富营养深水水库底栖动物群落与浮游生物相关性分析*
2020-07-17池仕运韦翠珍周连凤胡菊香
池仕运,韦翠珍,胡 俊,王 瑞,周连凤,胡菊香
(1:水利部中国科学院水工程生态研究所,水利部水工程生态效应与生态修复重点实验室,武汉 430079)
(2:淮河流域水资源保护局淮河水资源保护科学研究所,蚌埠 233001)
我国水库数量众多,其在国民经济建设中发挥了举足轻重的作用. 随着社会和经济的高度发展以及人为活动的加剧,污染导致河流和湖泊供水功能弱化,水库成为了主要的替代供水水源,这也导致人们对水库生态环境的关注度逐渐提高[1]. 富营养化和藻类水华已成为当前水库生态环境的重要威胁,导致部分水库供水功能的丧失. 不同于天然湖泊,由于人类的高强度利用,水库水环境条件会发生快速波动,从而对生物群落形成选择压力,r-对策者类型的生物由于个体小,生活周期短,能够适应这种选择压力,通常在水库生物群落中占据主导地位,如浮游生物(浮游植物、原生动物、轮虫和浮游甲壳动物)和底栖动物中的小型种类如寡毛类和摇蚊类水生昆虫[2],而体型较大的底栖动物如螺贝类则在水库中多不常见.
热分层是深水水库的重要湖沼学特性,多发生于气温较高的夏秋季以及湖库的深水区. 湖库表层水体浮游植物光合作用产生的氧气是水体溶解氧的重要补充来源,但温跃层的存在阻碍了溶解氧的垂向传递. 在热分层期间,存在富营养化现象的湖库中较高浓度的氮磷水平会促进水体中浮游生物的大量繁殖,进而恶化水质,浮游生物大量死亡后沉于库底的分解活动以及沉积物的耗氧作用会进一步消耗库底溶解氧,并产生大量的硫化氢,导致库底常常处于缺氧或厌氧状态,利于厌氧微生物的活动,但对底栖动物的生长繁殖却极为不利,即使较为耐受低氧的种类也不能长期生存. 随着气温的下降,多数湖库的热分层现象会逐渐消失,水体在冬春季处于完全混合状态,底部的溶解氧水平得以提高,进而底栖动物的生存环境得到暂时改善[3-5]. 总之,存在热分层现象且处于富营养化进程中的湖库中的底栖动物会遭受周期性的缺氧扰动,其种群规模和生物多样性必然受到一定程度的影响.
在水库食物网中,浮游生物和底栖动物是鱼类的主要饵料. 浮游植物是重要的初级生产者,浮游动物和底栖动物则是重要的次级生产者. 浮游生物和底栖动物在水库分别占据着不同的生态位,对压力的响应选择不同的对策,通过食物网发生复杂的交互作用,在生态系统物质循环中发挥重要作用[6-9]. 研究表明,藻型湖泊中的底栖动物类群的主要食物来源于浮游生物类群[10-13],其比重可以占到整个湖沼次级生产力的42%[14-15],其群落结构状态对维系湖沼生态系统的正常运转至关重要. 自1980s以来,由于大力发展水库渔业的需要,作为鱼类的主要饵料来源,水库中浮游生物和底栖动物资源量的调查引起人们的重视,相关调查资料较多,但底栖动物和浮游生物的相互关系却很少引起关注,尽管这些类群在水库生态系统食物网的构建中发挥着不可替代的作用[16-18].
南湾水库是一座兴建于1950s成库60余年的老水库,具有供水、灌溉、防洪、发电、养殖、旅游等多种功能. 该水库2000-2009年10年间,除2006年和2007年处于轻度富营养化外,其余年份均处于中度富营养化水平,水温较高的月份深水区存在水温分层现象[19-20]. 汤浦水库是一座以供水为主兼防洪和改善水环境综合利用的水库,兴建于2001年,2002-2005年的监测表明水库处于中度富营养化水平[21],自4月后大部分时间存在较为明显的热分层现象. 近年来,公众对水环境质量要求越来越高,作为城市首选供水水源地的水库的富营养化和藻类水华问题也成为当政者和民众关注的焦点[21]. 本文选取以上两座存在富营养化现象以供水为主的深水水库作为调查对象,定量描述底栖动物群落状态及其与浮游生物的相互关系,探讨其相互关系与富营养化程度的耦合性,从而揭示富营养化对存在水温分层现象的水库生态系统的影响效应.
1 材料与方法
1.1 研究区域
本文选取的两座深水水库均存在富营养化和热分层现象,虽然地理位置相距较远,但均处于北亚热带区域,为典型的季风气候区,四季分明,降雨充沛,年均降雨量1000 mm以上,且均为大型丘陵性水库,水力滞留时间较长,水体交换系数较低,在生态意义上具有较多的共性.
南湾水库(31°50′~32°10′N,113°56′~114°32′E)位于河南省信阳市,属于淮河支流浉河流域,所在流域属于亚热带向暖温带过渡的季风气候区,多年降雨量平均为1226.2 mm,多集中在5-9月,占全年降雨量的70%. 南湾水库有董家河、五道河、小浉河、飞沙河和谭家河等几条汇水河流[22]. 水库兴利库容6.7×108m3,集雨区面积1100 km2,最大水面面积130.7 km2,正常水面面积77.65 km2,水库补给系数(水库集雨区面积与水库最大面积之比)为8.42. 平均水深17.4 m,最深处达38 m,水体交换系数为0.95,为信阳市的主要供水水源地[23-25].
汤浦水库(29°38′~29°54′N, 120°30′~120°45′E)位于浙江省绍兴市,属于曹娥江支流小舜江流域,水库地处亚热带季风区,多年平均降水量1564.4 mm,水库内降雨月均分配极不均匀,3-8月降雨量约占全年降水量的85%以上. 水库水系由南溪、北溪、王化溪与万宝溪四大支流组成,其中南溪、北溪在库前合流成双江溪,其入库流量占水库总径流的比例分别为双江溪75.4%、王化溪13.3%、万宝溪4.5%[26]. 水库正常蓄水库容1.8513×108m3,集雨面积460 km2,最大水面面积15.93 km2,正常水面面积14.13 km2,水库补给系数(水库集雨区面积与水库最大面积之比)为28.88. 平均水深达20 m,最深处可达30 m以上,水体交换系数为1.38. 是虞绍平原重要的饮用水源[21,26].
