白菜bHLH转录因子家族的全基因鉴定及表达特征分析
2020-07-23唐文武吴秀兰钟佩桥
唐文武,吴秀兰,钟佩桥
(1.肇庆学院 生命科学学院,广东 肇庆 526061;2.肇庆学院 食品与药品工程学院,广东 肇庆 526061)
bHLH (basic helix-loop-helix)转录因子是一类含有bHLH结构域,在真核生物中广泛存在的转录因子超家族[1]。bHLH结构域一般由50~60个氨基酸组成,包括N端由10~15个碱性氨基酸组成的DNA结合区域,以及C端由约40个氨基酸组成的α螺旋-环-α螺旋区域(HLH)[2]。高等植物具有数目较多的各类bHLH转录因子,目前已在拟南芥中发现21个亚家族的147个bHLH转录因子[3,4],在水稻中鉴定出了22个亚家族的167个bHLH转录因子[5],它们在植物生长发育、形态建成、花器官形成、激素应答、次生产物代谢和抗逆性等方面发挥重要作用[6-8]。
白菜是重要的蔬菜作物,与拟南芥有着共同的祖先,是研究多倍体事件的经典材料[9]。bHLH是植物重要的转录因子,能通过调控花器官相关基因的表达而影响小孢子发育,与植物雄性不育发生密切相关。目前已在拟南芥、水稻、玉米等模式植物中发现bHLH转录因子在花药绒毡层发育、小孢子母细胞减数分裂,以及雄蕊发育等过程中起到重要作用,其基因突变或功能异常会引起植物雄性不育[10]。目前已在拟南芥、水稻中鉴定了AMS[11]、DYT1[12]、JAM[13]、UDT1[14]、TDR1[15]等与雄性不育相关的 bHLH基因,但在白菜中开展 bHLH转录因子的研究较少。为了深入挖掘与白菜细胞核雄性不育相关的bHLH转录因子,本研究利用生物信息学方法,对白菜bHLH转录因子家族的成员进行了全基因鉴定,对其bHLH结构域、进化关系进行了分析,并利用转录组数据对bHLH转录因子家族基因在可育和不育花蕾中的表达差异进行了分析,筛选了与雄性不育相关的bHLH基因,以期为进一步揭示白菜bHLH家族基因在雄蕊发育中的生物学功能和雄性不育应用提供参考。
1 材料与方法
1.1 白菜bHLH转录因子家族的获取与分析
