不同温度下秋茄树幼苗叶绿素荧光参数对光强的响应
2020-07-27郑春芳陈继浓康华靖
李 红,郑春芳,陈继浓,于 冯,康华靖
(1.温州市农业科学研究院,浙江 温州 325006;2.浙江省近岸水域生物资源开发与保护重点实验室,浙江省海洋水产养殖研究所,浙江 温州 325005)
红树林是国际《湿地公约》《生物多样性公约》《联合国海洋法公约》《中国湿地行动计划》和《中华人民共和国海洋环境保护法》等的重要保护对象之一[1]。它的高生产力、高归还率和高分解率功能给近海海洋生态系统提供了强大的物质基础,在防止海岸侵蚀、减缓海平面上升、减轻污染、海洋药物研发、科研教育、生态旅游等方面发挥着重要作用[2]。红树植物广泛分布于热带亚热带海岸潮间带地区,温度是影响其分布的最主要因素。温度是影响植物生长发育及其地理分布的一个重要生态因素,植物只有在适宜的温度环境下才能正常生长发育并顺利完成其生活史[3]。研究树种生境中温度等逆境胁迫对光合作用的影响及其与植物的生态分布和演替状态之间的联系,一直倍受关注[4-6]。
光合作用是绿色植物生长发育的基础。目前低温对红树植物光合作用的影响一直深受关注。杨盛昌等[7]发现,低温抑制红树植物叶片净光合速率、气孔导度、蒸腾速率和水分利用率,促进胞间CO2浓度增加。Kao等[8]认为,低温胁迫使红树植物叶片光系统II(PSII)反应中心电荷分离实际量子效率(ΦPSII)以及电子传递速率(ETR)降低,抑制了PSII活性和ETR,致使光合功能受损。秋茄树Kandelia obovata是一种典型红树植物,由于其适应性广,是我国亚热带海岸滩涂绿化应用最广的红树之一[9]。秋茄树属于耐寒红树植物之一,也是浙江省温州市红树林引种成林最北缘的唯一品种[10]。另外,杨娟等[11]研究了温度对相同引种地三个秋茄树居群遗传多样性的影响,结果表明低温导致了秋茄树遗传多样性的减少。为了缓解低温对红树的伤害,余敏等[12]探讨了外源NO缓解秋茄树低温胁迫的生理机制;郑春芳等[10]研究了烯效唑对低温胁迫下秋茄树幼苗光合作用与抗氧化系统的影响,认为烯效唑处理能减少低温胁迫对秋茄树叶片细胞的伤害。