1.2 数据收集与处理
从水库上游到下游,依据水库结构及吞吐流特征,纵向可分为河流区(上游)-过渡区(中游)-湖泊区(下游)等不同梯级区域,在不同区域再根据水库功能将采样站位具体化. 南湾水库划分为4个分区,每个分区设1~2个采样站位,共计9个采样站位(图1). 其中站位9位于河流区,站位3、7位于河湖过渡区,站位1、2位于湖泊区,站位4、5、6、8位于库湾区. 自2015年11月-2016年9月按季节进行采样:冬季(2015年11月)、春季(2016年3月)、夏季(2016年7月)、秋季(2016年9月).
汤浦水库划分为4个分区,每个分区设1~2个采样站位,共计6个采样站位(图1). 其中站位1、2位于河流区,站位3、4位于河湖过渡区,站位6位于湖泊区,站位5位于库湾区. 自2010年7月至2011年11月分5个时段进行样品采集,分别为2010年7月、10月,2011年4月、5月和7月.
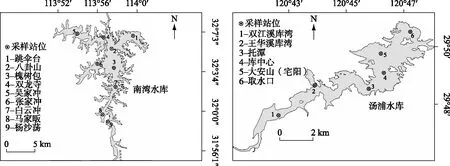
图1 南湾水库和汤浦水库采样站位示意
采用改良彼得逊采泥器(1/16 m2)进行底栖动物样品采集,每个站位采集2~4个重复样合并成一个大样. 采集的泥样经60目(250 μm)筛绢布用筛选干净后,将样品放入封口袋中带回室内进行分拣,然后置入100 mL的塑料标本瓶中,用10%的福尔马林液进行固定保存. 浮游植物、原生动物和轮虫定量样品采用5 L有机玻璃采水器采集表层、中层和下层的水样等体积均匀混合后,取2 L水样按1.5%加入鲁哥试剂现场固定,送入实验室静置沉淀浓缩至30 mL后镜检备用. 浮游甲壳动物定量样品采用25号浮游生物网过滤取自表层、中层和下层的20 L混合水获取定量标本. 采集生物样品的同时同步采集水样,带回实验室测定水体总氮(TN)、总磷(TP)、高锰酸盐指数(CODMn)和叶绿素a(Chl.a)浓度. 各指标的测定方法按照参照《水和废水监测分析方法》进行[27]. 野外采用YSI Plus多功能水质分析仪同步测定pH、溶解氧浓度,采用塞氏盘测定水体透明度.
1.3 样品鉴定
底栖动物标本送入实验室采用Olympus CX41显微镜和Leica EZ4D解剖镜进行鉴定. 鉴定时,水生昆虫除摇蚊类鉴定到属外鉴定到科,寡毛类和软体动物鉴定到属或种,其他类群鉴定到门或纲[28-29]. 底栖动物标本鉴定后进行计数称重,按采样面积分别折算成密度(ind./m2)和生物量(湿重g/m2). 浮游生物标本采用Olympus CX41显微镜进行镜检. 浮游植物计数前充分摇匀样品,取0.1 mL样品于0.1 mL计数框内进行视野法计数,原生动物和轮虫计数合用浮游植物样品,取1 mL定量样品显微镜下计数,重复取样3次取均值. 浮游甲壳动物标本在Leica EZ4D解剖镜下全部计数. 浮游植物的鉴定主要参照《中国淡水藻类——系统、分类及生态》[30],原生动物和轮虫的鉴定主要参照《淡水浮游生物研究方法》和《微型生物监测新技术》[31-32]. 浮游甲壳动物的鉴定主要参照《中国动物志——节肢动物门·甲壳纲·淡水枝角类》和《中国动物志——节肢动物门·甲壳纲·淡水桡足类》[33-34]. 浮游生物鉴定到种属水平,并计算各个类群的密度(浮游植物cells/L,浮游动物ind./L).
1.4 数据分析
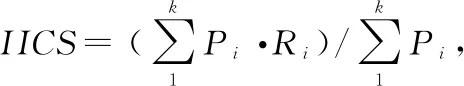
2 结果与分析
2.1 水体营养状态比较分析
调查期间,南湾水库浮游植物总平均密度为8.80×106cells/L,其中2015年11月平均密度为1.24×107cells/L,2016年3月为2.50×106cells/L,7月为7.37×106cells/L,9月为1.30×107cells/L(图2A). 汤浦水库浮游植物总平均密度达7.07×106cells/L,其中2010年7月为2.67×106cells/L,10月为6.25×106cells/L,2011年4月平均密度为2.09×107cells/L,5月为3.92×106cells/L,7月为1.65×106cells/L(图2B). 根据藻类密度来看,两座水库浮游植物总平均密度均超过106cells/L,具备藻类水华发生的条件(见全国重点湖库藻类试点监测技术规程),其中南湾水库秋冬季(2015年11月和2016年9月)和汤浦水库春季(2011年4月)浮游植物平均密度超过107cells/L. 就浮游植物门类组成来看,南湾水库蓝藻门占据绝对优势地位,为蓝藻占优势的水库,而汤浦水库硅藻门占据优势地位,为硅藻占优势的水库(图3).