从芸薹属BRAD基因组数据库(http://brassicadb.org/brad/)中下载白菜(B.rapa)全基因组序列(V1.5版),在Pfam数据库(http://pfam.xfam.org/)下载bHLH结构域(PF00010)的隐马尔可夫模型序列谱,并利用HMMEMIT软件获取其蛋白保守序列,用此序列检索白菜BRAD(http://brassicadb.org/brad/)和NCBI(http://www.ncbi.nlm.nih.gov/)中的蛋白质数据库;然后利用SMART(http://smart.embl-heidelberg.de/)进行保守结构域预测,并去除假阳性序列,获得白菜bHLH转录因子家族候选基因,用于后续分析。
1.2 白菜bHLH转录因子家族的生物信息学分析
利用ExPASy(http://web.expasy.org/)在线预测bHLH转录因子的氨基酸数量、分子量、等电点等基本理化特征。从PGDD数据库(http://chibba.agtec.uga.edu/duplication)获取基因复制及共线性分析数据,利用Circos软件对bHLH家族基因的染色体位置和共线性关系进行可视化分析[16]。利用clustal X对白菜bHLH转录因子的保守结构域进行比对,并利用Web Logo 3(http://weblogo.threeplusone.com/)进行可视化作图[17]。最后利用MEGA 7程序的邻接法构建白菜bHLH转录因子的系统进化树,其Bootstrap值设为1000。
1.3 白菜bHLH家族基因的育性及组织表达特异性分析
白菜基因的表达数据来源于NCBI中的GEO数据库,利用白菜细胞核雄性不育两用系中可育及不育花蕾的转录组数据[18],以FPKM 值作为衡量基因表达水平的指标,筛选bHLH转录因子家族基因在可育和不育花蕾中的差异表达序列,以log2倍数绝对值大于2为依据筛选差异表达的bHLH基因。
2 结果与分析
2.1 白菜bHLH转录因子家族鉴定及结构域分析
