葛洲坝下中华鲟(Acipenser sinensis)性腺退化严重吗?*
2020-07-17黄真理王鲁海
黄真理,王鲁海
(中国水利水电科学研究院,北京 100038)
1981年1月4日,葛洲坝工程截流,中华鲟(AcipensersinensisGray, 1835)被阻隔在葛洲坝下形成新的产卵场(图1). 之后,中华鲟被列为葛洲坝救鱼的唯一对象. 关于葛洲坝对中华鲟到底带来什么样的影响?有共识,也有分歧. 无法回避的事实是:中华鲟被阻隔,洄游距离缩短1175 km;中华鲟的产卵场地点改变,空间范围上从葛洲坝截流前的宜宾到万州800 km江段缩小到葛洲坝截流后坝下30 km江段,产卵场数量也从19处缩减为1~2处. 但是,对于中华鲟的发育、产卵繁殖规模、性腺退化是否受到葛洲坝乃至长江水坝的影响,专家们的调查结论莫衷一是. 其中,性腺退化与否是核心和关键.
1982-1985年,中国水产科学研究院长江水产研究所的柯福恩等调查发现,葛洲坝导致中华鲟的性腺退化十分严重,产卵规模相当有限,若干年后资源量将进一步衰竭[1]. 然而,中国科学院水生生物研究所(以下简称中科院水生所)周春生等[2]和曹文宣等[3]认为:中华鲟在葛洲坝下性腺能够发育成熟,个别性腺发育不正常被认为是病态或鱼体受伤所致. 1990年,陈金生等的调查研究[4],否定了柯福恩等关于葛洲坝导致中华鲟性腺退化的结论. 此后,尽管也有研究报告指出性腺退化问题[5],未再引起重视和深入研究. 葛洲坝修建后,乐观派认为葛洲坝对中华鲟产卵繁殖影响不大,这个观点一直占据主导地位,导致中华鲟性腺退化问题一直被忽视. 我们认为,这是关系到葛洲坝影响和中华鲟能否维持种群数量的重大问题,有必要重新认识和评估.
1 材料和方法
1.1 中华鲟在长江的时空分布和性腺发育
要分析前人研究为什么出现不同的结论,我们需要了解中华鲟在长江的时空分布和性腺发育. 如图1所示,中华鲟每年6-8月进入长江口,开始溯河洄游,第2年秋季10-11月产卵繁殖后,迅速降河洄游海洋. 幼鱼出生后在第3年的夏季5-7月进入河口地区. 因此,在长江存在两个繁殖股群(cohort),如图2所示,我们把当年进入长江的繁殖群体,叫做新股群;把上一年进入长江的繁殖群体叫做老股群(图2)[6]. 我们提出洄游动力学模型(migration dynamics model, MDM)应用于长江中的中华鲟繁殖群体[7]和幼鱼的洄游过程[8],获得了中华鲟时空分布和生活史(图3). 通过对比葛洲坝截流前后中华鲟的洄游过程的时空分布,我们揭示中华鲟新、老股群的时空分布规律如下[7]:

图1 长江中华鲟洄游路径和产卵场
1)中华鲟新股群在每年6-8月进入长江口,上溯速度先快后慢,8月中下旬就进入了湖北江段,9月初-9月下旬,中华鲟新股群在湖北江段密度分布最高,这就是历史上(葛洲坝修建前)为什么在湖北江段9月份是捕捞中华鲟新股群的最佳时机(金沙江江段为10月). 中华鲟上溯到达湖北后,“大部队”(密度峰值)在汉口-宜昌江段停留的时间大约4个月(9月初-12月底).
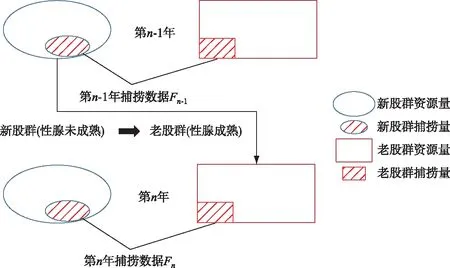
图2 长江中华鲟繁殖群体的种群结构