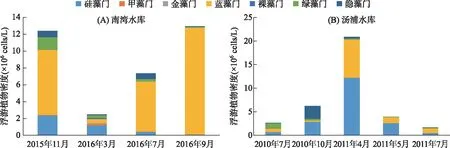
图2 两座水库浮游植物不同采样时段密度组成
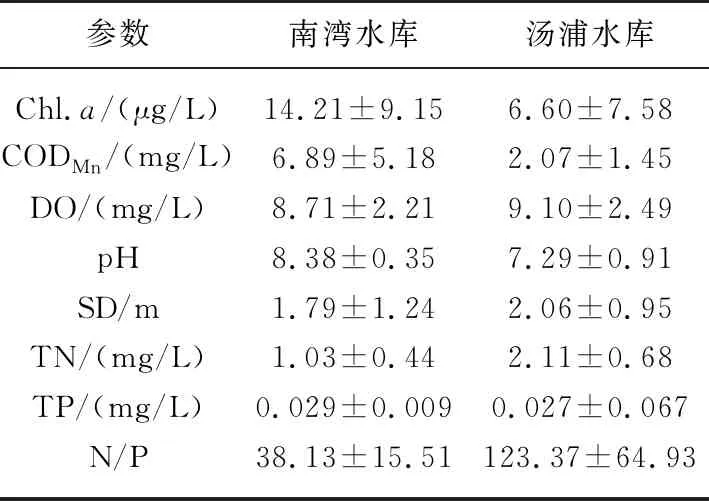
表1 调查期间两座水库的水质理化特性
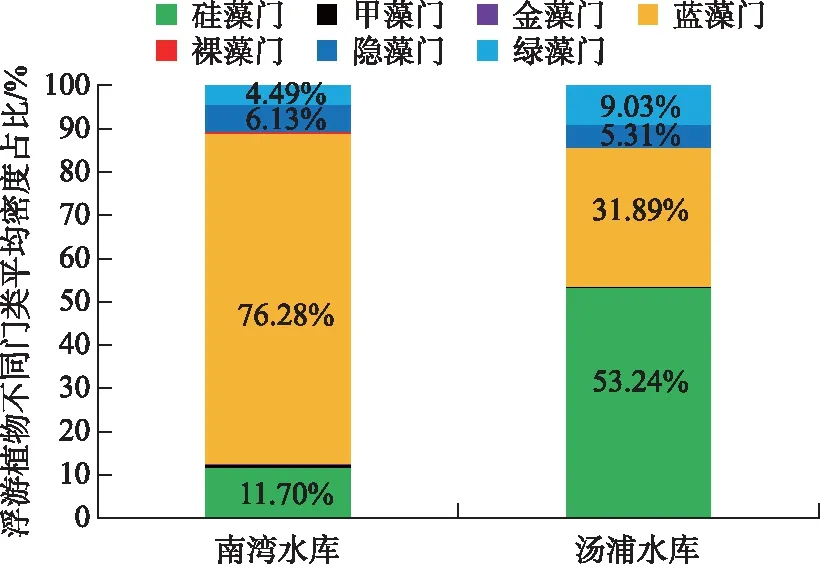
图3 调查期间两座水库浮游植物各门类组成比例
就水体理化性质而言,南湾水库的Chl.a浓度远高于汤浦水库,高锰酸盐指数是汤浦水库的3.33倍,水体由于藻类丰度较高处于弱碱性,而汤浦水库由于藻类丰度相对较低水体基本处于中性状态. 两座水库的总磷水平较为接近,但南湾水库的水体透明度和总氮水平要低于汤浦水库. 南湾水库的氮磷比为38.13,汤浦水库的氮磷比为123.37(表1).
就不同时段而言,南湾水库水温较高的7月和9月,Chl.a浓度和pH要明显高于水温较低的3月,而透明度则正好相反;汤浦水库一年的时段中,自4月后,透明度急剧下降,5-8月处于藻类急剧增长阶段,Chl.a浓度和pH明显高于其他时段. 从两座水库不同时段的理化性质结果来看,热分层期间藻类基本处于急剧增殖阶段,透明度要明显低于其他时段,pH和Chl.a浓度则明显高于其他时段,这表明两座水库的富营养化与热分层基本处于同步耦合状态(表2).
2.2 底栖动物群落结构比较
南湾水库4次调查共检出底栖动物20种,隶属6科16属,其中水生昆虫12种,寡毛类7种,线虫1种,r-对策者类型有20种,K-对策者0种. 全年来看,优势种为霍甫水丝蚓Limnodrilushoffmeisteri、裸须摇蚊Propsilocerussp.和前突摇蚊Procladiussp.;汤浦水库5次调查共采集到底栖动物27种,其中水生昆虫摇蚊类17种,软体动物2种,寡毛类7种,线虫1种,r-对策者类型有25种,K-对策者2种. 优势种为霍甫水丝蚓和前突摇蚊(表3). 就不同类群而言,南湾水库以寡毛类为主,而汤浦水库则以水生昆虫为主(图4A). 就摄食功能类群而言,南湾水库收集者占据绝对主导优势地位,相比较而言,汤浦水库收集者数量百分比远低于南湾水库(图4B). 就不同时期而言,南湾水库热分层期间(2016年7月和9月)检出的物种数分别为8和6种,远少于春季的16种和冬季的14种,汤浦水库在热分层期间(2010年7月、2011年4月、5月、7月)检出的物种数分别为8、8、9和6种,远少于2010年10月的13种.
2.3 底栖动物现存量和多样性比较分析
经Mann-Whitney检验,就底栖动物现存量而言,南湾水库密度显著高于汤浦水库,而生物量却显著低于汤浦水库;就Shannon-Wiener指数和单站出现的物种数而言,两座水库差异不显著,但Shannon-Wiener指数均值南湾水库稍低于汤浦水库,而单站物种数均值南湾水库略高于汤浦水库(表4).
就具体采样时段来看,南湾水库热分层期间(2016年7月和9月)的底栖动物密度、单站物种数和Shannon-Wiener指数的均值明显低于水温较低的2015年11月和2016年3月;汤浦水库热分层期间(2010年7月、2011年4月、5月和7月)的底栖动物单站物种数和Shannon-Wiener指数的均值均低于2010年10月(表5).
2.4 底栖动物群落结构状态与浮游生物关系
以IICS为因变量(图5A和图5B),以浮游生物各类群密度为自变量,分别针对南湾水库和汤浦水库构建PLS模型. 由于潜在因子1解释的方差最大,是很好的预测因子,因此我们考察各变量在潜在因子1中的权重(表6). 由表6可以看出:在南湾水库中,原生动物密度、轮虫密度、浮游甲壳动物密度和浮游植物密度均对IICS分值有重要影响,是很好的预测变量;在汤浦水库中,原生动物密度、轮虫密度和浮游植物密度也对IICS分值有重要影响,可以很好地预测IICS.