根据bHLH结构域PF00010的特征文件,在NCBI、BRAD的白菜数据库中鉴定到249个bHLH转录因子家族蛋白,按照其在BRAD数据库中的基因序号,分别命名为BraHLH001~BraHLH249。理化性质预测显示白菜bHLH转录因子家族成员的基因长度为1134~13231 bp,蛋白质氨基酸数目介于84~1201,分子量为9.80~132.38 kDa,理论等电点分布在4.69~9.81范围内。
进一步对249个白菜bHLH转录因子家族成员的bHLH结构域序列进行分析,发现bHLH结构域基序长度为41~66,平均氨基酸数目为49.8。用clustal X对所有的bHLH结构域进行多序列比对,然后采用WebLogo作图,结果(图1)显示白菜的bHLH结构域由50个氨基酸组成,其N端的Basic区含有植物中高度保守的His5-Glu9-Arg13序列,该序列对靶基因启动子上游的E-box识别结合必不可少;在C端的HLH区域,第23、49位的Leu高度保守,第39、43、46位的Leu/Ile/Val也非常保守,这些氨基酸残基对于二聚体的形成和功能非常重要。
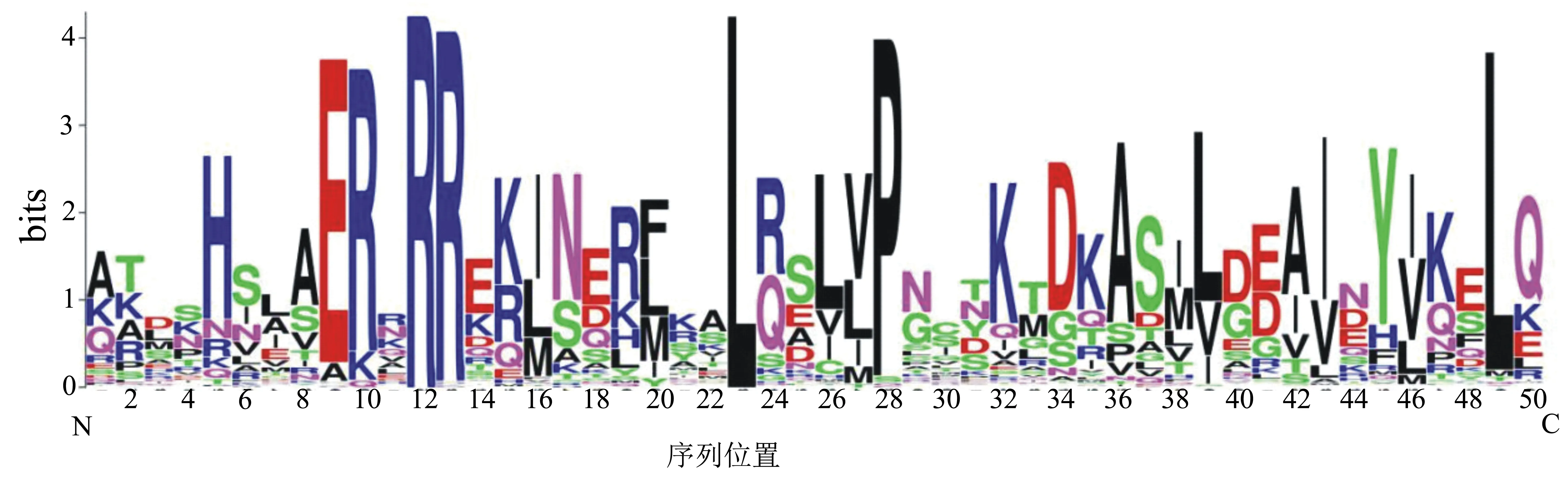
图1 白菜bHLH转录因子家族的保守结构域
2.2 白菜bHLH家族基因的染色体定位、基因复制和共线性分析
白菜bHLH转录因子家族基因的染色体定位结果(图2)显示,除BraHLH240、BraHLH247、BraHLH248、BraHLH249定位在scaffold上外,其余的245个基因不均匀地分布于白菜的10条染色体上,其中在A9染色体上的bHLH转录因子家族基因最多,达39个;其次是在A3染色体上,有33个基因;而在A8、A10染色体上分布的基因较少,分别仅含有16、14个基因;在其它6条染色体上分布有22~27个bHLH转录因子家族基因。
为了进一步确定白菜bHLH基因之间的关系,以及进化中潜在的重复事件,对249个白菜bHLH基因的旁系同源基因对进行共线性分析。结果(图2)表明,除了49个白菜bHLH基因没有同源重复外,其余200个bHLH基因均有对应的旁系同源基因,但多数旁系同源基因分布在不同染色体上,该结果可能是由白菜全基因组三倍化事件引起的。
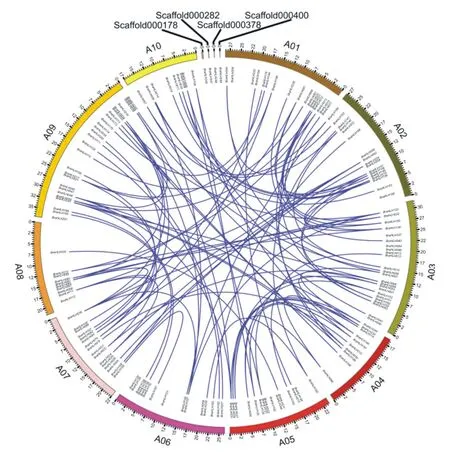
图2 白菜bHLH转录因子基因家族的共线性关系及其在染色体上的分布
