌、发光杆菌属(Photobacterium)、假交替单胞菌属(Pseudoalteromonas)是野生双斑东方鲀特有的(图4)。

图4 细菌属水平的菌群分布
2.3 双斑东方鲀肠道细菌Alpha多样性分析
从Chao1、observed-species、PD_whole_ tree、香农指数4个方面使用Alpha多样性指数进行统计分析,由图5可见,Chao1、observed-species、PD_whole_ tree、香农指数均为养殖型双斑东方鲀大于野生型双斑东方鲀,表明养殖型双斑东方鲀肠道细菌多样性和丰富度高于野生型双斑东方鲀。
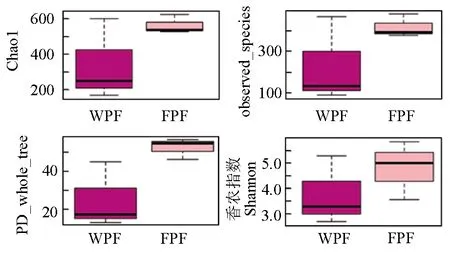
图5 Alpha多样性指数box图
韦恩图(图6)可以直观地反映样品共有和各自特有的运算分类单元数目,对样品进行相似度97%的聚类,野生双斑东方鲀运算分类单元数目为524,独有的运算分类单元数目为68, 其中OTU-18占3.415%,OTU-61占0.955%,OTU-33占0.945%,OTU-48占0.570%、OTU-81占0.37%,养殖双斑东方鲀运算分类单元数目为576,独有的运算分类单元数目为120,OTU-274占0.075%、OTU-565占0.040%。养殖双斑东方鲀的运算分类单元数目比野生双斑东方鲀的运算分类单元数目多,说明养殖双斑东方鲀肠道细菌的种类比野生双斑东方鲀肠道细菌的种类更丰富。
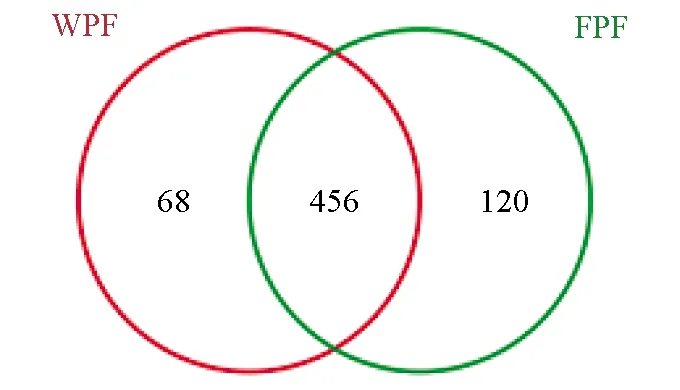
图6 样品中细菌群落共享和独有运算分类单元的韦恩图
2.4 Beta多样性分析
主成分分析,一种可以对测得的数据进行简单化分析的技术。通过分析不同样本运算分类单元(97%相似性)组成可以反映样本间的差异和距离,将数据进行R语言主成分作图和分析(图7)。基于UniFrac的非加权分析,第一主成分和第二主成分的贡献率分别为60.19%和20.80%。养殖双斑东方鲀生活范围相对有限,生活环境相似,因此3个样品比较集中,野生双斑东方鲀由于生活范围广泛,生活环境多变,3个样品相对比较分散。
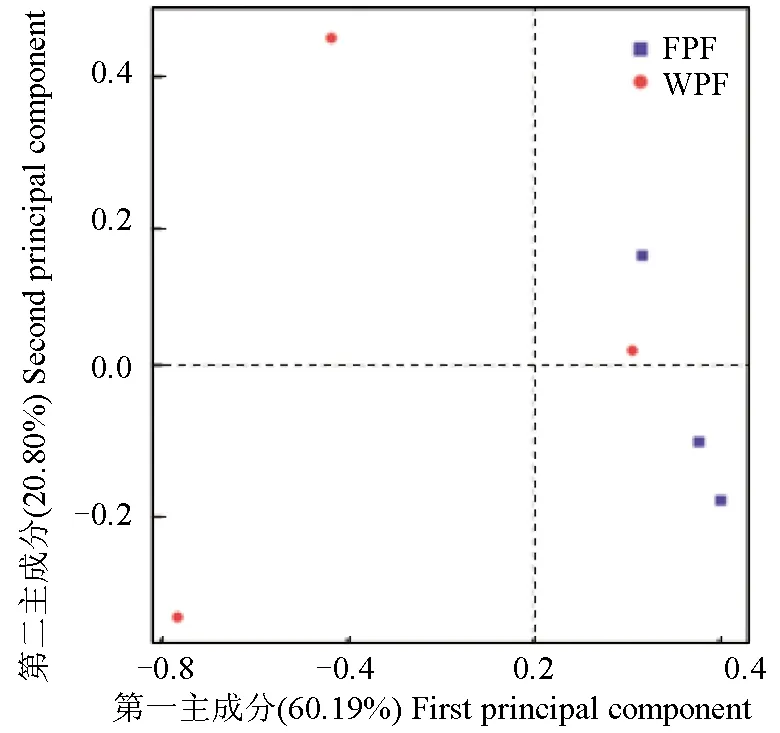
图7 基于运算分类单元水平的主成分分析
聚类分析的热图见图8。图中横坐标代表样本的名称,纵坐标代表聚类情况,图中不同颜色的方块代表每个样品属的丰度,颜色的深浅变化的差异性和相似性代表不同样品群落组成的相似程度。弓形杆菌属在两种双斑东方鲀中占的比值最大,在养殖双斑东方鲀占比39.6%,在野生双斑东方鲀占比35.97%,希瓦氏菌属、纤毛菌、发光杆菌属在野生双斑东方鲀肠道有发现,而在养殖双斑东方鲀中没有发现,Brevinema在养殖双斑东方鲀中占10.4%,在野生双斑东方鲀中只占0.26%,表明不同生活环境的鱼肠道微生物群落的组成不同且差异性较大。
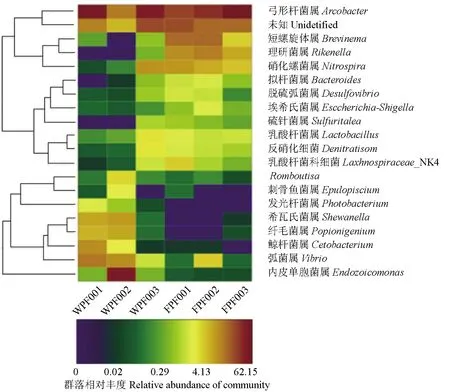
图8 聚类分析的热图(属水平)
3 讨 论
3.1 野生和养殖双斑东方鲀鱼肠道微生物多样性分析