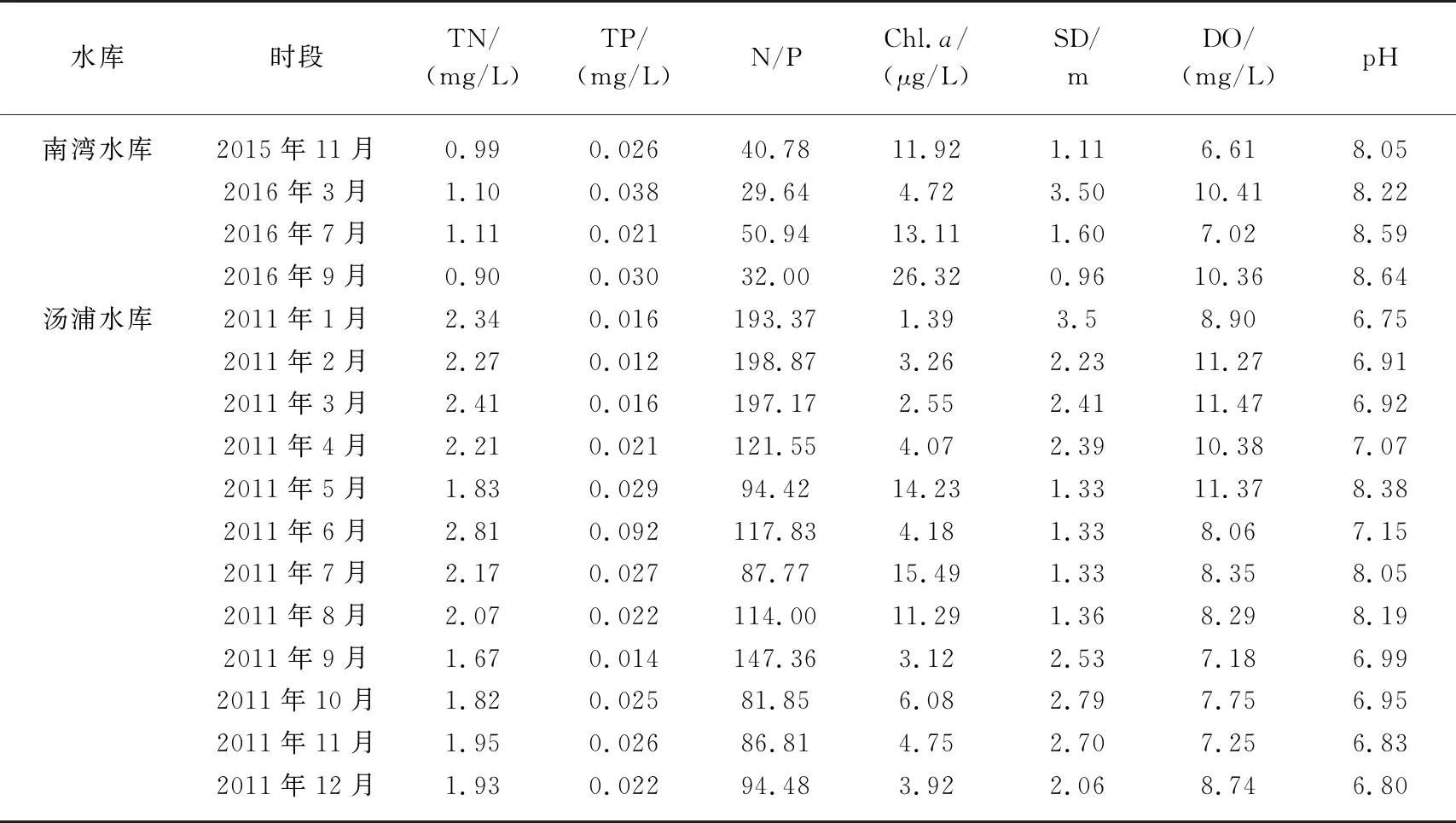
表2 两座水库不同调查时段的水质理化特性
Spearman秩相关分析结果表明:在南湾水库中,IICS分值与原生动物密度(ρ=0.504)和浮游植物密度(ρ=0.506)呈显著正相关关系(P<0.01),而在汤浦水库中则与原生动物密度呈显著负相关关系(ρ=-0.597,P<0.01). 回归图分析表明南湾水库中原生动物、浮游甲壳动物和浮游植物的密度与IICS分值具有较好的线性关系,而在汤浦水库中,原生动物和轮虫的密度也与IICS分值具有较好的线性关系(图6和图7).