2.3 白菜bHLH转录因子家族的系统进化分析
利用MEGA 7软件对鉴定出的249个白菜bHLH转录因子家族蛋白的bHLH结构域序列构建系统进化树,并以拟南芥的bHLH转录因子亚家族分类方法作为参考。系统进化分析结果(图3)表明,白菜中249个bHLH蛋白可被分为24个亚家族,其中XⅡ、Ⅰb (2)、Ⅰa三个亚族的bHLH成员最多,分别包含29、25、23个bHLH蛋白;第Ⅲ(a+c)、Ⅲb、Ⅲ(d+e)、Ⅶ(a+b)、Ⅷa、Ⅷc、Ⅴb、X、XV共9个亚族的bHLH蛋白成员为10~19个;第Ⅱ、Ⅲf、Ⅰb(1)、Ⅳa、Ⅳb、Ⅳc、Ⅴa、Ⅷb、IX、XI共10个亚族的bHLH成员为3~9个;而Ⅳd和Orphans亚族的成员最少,均只含有1个bHLH蛋白。
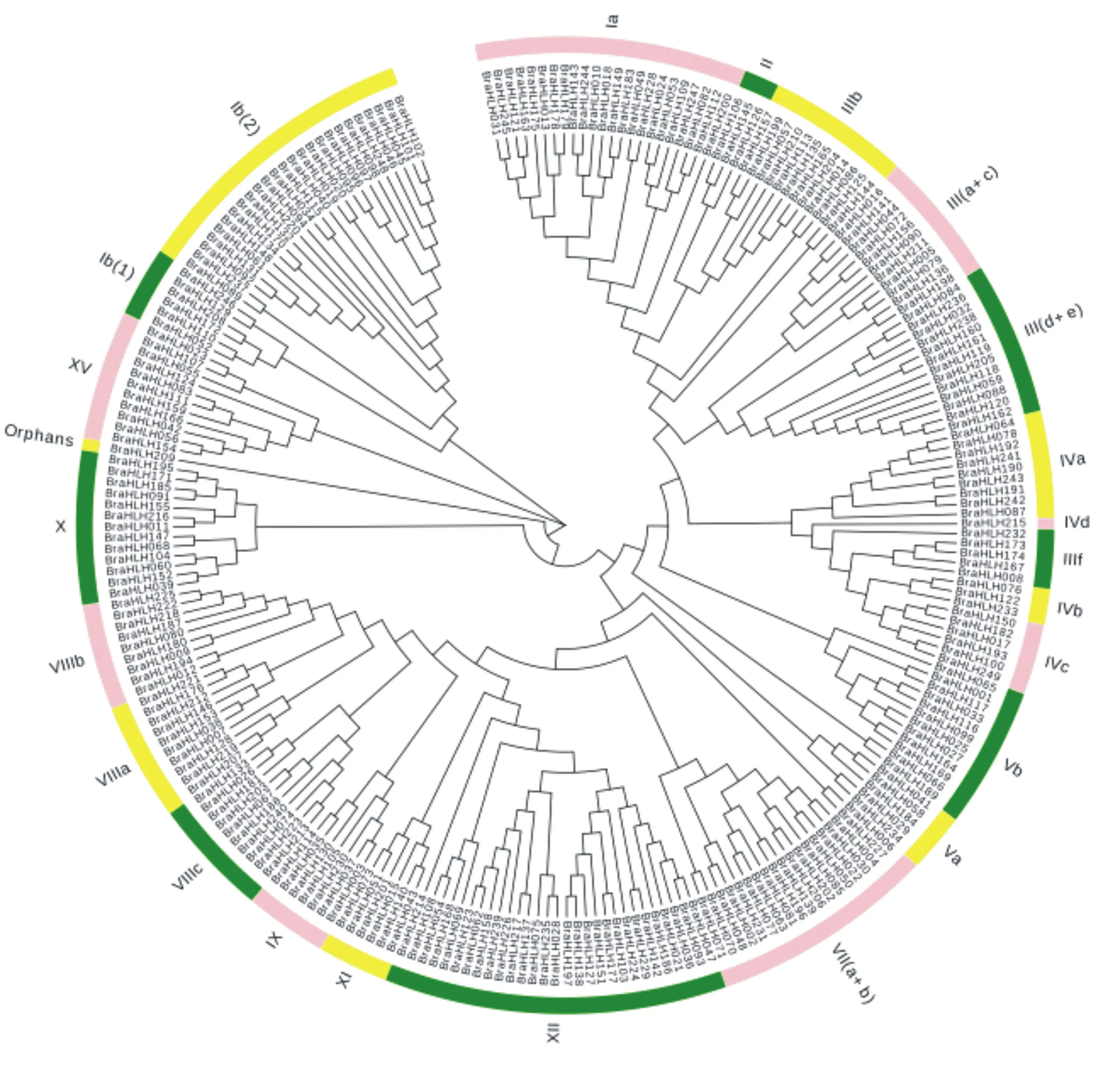
图3 白菜bHLH转录因子家族的系统发育进化树
2.4 白菜bHLH转录因子基因的育性表达差异分析
基于白菜核不育两用系中可育及不育花蕾的转录组数据,对249个bHLH转录因子家族基因在可育、不育花蕾中的表达差异性进行了分析,结果如表1所示,共有12个白菜bHLH转录因子基因在不育与可育花蕾中的表达量log2倍数值大于2。相较于可育花蕾,不育花蕾中有10个基因的表达量显著下调,另2个基因BraHLH191、BraHLH200的表达量显著上升。这12个表达差异的bHLH转录因子基因,可能与雄蕊不育性状相关。其中BraHLH034是拟南芥AMS的直系同源基因,可能参与花药绒毡层发育以及小孢子母细胞减数分裂的调控过程[11,19],而BraHLH106、BraHLH125与拟南芥的bHLH89、bHLH91为直系同源基因,通过与DYT1相互作用调控花药发育并引起雄性不育现象[20]。
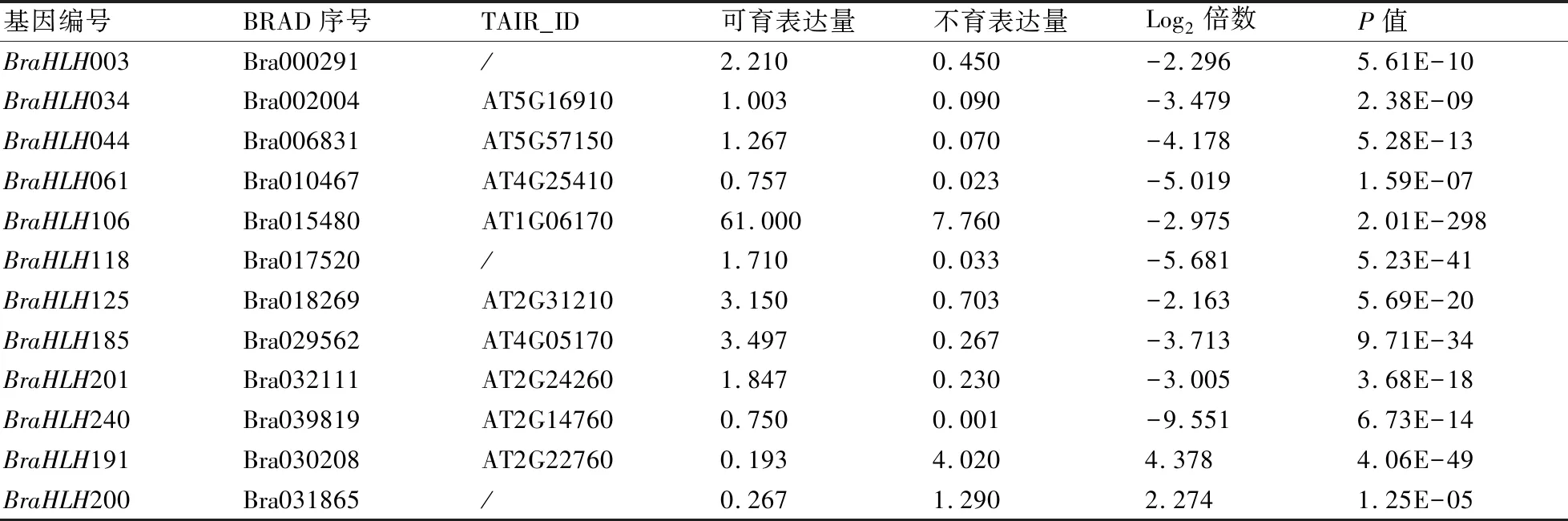
表1 白菜可育和不育花蕾中表达差异显著的bHLH转录因子基因
3 小结与讨论