鉴于秋茄树重要的生态功能和价值,人们一直致力于其向高纬度地区引种。秋茄树虽是众多红树植物中耐寒性最强的种类,但高纬度地区冬季极端低温天气会使得秋茄树遭受到冻害,甚至引起植株的死亡。因此,本文拟通过室内不同温度下的受控实验,研究秋茄树幼苗叶绿素荧光参数对光强(PAR)变化的响应,以期为高纬度地区秋茄树幼苗的繁育和造林提供理论依据。
1 研究方法
1.1 材料培养与试验设计
供试秋茄树为2年生移植容器苗,由浙江海洋水产养殖研究所提供。幼苗于2017年春季移植,统一种植在口径为25 cm,底直径为20 cm,高25 cm的塑料小水桶中。桶内装有3 kg自然风干的滩涂淤泥(NaCl含量为1.29 g·kg-1),每桶4株幼苗。于2018年1月置于温州市农业科学研究院实验室的培养箱(昼/夜温度为25/20℃,湿度为75%)中适应7 d,然后进行各试验处理,具体昼/夜温度分别设置为A(25/20℃),B(20/15℃),C(15/10℃),D(10/5℃)和E(5/0℃),昼、夜处理时间各为12 h,培养箱中PAR为400 µmol·m-2·s-1,湿度均设置为75%,由外置加湿器保证湿度的稳定,共处理28 d。每种处理3盆,每盆4株,常规管理,3次重复,完全随机排列。
1.2 测定内容与方法
测量前分别选取长势一致的植株各7株,每株上各选取一张叶片待测(由于E处理生长状况极差,叶片脱落严重,故不予测量)。此时秋茄树幼苗高度集中在35~ 40 cm,地径约1.5 cm。植株生长正常。所选叶片的叶色较绿,均为完全发育成熟且健壮的叶片。
利用便携式光合仪(LI-6400-40,LI-COR INC.,USA)于9:00-16:00进行数据测量。仪器参数设置参考文献[13],光梯度设置为1 800,1 600,1 400,1 200,1 000,800,600,400,200,150,100和50 μmol·m-2·s-1,并应用自动测量程序自动记录气体交换参数和叶绿素荧光参数,仪器可直接测量得到叶片的ETR,PSII反应中心的激发能捕获效率(Fv'/Fm'),ΦPSII,光化学猝灭系数(qP)和非光化学猝灭系数(qN)等参数对PAR的响应曲线。每次记录最小等待时间为60 s,最大等待时间为90 s。