3 讨论
3.1 两座水库的营养状态和底栖动物群落结构
南湾水库水质在1998年以前较为良好,随后动荡变化不定,2007年8-9月间的藻类水华的出现标志该水库局部水体接近或已经进入富营养状态[22]. 该水库日益突出的富营养化现象与库区旅游业以及库周沿岸茶叶与板栗种植业的高速发展导致大量有机物质和营养盐类进入水体,进而导致水体中总氮和总磷浓度超标频率较高有关[20,39]. 本次调查发现,南湾水库总氮、总磷浓度较高,其中总氮水平介于地表水Ⅲ~Ⅳ类之间,总磷水平超过地表水II类限值. 较高的总氮、总磷浓度会引起库区水体富营养化和藻类过度繁殖,甚至发生蓝藻水华,进而产生微囊藻毒素威胁水库水质[23,40-41]. 总氮均值1.03 mg/L,接近2007年的1.05 mg/L;总磷均值0.029 mg/L,接近2008年的0.021 mg/L[20]. 历史资料显示,南湾水库Chl.a浓度1997年为1.11 μg/L,2005年为26 μg/L[22],本次调查为14.21 μg/L,虽然较2005年有所下降,但仍远高于1997年的水平. 汤浦水库上游溪流较重的农业面源污染将大量的营养物质带入库区,造成水体氮、磷浓度过高,为库区藻类的过量繁殖提供了物质基础. 资料显示,在2002-2005年库区上游暴发过3次藻类水华,其中2003年5月的藻类水华最为严重,持续时间长达40天,全库水面被褐色水华覆盖,溶解氧小于2.0 mg/L[21];2011年4月监测到库区暴发硅藻水华[42-44]. 本次调查显示,汤浦水库总氮浓度均值2.11 mg/L,超地表水V类限值;总磷浓度均值0.027 mg/L,超地表水II类限值,处于中度富营养化水平[21]. 相比较而言,汤浦水库总氮含量要远高于南湾水库,总磷水平两座水库较为接近,氮磷比两座水库均远高于7.2,为磷抑制性水库[38],但汤浦水库要远高于南湾水库,表明汤浦水库受磷限制更为严重[26]. 就底栖动物而言,这两座水库出现的种类

