文冠果雌蕊发育及其糖分积累的生理机制研究
2020-07-29景文昭高述民
景文昭,高述民
(北京林业大学 生物科学与技术学院,北京 海淀 100083)
文冠果(Xanthocerassorbifolium)是一种中国北方特有的落叶灌木或者小乔木树种,属无患子科文冠果属[1-2]。因其花期长,具有较高的观赏价值。其种子营养价值高,种仁含油量高,是重要的木本油料作物和生物能源植物[3-4]。文冠果花为杂性花,顶生花序为两性花,即可孕花;侧生花序为雄花,即不孕花。文冠果花多但座果率低,俗称“千花一果”,主要原因是雄花数量远远高于两性花,导致后期落花落果比较严重[5]。
前人研究发现,制约植物座果率与产量的因素比较多,如小麦(TriticumaestivumL.)不孕花中有较高的IAA及GA含量,苹果(Malusdomestica)不孕与其花发育过程中的高温具有较大关系。文冠果座果率低的主要原因是开花后至结果前阶段落花落果严重,与树体营养物质的匮乏有着直接关系[6-8]。植物的营养物质主要为糖类物质,可溶性糖包括蔗糖、果糖和葡萄糖。糖分积累在梨(Pyrus spp)[9]、枣(ZiziphusjujubaMill.)[10]、薹菜[BrassicacampestrisL. ssp.Chinensis(L.) Makino var.tai-tsaiHort][11]等作物中已进行了大量研究,涉及糖分积累与糖代谢酶相关性的研究较多。目前对文冠果的研究集中在雌雄蕊败育[12]、花性别分化[13]、雄蕊不育[14]以及自交不亲和[15]等方面,其中雄蕊不育等内部败育机理方面的研究较多,对于雌蕊败育也只停留在其发育过程中的结构研究[16]。因此在前期观察解剖的基础上,利用细胞组织学以及生理糖含量的测定,分析去顶处理后不同位置花序子房膨大率与其糖含量的相关性,从生理水平上分析了雌蕊败育的内部机制,对进一步研究文冠果生殖败育与产量制约因素具有重要意义。
1 材料与方法
1.1 材料
试验材料为北京市昌平区大东流苗圃(N39°54′20′′, E116°25′29′′)内生长健康的10年生文冠果树,其树高、胸径、冠幅、树势、树形等性状基本一致,并已经进入结果期,能正常开花结实的果树。
1.2 方法
1.2.1 采样时期确认
第一年分别在文冠果花序芽鳞伸出2 d、6 d、10 d、16 d取样,每次采集两性花(源自顶生花序)和同期雄花(源自侧生花序)花蕾,采集之后迅速放入指定的离心管中,用于测定花蕾和子房长度和直径。利用游标卡尺对顶花序、侧花序花蕾及子房长度和直径进行测量。测量之后取平均值列表统计,绘制不同位置花蕾和子房的长度生长曲线。
1.2.2 处理设计
根据第一年生长动态曲线分别在四个时期采集两性花(源自顶生花序)、同期雄花(源自侧生花序)和同期转化两性花(源自去顶后原第一位侧生花序)雌蕊组织。其中,去顶处理在芽鳞未伸出之前进行,采样从芽鳞伸出开始,间隔3~4 d取一次样,生物学重复3次。采集之后放入指定离心管用于后续试验。
1.2.3 文冠果雌蕊外部形态观察
体视显微镜观察法:将采集到的不同位置花蕾置于体视镜下,一部分剥离除雌蕊以外的花器官,另一部分只剥离苞片,留下花药和中心的雌蕊组织,拍照记录文冠果不同位置花序雌蕊组织的发育情况。
扫描电镜观察法:将采集到的不同位置花蕾迅速剥离除雌蕊以外的花器官,马上放入FAA固定液中固定12 h以上。固定完成后依次使用浓度为70%、90%、100%的乙醇脱水处理,一次20 min。然后用100% 醋酸异戊酯置换两次,每次用时20 min。置于二氧化碳临界点干燥仪中干燥,样品完全干燥后取出。将观察面朝上用双面导电胶粘到样品台上,离子溅射仪喷镀后,扫描电子显微镜(日本日立,su8010)观察、拍照。
1.2.4 文冠果雌蕊中不同种类糖含量的测定
将采集到的花蕾迅速剥离除雌蕊以外的花器官后放入指定离心管,在-80℃冰箱保存用于糖含量测定。称取0.05 g左右的样品放入研钵中,加入少量80%乙醇磨碎成浆,将磨碎的样品转移至5 mL离心管中,置于80℃水浴中浸取30 min,不时搅拌。取出后冷却,离心5 min,收集上清液,其残渣加2~3 mL 80%乙醇反复提取2次(各10 min),冷却离心,合并上清液,用蒸馏水定容至5 mL,为可溶性糖待测液,每个样品重复3次。不同种类的糖含量的测定采用蒽酮硫酸法[17-19]。
1.2.5 文冠果雌蕊蔗糖代谢相关酶活性的测定
酶的提取:在0~4℃条件下称取样品0.05 g左右,置于预冷的研钵中,液氮中研磨5~10 min,加入4倍体积的提取液(0.1 mol·L-1Tris,pH=7.0,10 mmol·L-1MgCl2,2%乙二醇,20 mmol·L-1巯基乙醇,2 mmol·L-1EDTA)。在水浴中提取,4500 r·min-1冷冻离心20 min,取上清液透析24 h后,离心定容至5 mL,酶提取液用于酶活性检测。合成酶(SPS、SS-SD)和分解酶(SS-DD)酶活性按照Hubbard等和Lowell等的方法,并改进后进行检测[20-21]。
1.2.6 文冠果不同位置花序雌花数量的统计
在盛花期统计顶花序、侧花序以及转化花序雌花数量,计算雌花比例,即子房膨大率。用SPSS 19.0对数据进行显著性分析,Excel进行图表绘制。
2 结果与分析
2.1 文冠果不同位置花序子房膨大率
如图1所示,不同位置文冠果花序的子房膨大率从大到小依次为:顶花序>转化花序>侧花序,顶花序的子房膨大率显著高于侧花序和转化花序,去顶处理后的转化花序子房膨大率显著高于侧花序,说明去顶处理显著提高了花序的子房膨大率。