1.3 数据处理
根据光合计算软件4.1(井冈山大学生命科学学院 数理学院),分别应用光响应机理模型(Model I)、直角双曲线模型(Model II)、非直角双曲线模型(Model III)和双指数方程模型(Model IV)进行拟合及计算[14]。SPSS19.0版本(SPSS,Chicago,IL)软件进行计算、处理和方差差异显著性分析,采用Excel 2010作图。
2 结果与分析
2.1 秋茄树ETR 对PAR 的响应
由图1可以看出,各处理秋茄树幼苗叶片的ETR在PAR<600 μmol·m-2·s-1时均先随PAR的升高而快速增大,随后,ETR上升较为平缓,并在1 400 μmol·m-2·s-1时达到最大,之后,随着PAR的增加而略有下降。从图1还可以看出,A和B处理下秋茄树幼苗叶片的ETR较为接近,整体上远高于C和D处理(P<0.05)。另外,C处理的ETR整体上也明显高于D处理(P<0.05)。
以A处理数据为例,分别应用Model I,Model II,Model III和Model IV进行拟合,其结果见图2。从图2可看出,除Model II不能很好地拟合高PAR下ETR下降的趋势外,其余模型均能较好地拟合测量值的变化趋势。从确定系数(R2)来看,4种模型的R2均高于0.9(表1)。
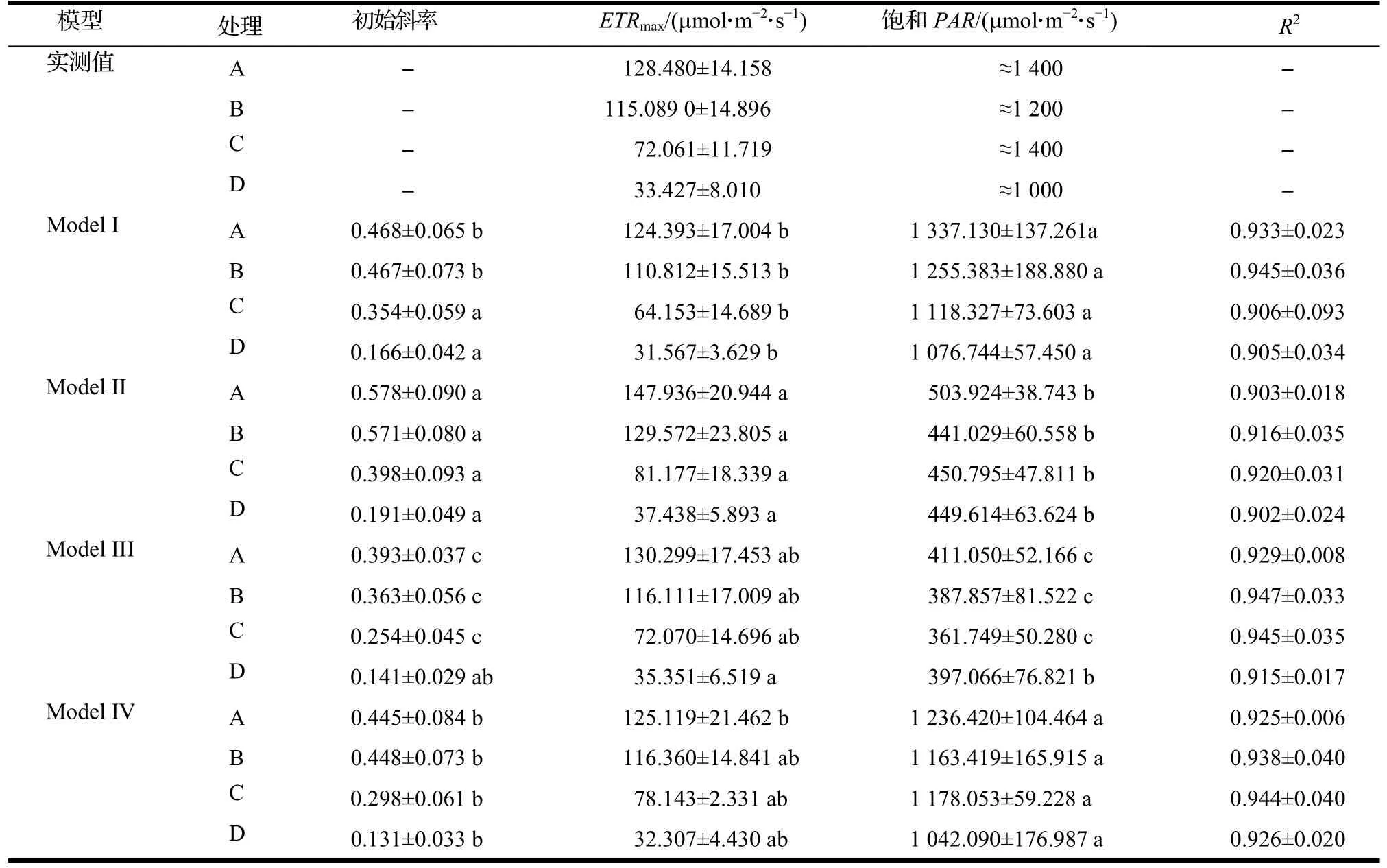
表1 ETR对光响应曲线的拟合结果 (平均数±标准误差)Table 1 Fitting of response curve of ETR to light
从拟合得到的最大电子传递速率(ETRmax)来看,Model I,Model III和Model IV拟合得到的结果较为接近,且与实测ETRmax较为吻合;虽然Model III拟合得到的值相对较高,但除A处理外,其余处理下的拟合值也与实测ETRmax间无显著差异(P>0.05)。然而,从拟合得到的饱和PAR来看,4种模型拟合的效果存在较大差异。