表3 研究区域底栖动物名录
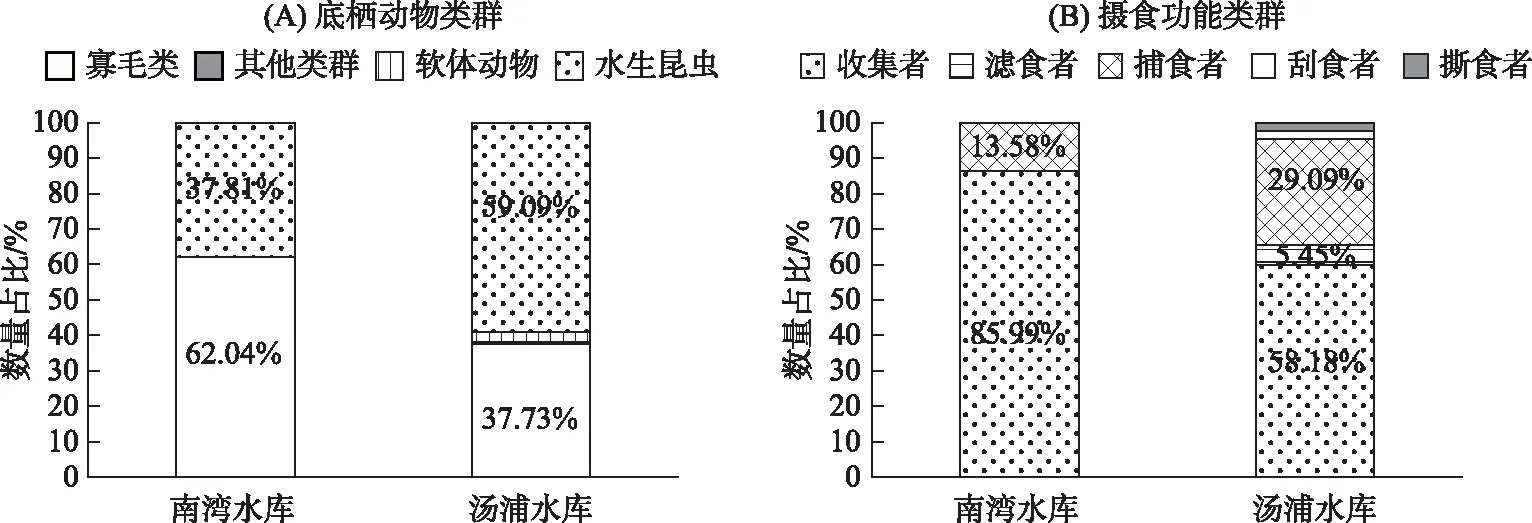
图4 两座水库的底栖动物类群和摄食功能类群组成比例

表4 两座水库底栖动物现存量、单站物种数和多样性指数比较
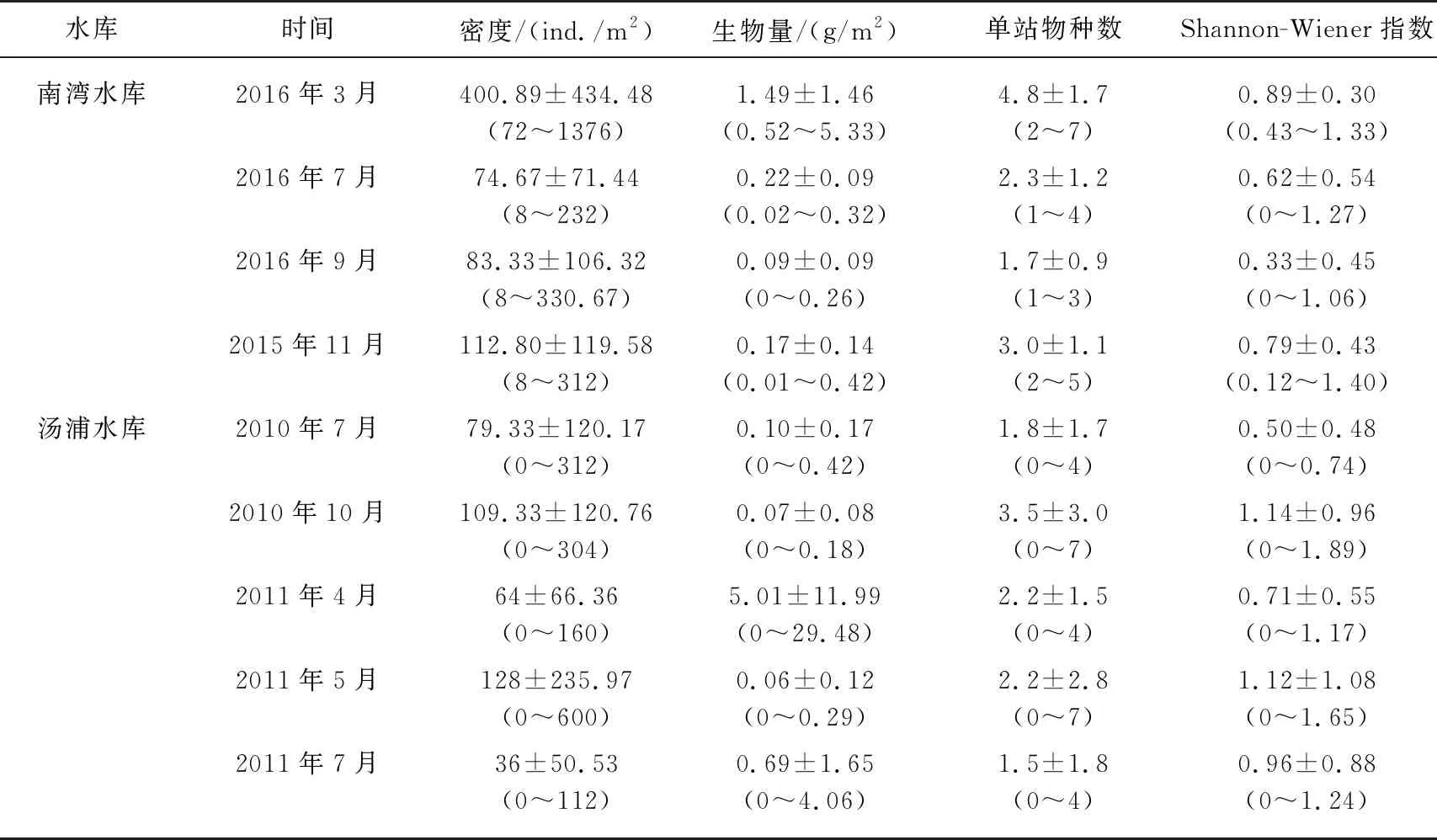
表5 两座水库在不同时段的底栖动物现存量、单站物种数和多样性指数
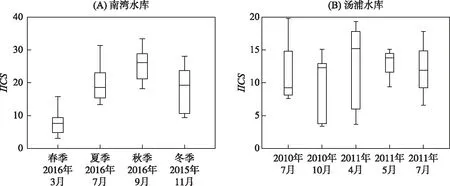
图5 南湾水库和汤浦水库的IICS分值箱线图(各水库IICS分值均为独立计算,也即单个水库不同时期的数据归集在一起进行IICS分值计算)
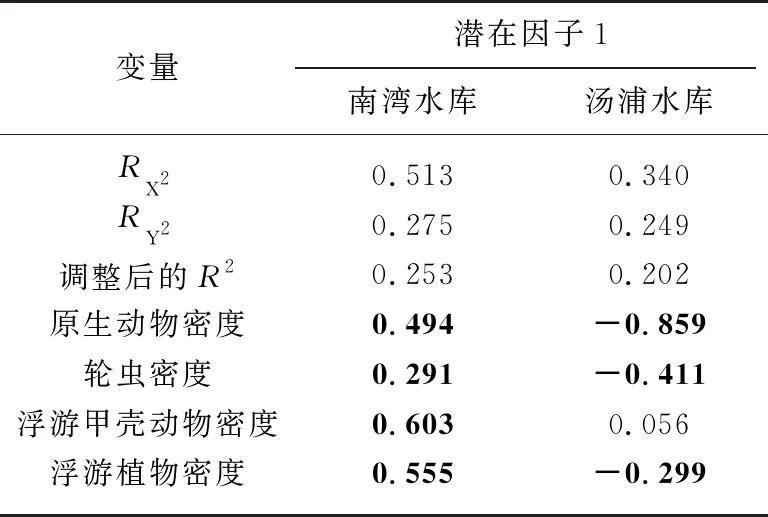
表6 PLS模型中潜在因子1解释的方差比例及各变量在潜在因子1的权重*
变量解释的方差比例, 权重绝对值大于0.2的变量粗体显示,表示该变量很重要.数均不超过30种,较为典型的富营养化指示种如霍甫水丝蚓和前突摇蚊在这两座水库中均为优势种. 研究表明,裸须摇蚊(德永摇蚊)也是典型的富营养化水体的指示生物[45],该物种广泛分布于我国南北方的湖泊和水库水体中并成为优势种[46-49]. 裸须摇蚊在南湾水库为优势种,更进一步证实该水库已经富营养化. 就藻类组成来看,南湾水库蓝藻占据绝对优势地位,更易暴发蓝藻水华,而汤浦水库硅藻占据优势,更易暴发硅藻水华. 就叶绿素a水平而言,南湾水库远高于汤浦水库,表明其富营养程度要高于汤浦水库. 就底栖动物类群而言,南湾水库寡毛类在数量上占据优势地位,而汤浦水库则以水生昆虫中的摇蚊类在数量上占优;就摄食功能群而言,南湾水库以收集者为主,其数量占比要远高于汤浦水库,这似乎表明,寡毛类和收集者更适应在富营养程度较高的水库中生存.