图1 文冠果不同位置花序的子房膨大率
2.2 文冠果不同位置花蕾和子房的生长曲线
文冠果顶花序雌蕊花蕾和子房的长度和直径在整个发育阶段不断增加,侧花序雌蕊花蕾的长度和直径也在不断增长,但始终小于顶花序雌蕊花蕾的长度和直径,子房的长度在大孢子母细胞分裂Ⅰ期之后不再增长,直径开始减少(图2A;图2B),说明子房开始败育。可以看出,从花芽期到开花前期,顶花序子房的长度和直径均与花蕾的长度和直径呈正相关增长,相关系数分别为0.9763和0.9925。并且在侧花序雌蕊子房开始败育之前,其子房的长度和直径也均与花蕾的长度和直径呈正相关,相关系数分别为0.9790和0.9567。

图2 文冠果顶、侧花序子房长度、直径随花蕾长度(A)、直径(B)的变化趋势
根据文冠果不同位置雌蕊的生长曲线,并结合文冠果的外部形态表现,将取样时间分为四个时期:Ⅰ期-花芽期(芽鳞伸出0~2 d),Ⅱ期-大孢子母细胞期(芽鳞伸出3~8 d),Ⅲ期-大孢子母细胞减数分裂Ⅰ期(芽鳞伸出9~12 d),Ⅳ期-大孢子母细胞减数分裂Ⅱ期前期(芽鳞伸出12~18 d)(图3)。结合各个时期文冠果花序特征将Ⅰ期和Ⅱ期称为发育前期,Ⅱ期和Ⅲ期称为发育中期,Ⅲ期和Ⅳ期称为发育后期(表1)。

表1 文冠果花序不同时期特征

图3 文冠果花序发育图
2.3 文冠果不同位置花蕾体视显微镜和扫描电镜观察
文冠果雌蕊各部分在Ⅰ期的发育是由外向内的向心式分化。从花原基开始分化后至Ⅱ期前,顶花序的两性花、侧花序的雄花以及转化花序的两性花雌蕊在形态上未见明显差别。顶花序两性花的8枚雄蕊同时轮状发育向上整齐包裹住由3个心皮组成的复雌蕊(图4A),可以看出柱头相邻两个心皮之间有一凹沟,各个心皮独立且分离(图4-B),此时心皮细胞很小,排列致密(图4C)。侧花序雄花此时8枚雄蕊同时发育整齐,包裹住由3个心皮组成的复雌蕊(图4D),心皮独立,分离(图4F),雄花子房大小与两性花一致(图4E)。转化花序两性花中3个心皮组成的复雌蕊由8枚雌蕊整齐包住(图4G),各个心皮是独立且分离的,排列紧密(图4I),子房大小与顶花序大小一致(图4H)。

