不同土壤水分下大豆根冠互作对产量和农艺性状的影响
2020-07-28李盛有董丽杰王雅珍曹永强闫春娟王文斌宋书宏
李盛有,董丽杰,王雅珍,曹永强,闫春娟,王文斌,宋书宏
(辽宁省农业科学院作物研究所,辽宁 沈阳 100161)
大豆是我国最重要的经济作物之一,在我国农业生产中占有极其重要的地位。从当前大豆产业发展形势来看,干旱频发和水资源日益减少是限制大豆产量可持续增长的关键环境胁迫因子[1-2]。选育耐旱型品种对于保证大豆产量稳定具有重要意义。
大豆的生长发育是一个地上部与根系相互影响、相互促进的过程[3-5]。土壤缺水条件下,大豆植株根系与地上部间的相互调节对抵御水分胁迫起到了重要作用。研究表明,适度干旱胁迫限制了地上部植株的生长,但促进了早期根系的生长来吸收更多的水分,维持较高的水分利用效率,从而并未带来大豆产量的大幅降低[6-7]。但若在营养生长阶段遭受严重和长期干旱胁迫,会导致大豆植株停止生长,干物质生产严重受阻,最终也会带来减产[8-9]。大豆对干旱胁迫最敏感的时期是在大豆生殖生长阶段[6]。其中,开花结荚期水分胁迫能大大降低单株有效荚数,鼓粒始期水分胁迫则会降低粒重,两个不同生长时期产量分别降低18%和22%[10]。
近年来,国内研究者通过产量、农艺性状和生理指标对不同大豆种质资源的抗旱性进行了精准鉴定,筛选出一批耐旱型材料[11-15]。然而,由于根冠互作的存在,以往研究无法明确这些材料的耐旱性是得益于地上部性状的贡献还是根系的贡献,亦或是二者共同的作用。本研究以不同耐旱型品种进行相互嫁接,明确结荚期干旱胁迫下大豆根冠互作对产量和农艺性状的影响,从而为大豆抗旱育种提供理论指导。
1 材料与方法
1.1 试验材料
选用辽宁省农业科学院选育的耐旱型大豆品种辽豆14(L14)和干旱敏感型大豆品种辽豆21(L21)作为试材[16],于子叶期采用劈接法对大豆植株进行嫁接,具体嫁接方法参照Pantalone等[17]。嫁接处理包括2个不嫁接处理L14和L21,以及4个相互嫁接处理L14/L14、L21/L21、L14/L21和L21/L14(表1)。其中,L14/L14和L21/L21分别为自身嫁接植株,L14/L21为L14接穗嫁接至L21砧木,L21/L14为L21接穗嫁接至L14砧木。
1.2 试验设计
试验于2018—2019年连续2年在辽宁农业科学院试验基地自动抗旱棚内进行。盆栽用土来自大田,土壤理化性质为有机质含量22.30 g·kg-1,总氮含量1.91 g·kg-1,速效氮含量0.09 g·kg-1,速效磷含量0.01 g·kg-1,速效钾含量0.11 g·kg-1,pH为6.8,田间持水量为26.5%。待幼苗期嫁接植株成活后(成活率>90%),将嫁接植株和不嫁接植株统一移栽至室外盆栽(25 cm×20 cm×30 cm塑料桶,12.5 kg土)中。每盆定苗2株,随机区组排列,3次重复,各试验处理见表1。于结荚期开始控水处理,持续20 d,而后开始恢复正常供水。供水处理(WW)使土壤含水量保持在田间持水量的80%,水分胁迫处理(WS)使土壤含水量保持在田间持水量的50%。每天用称重法测定土壤含水量,补水至设定值。
1.3 测定项目
于成熟期收获地上部植株并进行室内考种,测定株高、主茎节数、主茎干重、分枝数、分枝干重、荚数、瘪荚率、荚重、粒数和粒重。测定荚数、荚重、粒数和粒重时,分别将主茎与分枝分开进行,并且测定主茎每节的有效荚数。每个处理测定3次重复。
1.4 数据统计
数据分析采用SPSS 17.0软件进行。采用一般线性模型进行分析,包括2个水分处理、6个嫁接处理和2年试验。其中,水分处理、嫁接处理和水分处理×嫁接处理作为固定因子,水分处理×年份、嫁接处理×年份和水分处理×嫁接处理×年份为随机因子。多重比较采用最小显著差异法。