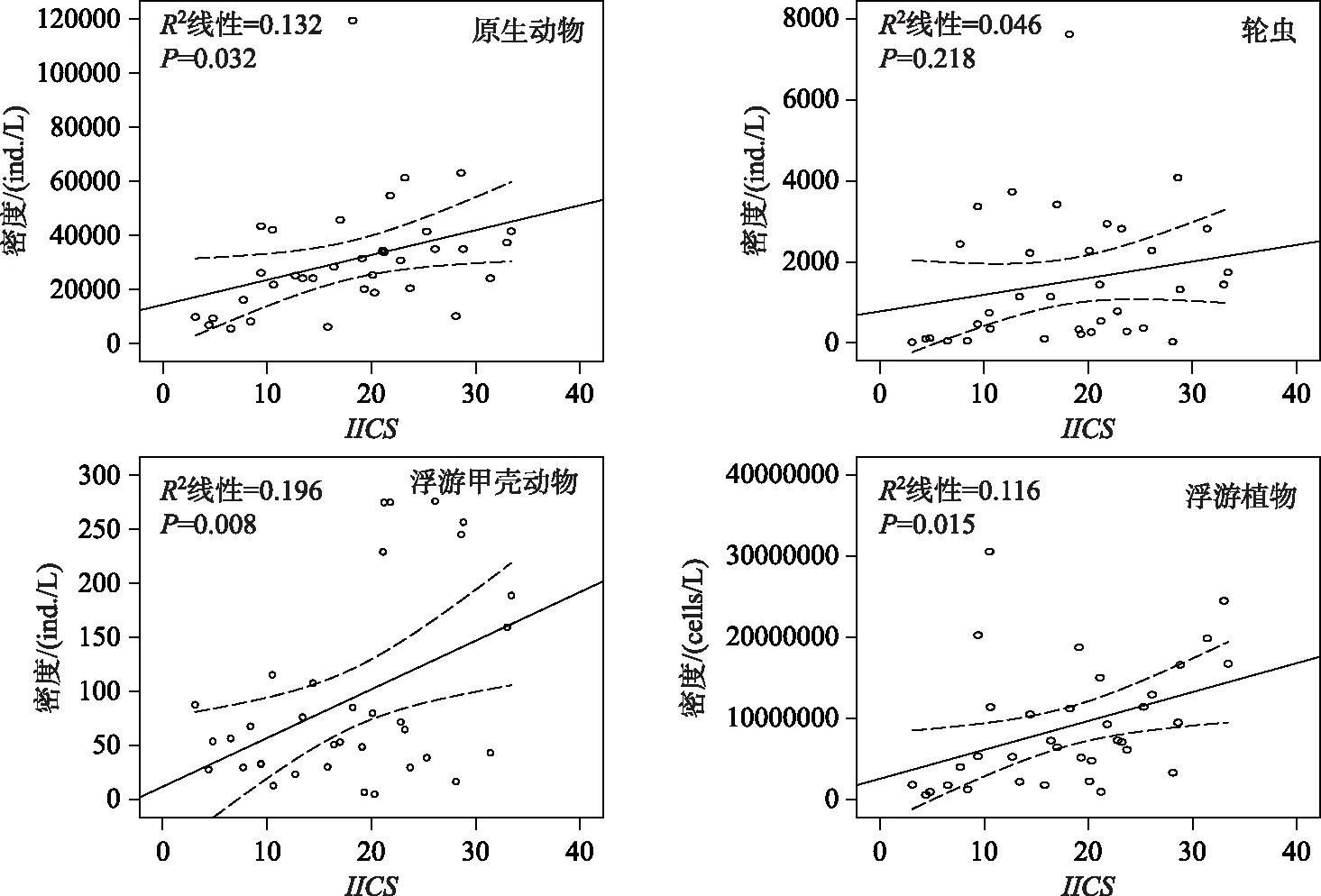
图6 南湾水库中浮游生物密度与IICS的回归图
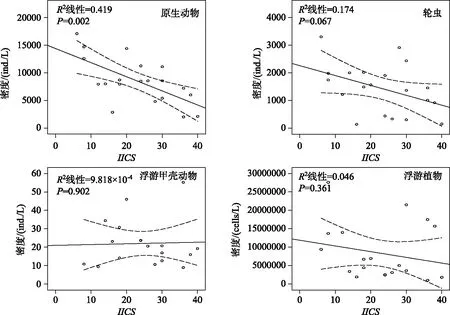
图7 汤浦水库中浮游生物密度与IICS的回归图
3.2 水温分层及富营养化对底栖动物的影响效应
大多数温带及亚热带的深水湖泊及水库都会在气温较高的季节出现水温分层现象,同为丘陵型水库的南湾水库和汤浦水库也不例外[26,50]. 水库在水温分层节律、溶解氧动态变化过程以及水力停留时间等方面均不同于天然湖泊和河流[51],其热分层和富营养化对溶解氧浓度的垂向分布有很大影响[4]. 有研究表明,汤浦水库水体表温层溶解氧基本处于饱和状态,最高可达14.8 mg/L,在不存在水温分层的情况下,溶解氧分布均匀,几乎不受水深影响,但在水体出现热分层现象时,生物呼吸、藻类尸体分解及还原性物质的氧化等活动持续消耗水中溶解氧,导致溶解氧呈现类似温度的梯度分布,库底最低时几乎为零[26]. 南湾水库也存在类似情况,由于夏季藻类过度生长,死亡的藻类残骸沉入水体下层并发生腐烂、分解等耗氧作用,水温分层现象阻碍了上下水层的溶解氧交换,进而造成水体在夏季的底部缺氧状态[20]. 研究表明,由于耐受能力差,对氧敏感的底栖动物种类会因低氧而减少或消失,而持续厌氧不仅会降低个体种类的丰度及季节与年度生长率,而且会极大地降低种类丰富度与群落生产力[52-56],与此同时,由于溶解氧限制,浮游生物会存在明显的垂向迁移行为. 南湾水库的相关调查资料表明,水温较高的夏季,浮游生物下层水体的生物量要低于中上层水体[57]. 汤浦水库的调查资料也发现浮游植物数量在进入变温层后急剧下降,至底温层后才平稳维持在一定的数量[26]. 国外的一些研究案例也表明浮游动物的垂向迁徙与溶解氧垂向分布有较大关系. 如在墨西哥北部湾,当水体底部溶解氧水平不足2 mg/L时, 浮游动物会由浑浊的深水层向表层垂直迁徙[58];在浅水湖沼系统中,当水体底部溶解氧水平趋近于零时,浮游动物由于溶解氧的限制向上层迁徙,其丰度水平远高于中下层[59]. 浮游生物的垂向迁移行为会导致水体底部食物来源的匮乏[60-61],这些均是底栖动物的分布和生长繁殖的限制因素[52-53]. 相比较而言,浅水湖泊的富营养化会促进底栖动物密度的持续增长[62-64],因这些水体中不存在水温分层现象,水体底部不存在持续时间较长的厌氧环境,浮游生物在水体中基本均匀分布,可为底栖动物提供丰富的饵料来源,从而有利于底栖动物群落的发展. 对于混合型水库而言,随着蓄水时间的延长,水库底部沉积物会越来越多,底栖动物现存量一般会呈现稳步增长的趋势[47,65]. 如丹江口水库是典型的混合型深水水库,该水库不存在水温分层现象和较为严重的富营养化现象,虽然其坝前水域深达40 m以上,底栖动物密度仍然高达30000 ind./m2以上[66]. 相比较而言,存在水温分层现象的南湾水库和汤浦水库,其底栖动物密度处于较低水平,很大可能是富营养化带来的反向效应,即富营养化程度的加重导致水体底部(温跃层以下)厌氧环境的产生和缺氧程度的加剧,浮游生物的趋避行为导致食物来源的缺乏[58-61,67-68],这些因素共同限制了底栖动物的分布和种群规模的发展. 调查期间发现,汤浦水库多个站位连续多次未采集到任何底栖动物,而南湾水库也有一个站位没有采集到任何样品,且这些站位底泥多呈现炭黑色,两座水库热分层期间底栖动物的单站物种数和Shannon-Wiener指数均值明显低于其他时段,这可能是水库热分层和富营养化对底栖动物综合影响效应的直接体现. 两个水库都有较长的水力滞留时间以及持续较长时间的水温分层现象,客观上造成库区藻类水华在一定年份出现季节性暴发现象[26,43,69]. 南湾水库在秋季由于上下层水体对流发生混合,溶解氧和营养物质相互补充,暴发蓝藻水华的概率较高,而汤浦水库则由于硅藻所占比例较高,在春季暴发硅藻水华的几率较高. 在水温分层现象期间,水库底部的缺氧或厌氧状态导致底栖动物多样性和现存量处于较低水平,分层现象结束后上下水体的对流混合将下层沉积的营养物质带入上层,客观上为藻类水华的暴发提供物质基础,藻类暴发后大量死亡的藻体沉入水底由于分解作用又加剧了水温分层期间库底的缺氧程度,从而导致底栖动物遭受周期性的缺氧扰动,其多样性和现存量难以得以提高.