图4 文冠果不同位置花序雌蕊Ⅰ期体视显微镜和扫描电镜照片
在大孢子母细胞期即Ⅱ期,雌、雄两器官的发育开始出现差异。顶花序两性花雌蕊的发育晚于雄蕊(图5A),8枚雄蕊已有花药形状,并且高于雌蕊。雌蕊从原来的短柱型逐渐变成三棱型向上生长,3个心皮逐渐愈合形成复雌蕊,心皮之间的凹沟细胞发育形成乳突状细胞(图5C),子房开始膨大,表面密布短毛(图5B)。侧花序雄花发育比顶花序两性花发育晚,雄蕊中的花药生长缓慢,发育不完全,花柱向上生长不明显(图5D),子房膨大(图5E),但雌蕊3个心皮不向中心愈合生长,心皮之间没有形成乳突状细胞(图5F)。转化花序的两性花生长类似顶花序,雌蕊发育晚于雄蕊发育(图5G),8枚雄蕊形成花药形态,雌蕊心皮呈三棱形向中心愈合形成复雌蕊,花柱伸长,心皮之间的细胞形成乳突状细胞(图5I),子房直径和长度逐渐增加(图5H),表面有短毛密生。
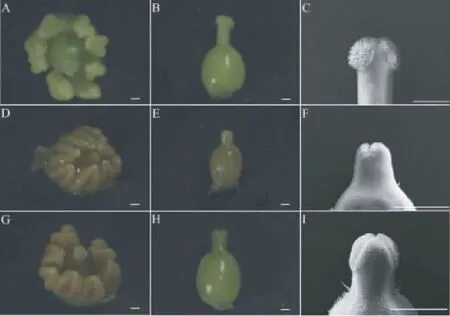
图5 文冠果不同位置花序雌蕊Ⅱ期体视显微镜和扫描电镜照片
在性别分化的关键期即大孢子母细胞减数分裂Ⅰ期,雌、雄两种生殖器官已经有明显的差异。从外部形态上看,顶花序两性花雌蕊的发育超过雄蕊的发育,花柱逐渐伸长,雄蕊花药生长缓慢(图6A),子房继续膨大,表面有短绒毛密生(图6B),柱头心皮之间的凹沟处乳突状细胞继续生长(图6C)。侧花序雄花雄蕊发育加快,8枚雄蕊发育饱满,花丝开始伸长,子房开始败育,3个心皮组成的复雌蕊中间开始出现空洞(图6D;图6E),心皮中心开始出现空洞(图6F)。转化花序两性花子房与顶花序两性花子房相似,子房体积不断膨大,表面也出现短绒毛密生(图6H,I),雄蕊发育不及雌蕊发育(图6G)。

图6 文冠果不同位置花序雌蕊Ⅲ期体视显微镜和扫描电镜照片
在性别分化完成期即大孢子母细胞减数分裂Ⅱ期前期,雌雄两器官已经分化完成。顶花序两性花的柱头高度逐渐超出雄蕊,子房体积继续膨大,表面绒状物密生,花柱继续伸长,心皮之间的乳突状细胞逐渐伸长,布满整个柱头(图7A;图7B;图7C)。侧花序雄花花柱并未伸长,子房已经萎缩,中间3心皮组成的复雌蕊中心空洞变大,8枚花药发育完全(图7D;图7E;图7F)。转化花序两性花的子房比前一个时期的体积要大,花柱继续伸长,心皮表面布满乳突状细胞,子房表面毛状物伸长(图7G;图7H;图7I)。