整体来看,Model I和Model IV拟合得到的饱和PAR较高,前者拟合得到的饱和PAR分别为1 337.130(A),1 255.383(B),1 118.327(C)和1 076.744(D)μmol·m-2·s-1;后者拟合得到的饱和PAR则分别为1 236.420(A),1 163.419(B),1 178.053(C)和1 042.090(D)μmol·m-2·s-1,且与实测ETRmax所对应的PAR较为接近,远高于Model II和Model III拟合的结果(P<0.05)。由此判断,Model I和Model IV的拟合效果较好。另外,由初始斜率的拟合值来看,在同一模型下,A和B处理值较为接近,且远高于C和D处理(P<0.05)。
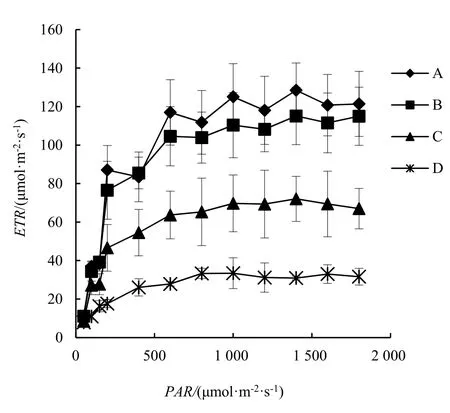
图1 不同温度处理下秋茄树幼苗叶片ETR 对PAR 的响应Figure 1 Response of ETR of different treated K.obovata on PAR
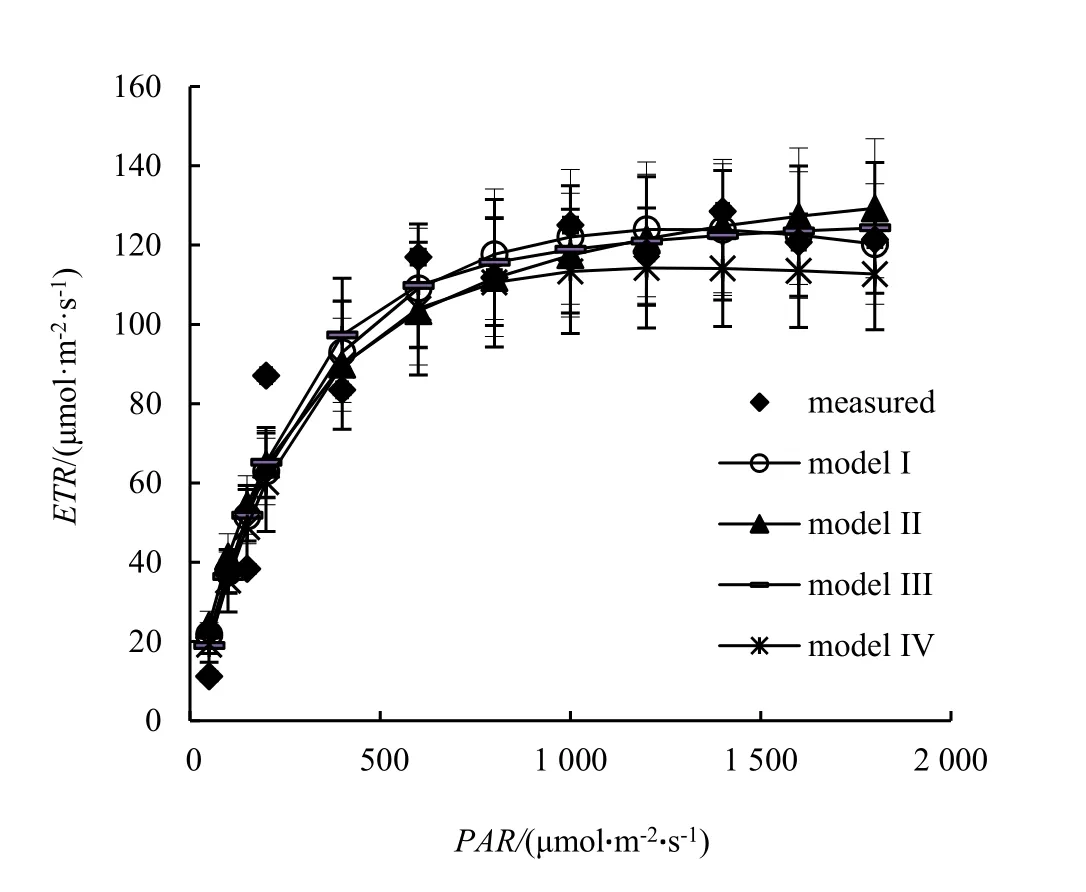
图2 Model I,Model II,Model III和Model IV对ETR-光响应曲线的拟合Figure 2 Fitting of 5 models and measure values of different treated K. obovata
2.2 秋茄树幼苗Fv'/Fm'和ΦPSII 对PAR 的响应
由图3 a可以看出,同一温度处理下,秋茄树幼苗的Fv'/Fm'随着PAR的升高而整体波动较小。比较不同温度下的Fv'/Fm'可以看出,在A和B处理下的Fv'/Fm'值极为接近,显著高于C和D处理(P<0.05)。由图3b可以看出,随着PAR的升高,秋茄树幼苗的ΦPSII逐渐降低。同样,比较不同温度下的ΦPSII可以看出,在A和B处理下的ΦPSII极为接近,显著高于C和D处理(P<0.05)。
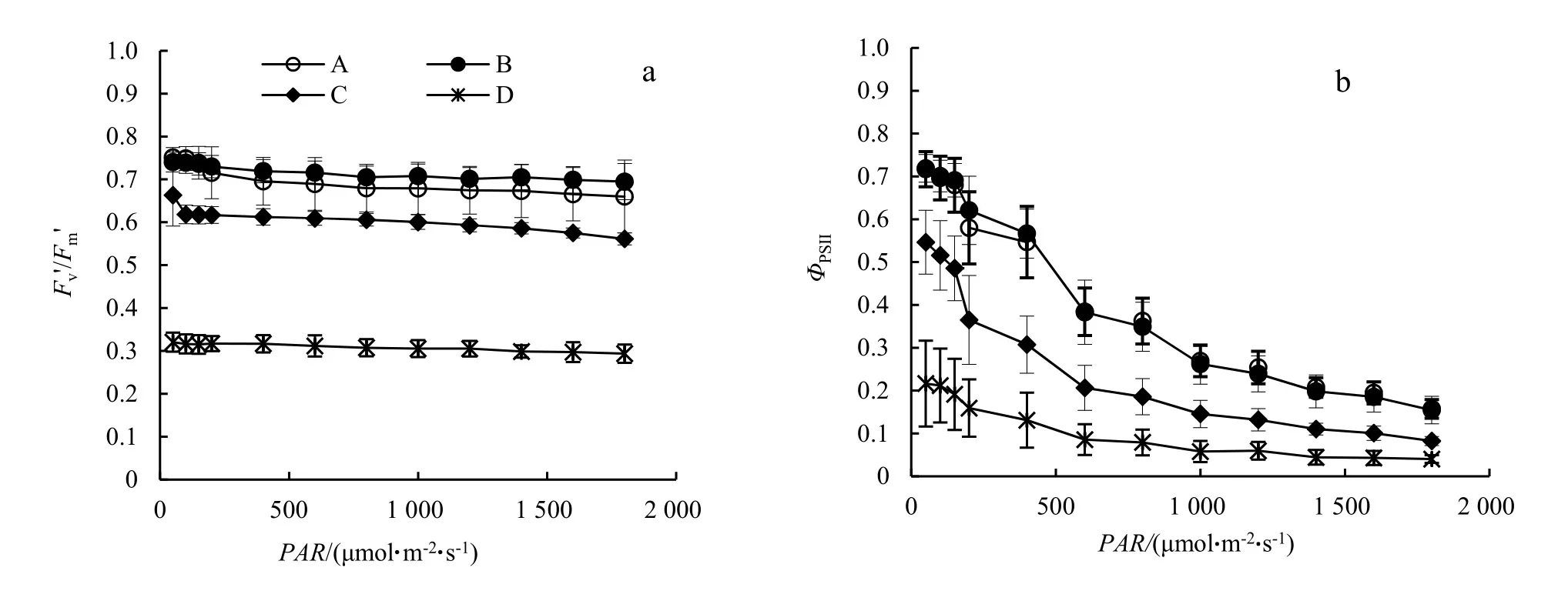