bHLH转录因子是植物中数量最多的一类转录因子,在植物生长发育的各环节均发挥重要的调控作用。目前对拟南芥、水稻等模式植物的bHLH转录因子的研究比较深入,但对蔬菜类bHLH基因家族的研究较少。本研究利用生物信息学方法从白菜基因组中鉴定出249个bHLH转录因子,并发现其bHLH结构域由约50个氨基酸组成,其N端的Basic区含有高度保守的H5-G9-A13序列,可与启动子区的E-box结合;而C端第23、49位的亮氨酸高度保守,第43、46位的亮氨酸/异亮氨酸/缬氨酸也非常保守,这些保守位点与bHLH转录因子的功能密切相关。染色体定位分析结果显示245个白菜bHLH转录因子基因不均匀地分布于10条染色体上,多数基因有对应的旁系同源基因。根据bHLH结构域的特征,可将白菜的249个bHLH蛋白分为24个亚家族,比拟南芥和水稻的亚家族数目多,其中XⅡ是最大类的亚家族,包含29个bHLH成员。
在模式植物中发现bHLH转录因子与雄性不育的形成密切相关,bHLH转录因子会参与花药绒毡层的发育或小孢子发育,其功能异常会引起绒毡层细胞提前或延迟降解,致使花粉形成受阻而败育[10]。目前已在拟南芥中发现DYT1[12]、AMS[11]、MYC[21]等bHLH转录因子在花药绒毡层发育和小孢子母细胞减数分裂中起到重要作用,发现3个JAM基因参与茉莉酸信号的转导,并在雄蕊发育中起到负调控作用[13]。在水稻中发现了UDT1[22]、TDR1[23]、EAT1[24]、TIP2[25]等bHLH转录因子在绒毡层细胞发育和程序性死亡中发挥重要作用,其功能异常也会引起雄性不育。本研究对白菜可育、不育花蕾中的bHLH转录因子家族基因的表达差异进行了分析,结果显示,12个白菜bHLH转录因子基因在不育与可育花蕾中的表达量差异显著。其中BraHLH034是拟南芥AMS基因的直系同源基因,可能参与花药绒毡层发育以及小孢子母细胞减数分裂的调控过程[11,19];而BraHLH106、BraHLH125与拟南芥的bHLH89、bHLH91为直系同源基因,与DYT1相互作用从而调控花药的发育,两个或以上的突变体会导致雄性不育现象[20]。我们分析认为BraHLH034、BraHLH106、BraHLH125等bHLH转录因子基因参与雄蕊的发育过程。本研究结果为探索植物雄蕊发育的调控机理,以及探讨bHLH转录因子参与植物雄性不育的发生机理奠定了基础。