2 结果与分析
2.1 不同土壤水分下大豆根冠互作对株型性状的影响
方差分析表明,株高、主茎节数和分枝数在不同嫁接植株间存在极显著差异(P<0.01),在不同水分处理间无显著差异(P>0.05),但分枝数在水分处理与基因型间存在显著的互作效应(P<0.05)。总体来看,自身嫁接植株与不嫁接植株间株高、主茎节数和分枝数无明显差异(表1)。水分胁迫使L14/L21、L21/L21 和L21/L14的分枝数分别降低了24.5%、26.8%和20.7%,而使L14/L14的分枝数提高了10.3%。株高和主茎节数主要受到接穗基因型影响。在正常供水和水分胁迫下,与L21/L21相比,L14/L21的株高分别提高了22.6%和10.0%,主茎节数分别提高了22.0%和15.9%,分枝数分别提高了88.9%和94.6%;与L21/L14相比,L14/L14的株高分别提高了25.9%和19.8%,主茎节数分别提高了17.2%和14.8%,分枝数分别提高了34.6%和87.2%;相比于L21接穗,L14接穗的株高分别平均提高24.3%和14.8%,主茎节数分别平均提高19.6%和15.3%,分枝数分别平均提高60.2%和90.6%。

表1 不同大豆嫁接植株株高、主茎节数和 分枝数对水分胁迫的响应
2.2 不同土壤水分下大豆根冠互作对干物质积累与分配的影响
方差分析表明,主茎干重、分枝干重、主茎荚干重、分枝荚干重和地上部干重在不同嫁接植株间存在极显著差异(P<0.01),在不同水分处理间存在极显著差异(P<0.01),并且在水分处理与基因型间存在显著的互作效应(P<0.05)。由图1可见,自身嫁接与不嫁接植株间主茎干重、分枝干重、主茎荚干重、分枝荚干重和地上部干重无显著差异。水分胁迫使大豆植株地上部干重显著降低,但不同嫁接植株的下降程度存在差异。其中,L21/L21的下降幅度较大,主茎干重、分枝干重、主茎荚干重、分枝荚干重和地上部干重分别下降了19.7%、11.3%、23.7%、55.1%和33.2%,而L14/L14、L14/L21和L21/L14下降幅度较小,主茎干重、分枝干重、主茎荚干重、分枝荚干重和地上部干重分别平均下降了4.0%、2.3%、27.4%、23.4%和17.8%。在正常供水和水分胁迫下,与L21/L21相比,L14/L14地上部干重分别显著提高了55.9%和92.1%,其中,分枝荚重的提高幅度大于主茎荚重,分枝荚重分别显著提高了56.7%和186.4%;与L21/L21相比,L14/L21地上部干重分别显著提高了58.3%和82.0%,其中,分枝荚重的提高幅度大于主茎荚重,分枝荚重分别显著提高了52.7%和164.9%;与L21/L21相比,L21/L14地上部干重分别提高了7.8%和41.5%,其中,主茎荚重的提高幅度大于分枝荚重,主茎荚重分别显著提高了35.9%和57.3%。
2.3 不同土壤水分下大豆根冠互作对荚数的影响
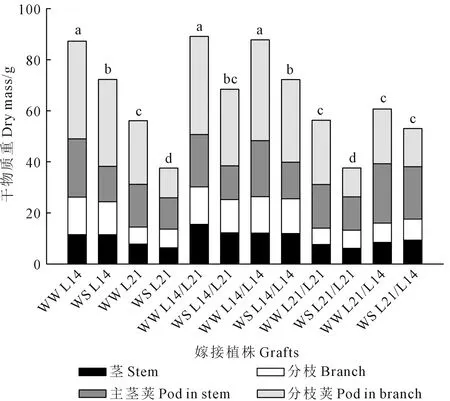
注:不同小写字母表示嫁接处理×水分处理下差异显著(P<0.05)。Note: Different letters mean the significant difference among grafting × water treatment interaction (P<0.05).图1 不同大豆嫁接植株各组织干物质重对水分胁迫的响应Fig.1 Response of dry mass of each tissue on different soybean grafts to water stress