图7 文冠果不同位置花序雌蕊Ⅳ期体视显微镜和扫描电镜照片
2.4 文冠果雌蕊发育过程中糖含量的变化
不同位置花序雌蕊内的糖含量测定结果显示(图8),在文冠果雌蕊发育前期,总糖含量呈现缓慢增加趋势。在发育前期,顶花序、侧花序和转化花序雌蕊中的总糖含量差异不显著。在Ⅱ期向Ⅲ期生长转变的过程中,顶花序和转化花序雌蕊中的总糖含量开始显著增加,与侧花序差异显著,发育后期转化花序雌蕊中的总糖含量最高(图8A)。
顶花序和转化花序雌蕊中蔗糖含量的变化趋势相似,在雌蕊发育的整个过程中含量不断增加,发育前期转化花序雌蕊蔗糖含量高于顶花序,发育后期顶花序雌蕊蔗糖含量逐渐高于转化花序,在Ⅳ期差异显著。侧花序雌蕊在前三个时期的蔗糖含量缓慢增加,在Ⅳ期含量发生下降,含量始终最低,在发育后期与顶花序和转化花序差异显著(图8B)。
顶花序、侧花序和转化花序雌蕊在前三个时期果糖含量不断增加,发育前期三者差异不显著,在Ⅲ期顶花序雌蕊果糖含量显著高于侧花序和转化花序。在Ⅳ期顶花序和侧花序雌蕊果糖含量均有所降低,转化花序雌蕊果糖含量增加,显著高于顶花序和侧花序(图8C)。
顶花序和转化花序雌蕊中葡萄糖含量的变化趋势相似,前三个时期含量不断增加,在Ⅳ期含量有所降低,在整个雌蕊发育过程中两者差异均不显著。侧花序雌蕊葡萄糖含量在发育前期不断增加,Ⅰ期含量显著高于顶花序和转化花序,发育后期含量不断降低,并显著低于顶花序和转化花序(图8D)。
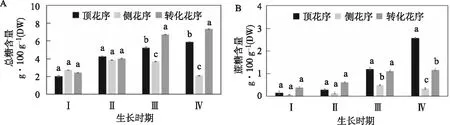
图8 文冠果不同位置花序雌蕊发育过程中糖含量变化
2.5 文冠果不同位置雌蕊发育过程中糖代谢相关酶的变化
不同位置花序雌蕊糖代谢相关酶活性的变化显示(图9)。在整个雌蕊发育过程中,顶花序和转化花序雌蕊SS合成方向酶活性在不断增加,侧花序雌蕊SS合成方向酶活性变化不大。在发育前期,侧花序雌蕊SS合成方向酶活性最高,与转化花序差异显著。在发育后期,顶花序雌蕊SS合成方向酶活性最高,转化花序次之(图9A)。由图9B可知,顶花序和转化花序雌蕊中SS分解方向酶活性的变化趋势相似,总体呈下降。侧花序雌蕊中SS分解方向酶活性不断增加,在发育前期酶活性低于顶花序和转化花序,但在发育后期显著高于顶花序和转化花序。由图9C可知,顶花序和转化花序雌蕊中SPS酶活性在整个雌蕊发育过程中不断增加,侧花序雌蕊中SPS酶活性呈现S型。在发育前期三者之间差异不显著,在发育后期,顶花序和转化花序雌蕊SPS酶活性显著高于侧花序。

图9 文冠果不同位置花序雌蕊发育过程中酶活性变化
2.6 文冠果不同位置雌蕊发育过程中糖含量与酶活性相关性分析
文冠果顶花序雌蕊生长发育的过程中,在发育前期,蔗糖的积累与SS合成方向活性显著负相关,r值为-0.969;果糖的积累与SS合成方向活性显著正相关,r值为0.867;葡萄糖的积累与SPS活性显著正相关,r值分别为0.971,与SS合成方向活性显著负相关,r值分别为-0.970;在发育中期,蔗糖的积累与SS分解方向活性显著负相关,r值为-0.911;果糖的积累与SS分解方向活性显著负相关,r值为-0.842;葡萄糖与SS合成方向活性显著正相关,r值为0.733。在发育后期,蔗糖的积累与SS合成方向、SPS活性显著正相关,r值分别为0.937、0.940,与SS分解方向活性显著负相关,r值为-0.831;果糖积累与各个时期都不显著;葡萄糖积累与SPS活性显著正相关,r值为0.879(表2)。

表2 文冠果顶花序雌蕊可溶性糖含量与酶活性相关性
文冠果侧花序雌蕊生长发育的过程中,在发育前期,蔗糖的积累与SS分解方向活性显著正相关,r值为0.733;在发育中期,葡萄糖的积累与SPS活性显著正相关,r值为0.824。在发育后期,葡萄糖的积累与SS分解方向活性显著正相关,r值为0.879(表3)。

表3 文冠果侧花序雌蕊可溶性糖含量与酶活性相关性
文冠果转化花序雌蕊生长发育的过程中,在发育前期,葡萄糖的积累与SPS活性显著正相关,r值为0.970,与SS分解方向活性显著负相关,r值为-0.815。在发育中期,蔗糖、果糖、葡萄糖的积累与各个糖代谢相关酶相关性不显著。在发育后期,蔗糖的积累与SS合成方向和SPS活性显著正相关,r值分别为0.820、0.907;果糖的积累与SS分解方向活性显著负相关,r值为-0.909(表4)。

表4 文冠果转化花序雌蕊可溶性糖含量与酶活性相关性
2.7 文冠果不同位置花序子房膨大率与糖含量的关系
由表5可知,顶花序子房膨大率在发育中期与总糖含量显著正相关,r值分别为1.000、0.998,在Ⅰ期和Ⅲ期与蔗糖含量显著正相关,r值分别为1.000、1.000,在Ⅳ期与果糖含量显著正相关,r值为0.997,各个时期的葡萄糖含量与子房膨大率没有显著相关。这说明在顶花序雌蕊糖代谢中,总糖和蔗糖含量对顶花序子房膨大率的调控最显著,主要表现在Ⅲ期。