肠道是动物体物质代谢与吸收的主要场所,而肠道的重要组成部分是肠道微生物,肠道微生物在动物的发育、进化、机体代谢方面有重要的作用[5-6]。对于在脊椎动物中占比最大的鱼类肠道中的微生物也备受人们的关注。本试验采集野生和养殖双斑东方鲀样品规格相似,由于生境不同,两者肠道细菌菌群的多样性有明显的差异,野生双斑东方鲀和养殖双斑东方鲀的总运算分类单元数目并无明显差异,但野生双斑东方鲀独有的运算分类单元数目为68,其中OTU-18占比最大,为3.415%,养殖双斑东方鲀独有的运算分类单元数目为120,占比最大的为OTU-274,仅占0.075%。Alpha多样性分析结果显示,养殖双斑东方鲀肠道细菌多样性和丰富度高于野生双斑东方鲀,这是因为宿主的饮食、生活环境、年龄等因素都会影响肠道微生物群落的结构与组成[7]。在门水平上的分析结果显示,变形菌门为野生双斑东方鲀和养殖双斑东方鲀肠道菌群的优势菌。梭杆菌门在野生双斑东方鲀肠道中占有很大比例,在养殖双斑东方鲀中所占比例相对较少。文献报道,变形菌门和梭杆菌门是大多数鱼类肠道微生物的主要菌群,苏月华等[8]对33只白甲鱼(Onychostomasima)肠道内容物进行高通量测序结果显示,白甲鱼肠道菌群主要是梭杆菌门、变形菌、螺旋菌门、厚壁菌门,其中梭杆菌门和变形菌门占比高达82%。Wu等[9]用高通量测序方法对草鱼肠道菌群进行研究,发现其主要为变形菌门、厚壁菌门、放线菌门。属水平上的分析结果显示,弓形杆菌属、弧菌属、硝化螺菌属在两者中均占有较大比例。弧菌属为养殖和野生双斑东方鲀肠道菌群的优势菌之一,这一结果符合弧菌为海水鱼肠道的主要菌群的说法[10]。据报道,红鳍东方鲀(T.rubripes)[11]肠道常见的细菌为弧菌属、希瓦氏菌属、假交替单胞菌属和芽孢杆菌属(Bacillus)。弧菌属、希瓦氏菌属、芽孢杆菌属在星点东方鲀(T.niphobles)[12]肠道内容物中也有发现。邢磊[4]自横纹东方鲀肠道内容物筛到49株菌,其中弧菌属有28株,发光杆菌属有1株,假交替单胞菌属3株,希瓦氏菌属有4株,这些细菌在双斑东方鲀肠道中均有发现。
3.2 野生和养殖双斑东方鲀鱼肠道微生物多样性与河鲀毒素的相关性
由于红鳍东方鲀得病率高,死亡率高,而水域对暗纹东方鲀具有限制性,福建地区近几年主要以养殖菊黄东方鲀(T.flavidus)和双斑东方鲀为主,双斑东方鲀的抗病能力较菊黄东方鲀强,因此双斑东方鲀投苗量越来越多[13]。野生双斑东方鲀和养殖双斑东方鲀最大区别在于河鲀毒素含量的差异性,前者比后者含量高37.5倍。关于河鲀毒性产生有微生物起源的学说,近几年,陆续发现了多种能产生河鲀毒素的细菌,主要有弧菌、假单胞菌、芽孢杆菌、交替单胞菌(Alteromonas)、发光杆菌、放线菌等。岳田芳[14]在海藻希瓦氏菌(S.algae)发酵产物中检测到了河鲀毒素。王少蓉等[3]在黄鳍东方鲀中筛选出产河鲀毒素能力较强的EL10菌株,经鉴定也属于希瓦氏菌属。王冠男等[15]指出,海胆中存在的假交替单胞菌可以产河鲀毒素,而从红鳍东方鲀体内分离出的达松维尔拟诺卡氏放线菌(Nocardiopsisdassonvillei)和梭形芽孢杆菌(B.fusiformis)可以产河鲀毒素。希瓦氏菌属、纤毛菌属、发光杆菌属、假交替单胞菌属存在于野生双斑东方鲀肠道中,而在养殖双斑东方鲀中未发现,因此肠道产毒菌群的差异性与野生和养殖双斑东方鲀毒性的差异可能具有某种联系,这为探究河鲀毒素微生物源提供一定的科学依据。