方差分析表明,主茎荚数、分枝荚数、单株荚数、主茎瘪荚率、分枝瘪荚率和单株瘪荚率在不同嫁接植株间存在极显著差异(P<0.01),在不同水分处理间存在极显著差异(P<0.01),并且在水分处理与基因型间存在显著的互作效应(P<0.05)。由表2可知,水分胁迫降低了大豆的主茎荚数、分枝荚数和单株荚数,提高了主茎瘪荚率、分枝瘪荚率和单株瘪荚率。其中,L21/L21的变化幅度较大,主茎节数、分枝荚数和单株荚数分别降低了22.5%、16.3%和19.8%,主茎瘪荚率、分枝瘪荚率和单株瘪荚率分别提高了122.1%、84.5%和107.0%;而L14/L14、L14/L21和L21/L14变化幅度则较小,主茎节数、分枝荚数和单株荚数分别平均降低了17.0%、2.4%和8.5%,主茎瘪荚率、分枝瘪荚率和单株瘪荚率分别平均提高了55.9%、57.8%和54.7%。在正常供水和水分胁迫下,与L21/L21相比,L14/L14单株荚数分别显著提高了73.9%和96.1%,主要得益于分枝具有较多的荚数和较低的瘪荚率,分枝荚数分别提高了159.4%和201.4%,分枝瘪荚率分别降低了57.0%和58.0%;与L21/L21相比,L14/L21单株荚数分别显著提高了66.1%和88.5%,也主要得益于分枝具有较多的荚数和较低的瘪荚率,分枝荚数分别提高了160.4%和194.4%,分枝瘪荚率分别降低了58.6%和62.8%;与L21/L21相比,L21/L14单株荚数分别显著提高了10.0%和27.4%,主要得益于主茎具有较多的荚数和较低的瘪荚率,主茎荚数分别提高了18.5%和32.3%,主茎瘪荚率分别降低了22.0%和48.6%。

表2 不同大豆嫁接植株荚数和瘪荚率对水分胁迫的响应
进一步分析表明,不同嫁接植株主茎荚的分布特点存在明显差异(图2)。L14、L14/L14和L14/L21的主茎荚分布于第5节位至第25节位,平均每个节位2个荚;而L21、L21/L21和L21/L14的主茎荚分布于第4节位至第20节位,但主要集中分布于第6节位至第15节位,平均每个节位3.9个荚。虽然砧木基因型并未改变主茎荚的分布特征,但会影响每节位着荚数量。与L21/L21相比,L21/L14正常供水下平均每个节位增加了0.5个荚,而水分胁迫下平均每个节位增加了0.7个荚,其中,在第9节位至第15节位间,平均每个节位增加了1.6个荚。

图2 不同大豆嫁接植株主茎荚数的分布Fig.2 Distribution of number of pod on stem of different soybean grafts
2.4 不同土壤水分下大豆根冠互作对产量相关性状的影响
方差分析表明,单株粒数、百粒重和单株粒重在不同基因型间存在极显著差异(P<0.01),在不同水分胁迫处理间存在极显著差异(P<0.01),并且在水分处理与基因型间存在极显著的互作效应(P<0.01)。由表3可知,自身嫁接与不嫁接植株间单株粒数、百粒重和单株粒重无显著差异。水分胁迫降低了大豆的单株粒数、百粒重和单株粒重,其中,L21/L21的下降幅度较大,单株粒数、百粒重和单株粒重分别显著下降了36.4%、15.6%和46.4%,而L14/L14、L14/L21和L21/L14下降幅度较小,单株粒数、百粒重和单株粒重分别平均下降了25.9%、6.60%和31.3%。在正常供水和水分胁迫下,与L21/L21相比,L14/L14的单株粒数分别提高了57.5%和87.8%,单株粒重分别提高了21.1%和64.6%,粒数和粒重的提高幅度在主茎与分枝间无差异;与L21/L21相比,L14/L21的单株粒数分别提高了52.7%和67.4%,单株粒重分别提高了18.6%和45.2%,粒数和粒重的提高幅度在主茎与分枝间也无差异;与L21/L21相比,L21/L14的单株粒数分别提高了0.8%和21.7%,百粒重分别提高了0.9%和5.2%,单株粒重分别提高了1.6%和20.4%,主要得益于主茎着粒数和粒重的提高,主茎粒数分别提高了27.3%和37.5%,主茎粒重分别提高了28.4%和44.7%。