3.3 底栖动物和浮游生物的关系
研究表明,在深水湖泊中浮游植物碎屑的沉积为底栖动物提供了可观的能量来源[10]. 肠道物检测结果表明,糠虾Mysisrelicta体内含有浮游植物以及浮游动物和轮虫残渣[70]. 就一年尺度的观察结果而言,瑞典维内尔湖片脚类动物Monoporeiaaffinis的密度和春季硅藻的生物体有着非常重要的关系[13,71]. 在湖泊和水库生态系统中,除了几种肉食性种类外,深底带的绝大部分大型底栖动物种类与个体是杂食性的,主要以细菌包裹的碎屑颗粒及沉降到沉积物表面的藻类为食[56]. 根据PLS模型的分析结果来看,原生动物密度、轮虫密度和浮游植物密度对南湾水库和汤浦水库的底栖动物群落状态均有重要影响,是很好的预测变量,但预测的方向正好相反. 在南湾水库中,原生动物、轮虫和浮游植物的密度上升伴随底栖动物群落状态的下降,也即这些浮游生物类群密度的升高不利于底栖动物群落的发展,其可能的原因是富营养化导致浮游生物的过度生长,水温分层现象的形成也伴随着溶解氧分层的形成以及库底缺氧或厌氧状态的形成,虽然浮游生物丰度的升高客观上为底栖动物提供了可观的食物来源,但由于底部生存环境的恶化,底栖动物已不能有效加以利用. 在汤浦水库中,原生动物、轮虫和浮游植物的密度上升有助于底栖动物群落状态的提高,也即这些浮游生物类群密度的增长有利于底栖动物群落的生长繁殖,这符合在湖沼生态系统中浮游生物为底栖动物提供饵料来源的预期[72-73],同时也表明汤浦水库富营养化促进了浮游生物丰度发展,虽然也存在水温和溶解氧分层现象,但浮游生物沉积的残渣能够为底栖动物充分利用,其生存环境似乎要好于南湾水库. 根据以上结果我们推断:在水库还未进入以蓝藻占优势类群为表征的富营养化状态时,浮游生物类群可为底栖动物提供较为可观的食物来源,其与底栖动物群落在食物链上存在较为紧密的正相关关系,但当水库已经进入以蓝藻占优势类群的富营养化状态时,水体的高营养状态虽然促进了浮游生物类群的大发展,但由于缺氧导致的生存环境恶化,浮游生物与底栖动物在食物链上的联系不再紧密,也即底栖动物已不能有效地利用浮游生物提供的饵料来源,反而由于大量浮游生物死亡残体的堆积造成的低氧或厌氧面积的扩大导致种群发展受限. Spearman秩相关分析和回归图分析结果也表明,富营养化程度较轻的汤浦水库中,浮游生物类群中的原生动物、轮虫和浮游植物密度与底栖动物群落之间存在较强的正相关关系,而富营养化程度较重的南湾水库中,浮游生物所有类群与底栖动物群落之间存在较强的负相关关系,这与PLS模型的分析结果可以相互印证.
根据以上分析可以看出,营养化程度的加重会导致存在水温分层现象的水库中与沉积物接触的恒温层存在严重的缺氧乃至厌氧现象,同时浮游生物特别是浮游动物存在垂向迁移现象,可以规避不利生境从而向溶解氧和光照丰富的上层水体迁移[61,67,73-74],导致水体底部浮游生物种群的匮乏,而上层水体密度过大的藻类死亡后沉入水体底部在矿化分解过程中加速了溶解氧的消耗进而加重了水底的缺氧程度[75-76],最终饵料来源的缺乏以及厌氧环境的形成导致底栖动物种群的衰退. 总之,水库水温分层现象的存在对底栖动物群落的发展是一大制约因素,富营养化程度的加重可导致底栖动物种群的持续衰退,底栖动物和浮游生物的关系也会由相互促进的正相关关系转变为彼此制约的负相关关系. 而这种转变又与富营养化程度的加剧导致的水底栖息环境的恶化和浮游生物对不利环境的趋避行为导致的食物匮乏有关. 当然这些推断需要更多实证数据的支撑,今后在富营养化水温分层水库中系统开展浮游生物的垂向分布与底栖动物群落及其分布关系的研究很有必要. 如果这一推论得到更多野外数据的支撑,则今后可通过探讨浮游生物和底栖动物群落的关系来推证水库的富营养化程度,从而为水库管理方提供更多有价值的参考信息,进而采取必要措施保护和改善水库生态环境.
致谢: 感谢水利部中国科学院水工程生态研究所的李嗣新、张俊芳、陈威同志对野外工作的帮助. 感谢河南省南湾水库管理局和浙江省绍兴市汤浦水库有限公司的同志予以的大力支持与配合.