图3 不同温度下秋茄树幼苗实际Fv'/Fm'(a)和实际ΦPSI(b)对PAR的响应Figure 3 Fv'/Fm' and ΦPSII of different treated K. obovata
2.3 秋茄树幼苗qP 和qN 对PAR 的响应
由图4a可以看出,同一温度处理下,秋茄树幼苗光的qP随着PAR的升高而逐渐降低。比较不同温度下的Fv'/Fm'可以看出,随着处理温度的降低,秋茄树幼苗的qP也逐渐下降。由图4b可以看出,随着PAR的升高,秋茄树幼苗的qN整体波动较小;而比较不同温度下的qN可以看出,在A和B处理下的qN值极为接近,且显著高于C和D处理(P<0.05)。同样,在C和D处理之间的qN也存在显著差异(P<0.05)。

图4 不同温度下秋茄树幼苗qP(a)和qN(b)对PAR 的响应Figure 4 Light-response curves of qP and qN of different treated K.obovata
3 结论和讨论
叶绿素荧光动力学技术作为研究植物光合功能快速、无损伤的探针,在测定叶片光合同化过程中,光系统对光能的吸收、传递、耗散、分配等方面具有显著的优势。因此,本文基于荧光数据比较了不同温度处理秋茄树幼苗对不同PAR的响应。
本试验运用了4种模型对不同温度下秋茄树叶片的饱和PAR进行了探讨。结果显示,Model I拟合给出的饱和PAR分别为1 337.130(A),1 255.383(B),1 118.327(C)和1 076.744(D)μmol·m-2·s-1;Model IV拟合给出的结果分别为1 236.420(A),1 163.419(B),1 178.053(C)和1 042.090(D)μmol·m-2·s-1,且以上2种模型拟合结果与实测ETRmax对应的PAR较为接近(P>0.05)。由此可见,秋茄树幼苗叶片的饱和PAR范围为1 000~ 1 400 μmol·m-2·s-1,说明秋茄树幼苗表现出较强的阳生植物特性。