表3 不同大豆嫁接植株产量相关性状对水分胁迫的响应
3 讨论与结论
为了研究大豆地上部与根系间的相互作用,许多研究者将嫁接技术进行了诸多研究[17-18]。本研究通过嫁接试验表明,大豆地上部农艺性状和产量主要由地上部基因型决定,但水分胁迫下会受到根系基因型的较大影响。
大豆开花与结荚期遭遇干旱胁迫,产量下降幅度最大[19]。一方面主要是由于干旱引起的光合能力下降,碳水化合物供应能力不足[20]。不同耐旱基因型大豆的光合作用对水分胁迫的响应存在明显差异[21-22]。水分胁迫下光合作用的稳定与否是衡量大豆耐旱性的一个重要指标[23]。前期研究发现,水分胁迫下辽豆14叶片仍能维持较高的光合速率,主要归因于非气孔因素[24]。本研究表明,在不同土壤水分下,辽豆14接穗的荚重和植株生物量均显著高于辽豆21接穗,这与闫春娟等[16]研究结果基本一致。另一方面,土壤干旱引起根系脱落酸的大量合成,并积累于花荚中,最终导致落花率和落荚率的增加[25]。本研究发现,水分胁迫下辽豆14接穗与辽豆21接穗相比,分枝上具有较多的荚数和较低的瘪荚率,从而导致较高的单株荚数、单株粒数和单株粒重,分别显著增加了88.5%、67.4%和45.2%。由此说明,干旱胁迫下辽豆14叶片具有较强的光合物质生产能力,改善了同化物对花荚发育的供应水平,从而能够形成相对较多的单株粒数。
众所周知,大豆根系不仅具有养分和水分的吸收以及信号传递的功能,并且具有与其共生的根瘤固氮菌,在调控地上部组织的生长和发育中起着极为重要的作用[26-27]。根系的重要性虽早已为国内外研究者们所注意,但由于土壤限制了研究者对根系的观察和测定,对作物根系性状的相关研究远远滞后于对地上部性状的相关研究[28]。本研究表明,砧木基因型对大豆的株高、主茎节数和分枝数无显著影响,但对单株荚数、单株粒数、百粒重和单株粒重存在显著影响。Pantalone 等[17]研究表明,大豆抗旱种质资源PI416937作为砧木能够使其它基因型大豆接穗籽粒干重获得显著提高。前期研究发现,在正常供水条件下,辽豆14作为砧木在生育后期具有较高的根系生理活性,可以显著提高当代育成品种的光合速率[29],从而提高百粒重和单株产量[30]。在水分胁迫下,嫁接辽豆14砧木同样能够使辽豆21接穗的叶片相对含水量、光合速率、Rubisco羧化效率显著提高[24]。在本研究中,嫁接辽豆14砧木使水分胁迫下的辽豆21接穗的单株荚重、单株荚数、单株粒数、百粒重和单株粒重显著提高了45.8%、27.4%、21.7%、5.2%和20.4%。其中,荚粒数的提高主要得益于主茎上较多的有效荚数和较低的瘪荚率。进一步分析发现,辽豆21接穗嫁接辽豆14砧木后,主茎第9节位至第15节位间,平均每个节位增加了1.6个荚。因此,通过根系改良也是提高干旱下大豆产量的一条重要途径。
综上所述,在正常供水和水分胁迫下,与辽豆21相比,辽豆14接穗的株高平均提高24.3%和14.8%,主茎节数分别平均提高19.6%和15.3%,分枝数分别平均提高60.2%和90.6%,单株生物量平均提高57.1%和87.1%,单株荚数平均提高70.0%和92.3%,分枝荚数平均提高159.9%和197.9%,分枝瘪荚率平均降低57.8%和60.4%,最终单株粒重平均提高19.9%和54.9%。与辽豆21自身嫁接植株相比,水分胁迫下,嫁接辽豆14砧木使单株荚重、单株荚数、单株粒数、百粒重和单株粒重分别显著提高了45.8%、27.4%、21.7%、5.2%和20.4%,产量性状的提高与主茎第9~15节位间有效荚数的提高有关。因此,在品种选育过程中,在对地上部产量相关性状进行改良的同时,也要关注根系性状的选择,从而提高大豆品种的抗旱性。