表5 不同位置花序子房膨大率与糖含量相关性分析
侧花序子房膨大率在Ⅰ、Ⅱ和Ⅳ期与总糖含量显著正相关,r值分别为0.998、1.000和0.999,在Ⅳ期与蔗糖含量显著正相关,r值为0.999,在Ⅱ期与果糖含量显著正相关,r值为0.998,在Ⅰ、Ⅱ和Ⅳ期与葡萄糖含量显著正相关,r值分别为0.998、0.999和0.998。说明糖代谢中总糖和葡萄糖含量对侧花序子房膨大率的调控最明显,主要表现在发育前期。
转化花序子房膨大率在Ⅰ期与总糖含量显著相关,r值为1.000,在Ⅰ、Ⅱ和Ⅲ期与蔗糖含量显著相关,r值分别为1.000、1.000和1.000,在Ⅱ期与果糖含量显著相关,r值为1.000,在发育前期和Ⅳ期与葡萄糖含量显著相关,r值分别为0.999、1.000和1.000。说明去顶处理促进了蔗糖等糖类的含量增加,进而促进糖类积累,提高了雌花比例。
3 讨论
3.1 文冠果雌蕊的发育是选择性败育的结果
植物功能性雄蕊和雌蕊的发育是一个十分复杂的生理学过程。在发育前期,花的内部既有雌蕊原基也有雄蕊原基,在形态上没有任何差别。之后伴随着雄性或者雌性器官发育阻滞(Developmental Arrest),选择性败育分化成不同类型的花,行使不同功能[22-23],如在白麦瓶草(Silenelatifolia)[24]和黄瓜(Cucumissativus)[25]等作物中均存在这种现象。本研究中文冠果的两性花和雄花在发育早期都经历了一个“两性花”阶段(Bisexual Stage),形态和子房的直径和长度没有差别,发育后期分别分化形成了提供花粉的雄花及接收花粉的两性花。花器官选择性败育阶段在不同植物中不尽相同,可分为四个阶段:雌雄原基发育前,雌雄蕊发育初期,减数分裂前和减数分裂后[24]。研究发现,文冠果雄花中雌雌蕊选择性败育发生的阶段在大孢子母细胞减数分裂Ⅰ的双线期[5, 14-15],外部形态是蕾期向开花初期过渡的阶段[26-27],与本试验中的研究结果一致。
植物雌蕊的发育过程由雌雄性别决定和配子体的分化、发育和成熟组成。大量研究结果表明,雌蕊的发育与特定基因选择性表达、激素调控、miRNA的调控、程序性细胞凋亡(Programmed cell death,PCD)和细胞周期的停滞等复杂过程密切相关[28-30]。研究发现,雌蕊在性别决定过程中容易受到植物激素、营养、光照、温度和湿度等因素影响,表明植物发育过程中的性别分化具有一定可塑性。本试验发现文冠果雌蕊的败育发生在发育后期,此时两性花和雄花已经分化完成,但在短时间内迅速败育,说明侧花序雌蕊的败育涉及到发育受阻以及后期的结构解体等复杂过程,与PCD可能有密切联系,为后续从分子角度剖析机理打下了基础。
3.2 去顶处理显著提高了雌蕊内部糖含量以及花序子房膨大率
植物顶端优势使植物主茎顶端的生长占优势地位,而邻近侧芽的生长受到抑制,使侧芽处于休眠状态。本试验中在文冠果蕾期性别分化前去除顶花序,导致约35% 的两性花出现在临近原顶花序的第一侧花序上。油橄榄中存在与文冠果相似的情况,顶花序均为两性花,一级分枝很少或没有两性花,应与糖的供给等营养因素有关[31]。不同位置营养供给的差异而导致两性花和雄花的产生。也有研究认为,这种现象与激素调控密切相关[32]。糖与各种激素相互影响,共同调控植物不同位置的花芽发育。Mason等[33]的研究结果表明有效糖与顶端优势强相关联,而不是IAA。顶芽对糖的强烈需求限制了有效糖转运至腋芽,从而抑制腋芽萌发[33]。本试验中发现去顶处理能显著提高蔗糖含量与子房膨大率,将糖含量、顶端优势以及子房膨大率紧密联系为一体,为研究糖和激素在子房膨大率的作用提供了依据。