从ETRmax的拟合结果来看,4种模型的拟合结果整体上与实测ETRmax较为吻合。而A,B,C,D不同处理下的ETRmax之间差异显著(P<0.05),以光响应机理模型的拟合值为例,在A,B,C,D处理下,秋茄树叶片的ETRmax分别为124.393,110.812,64.153和31.567 mol·m-2·s-1,不同温度间ETRmax的值均达到显著差异(P<0.05)。可见,温度低于20℃对秋茄树幼苗的光合影响较大。这与秋茄树幼苗植株生长的外部形态观察结果较为一致。
Fv'/Fm'代表光合反应中心PSII的光合效率,反映了光合反应中心在部分关闭的情况下的实际捕获能量的传递效率[15]。同样,ΦPSII也可作为研究植物光抑制的一种快速的、非损伤性的指标[16]。本试验结果表明,在C和D处理下,秋茄树幼苗的Fv'/Fm'和ΦPSII显著降低(P<0.05)。qP反映PSII天线色素吸收的光能用于光化学电子传递的份额,并进一步反映PSII反应中心的开放程度和电子传递活性的大小[17-18]。从测量数据来看,在A和B处理下,秋茄树幼苗叶片的qP较为接近,均显著高于C和D处理(P<0.05),这说明温度高于20℃处理的秋茄树幼苗叶片PSII反应中心的开放程度和电子传递活性的大小基本稳定。植物吸收的光能一部分用于光合作用外,还有一部分可通过qN过程来调整过量能量的耗散,保护PSII反应中心免受因吸收过多光能而引起的光氧化和光抑制伤害[19]。比较不同温度下的qN可以看出,在A和B处理下的qN较高,显著高于C和D处理(P<0.05),说明较低温度下秋茄树幼苗叶片通过qN过程来调整过量能量耗散的能力较差。这与较低温度对红树的相关研究相一致[7-8]。郑春芳等[10]研究表明,烯效唑处理能改善低温胁迫下秋茄树植株抗氧化系统运行,减少低温胁迫对秋茄树叶片细胞的伤害。因此,在温州地区进行秋茄树的推广应用,在其幼苗阶段建议喷施植物生长调节剂以提高其抗寒性。
综合本试验结果表明,不同温度下的秋茄树幼苗均表现出阳生植物特性,幼苗叶片饱和PAR范围为1 000~1 400 μmol·m-2·s-1;温度低于20℃对秋茄树幼苗的光合影响较大;最低温度低于5℃将影响幼苗的生存。因此,建议在幼苗阶段,夏季进行适当遮阳,冬季喷施植物生长调节剂或采用其他措施进行保温。