图3 中华鲟的生活史(a)和性腺发育阶段(b)[7-8]
2)葛洲坝截流前,每年9-11月,中华鲟在长江的分布主要集中在湖北江段(新股群)和四川江段(老股群),其中,新股群主要集中在城陵矶-宜昌江段,老股群主要集中在宜宾江段. 新股群和老股群相对独立分布,没有混杂在一起. 因此,在宜宾江段捕捞的通常是老股群,在湖北江段捕捞的是新股群. 葛洲坝截流后,中华鲟被阻隔在坝下,每年9-11月中华鲟新、老股群交织在一起,因此,繁殖季节的捕捞包括新、老两个股群.
3)1981年1月4日,长江中只有1980年进入的老股群. 葛洲坝截流,正好发生在1980年股群上溯通过葛洲坝的时候. 通过利用历史种群模型和MDM模型计算得到一致的结果[6-7]:葛洲坝截流,将中华鲟繁殖群体一分为二,上游660尾(占1980年群体1009尾的65%),继续向上游洄游并在金沙江繁殖;其余349尾被阻隔在葛洲坝下,参与1981年秋季葛洲坝下的繁殖. 1981年秋季也成为历史上金沙江和葛洲坝下同时存在繁殖行为的唯一年份.
中华鲟是产卵洄游性鱼类,洄游长江的目的是为了繁殖后代,在洄游过程中禁食. MDM模型和种群模型,有助于了解野外捕捞调查所对应的种群结构特征. 过去,由于缺乏中华鲟的时空分布和种群数量,加上采集地点的差异和捕捞样本的限制,利用野外捕捞数据评估中华鲟性腺发育状况,不区分新、老股群,结果差异较大,具有一定的误判和局限性.
我们提出中华鲟性腺成熟参数(maturity parameter of gonads)H的概念[7],参照性腺分化后的卵巢(精巢)发育7个时期(0~Ⅵ)来划分中华鲟的性腺,对应性腺成熟参数H数值为0~6. 中华鲟繁殖群体在长江中的性腺发育为Ⅲ~Ⅵ期,因此,我们重点研究H=3~5的性腺发育过程及其影响因素. 性腺成熟参数H是对中华鲟性腺发育的分级计量,与传统的性腺成熟指数(gonadosomatic index,GSI)不同,GSI是指卵巢或精巢重量与鱼重的比值. 中华鲟亲鱼性腺发育依靠吸收性腺脂肪和沿途溯河过程中水流的刺激,经过1年时间从Ⅲ期(H=3)发育到Ⅳ期(H=4),又经过3个月时间从Ⅳ期(H=4)发育到Ⅳ2期(H=4.2,性腺成熟基线值). 因此,性腺成熟参数H可以表达为洄游距离和时间的函数,我们可以获得中华鲟的性腺发育模型[7]. 图4a为中华鲟上溯过程中的性腺发育图. 由于葛洲坝的阻隔影响,导致中华鲟的性腺发育达到成熟的时间推迟37天(图4b).

图4 中华鲟繁殖群体在长江的性腺发育过程和葛洲坝的影响[7]:(a)中华鲟性腺发育过程;(b)性腺成熟比例
根据中华鲟上溯过程的性腺发育模型,计算结果表明[7]:亲鱼的性腺成熟参数由H=3(Ⅲ期)发育到H=4(Ⅳ期),性腺成熟参数H在长江的增量部分,80%是靠吸收性腺脂肪发育完成的,20%依靠流水刺激. 这是从种群角度来说的,针对个体亲鱼,由于其脂肪储存的差异,相对比例可能是个变量. 另一方面,流水刺激等环境条件对性腺发育发挥着重要作用,其作用机制尚不清楚,葛洲坝的影响不可忽略.
1.2 中华鲟性腺退化的调查结果为何完全不同?
目前,我们能够收集到的关于葛洲坝下性腺发育调查的早期文献共有5篇,如表1所示,其中长江水产研究所2篇,中科院水生所3篇①陈金生, 余志堂, 刘家寿(1990)的报告《葛洲坝下游中华鲟性腺发育状况调查》,收录在文集中没有单位的署名. 据了解,3位作者1987年以前的工作单位是中国科学院水生生物研究所,以后调到水利部中国科学院水库渔业研究所(1987年成立).. 这5篇文献中,曹文宣等[3]是一篇关于葛洲坝救鱼的综述性文章,没有性腺发育正常与否的相关数据和分析,肯定了“被滞留于葛洲坝下江段的中华鲟性腺能正常发育成熟”,说明葛洲坝对中华鲟的性腺发育没有显著影响;认为少数个体性腺呈现退化现象, 可能与鱼体受伤有关. 危起伟等[5]是一篇关于中国鲟鱼保护的综述性文章,给出了葛洲坝修建前中华鲟Ⅲ期和Ⅳ性腺的特征值,包括性腺脂肪、成熟系数和卵径,作为分析葛洲坝修建后性腺发育正常与否的参照. 该文给出了1984年10月的10尾雌鱼的特征参数,只有1尾达到成熟,其余9尾性腺脂肪耗尽但成熟系数和卵径均不符合IV期性腺的参照值,说明葛洲坝修建后性腺退化严重. 陈金生等[4]是一篇内部简要报告(3页正文),受水电部驻葛洲坝代表处的委托进行的调查,该文的目的针对“水产工作者提出中华鲟性腺退化、成熟个体比例变化,预言中华鲟资源将衰竭”进行复核. 该文的“材料和方法”部分语焉不详,比如,调查样本76尾,来源模糊. 从附件关于亲鱼捕捞记录的时间(10月23日-11月6日,15天)来看,他们调查资料主要来自中华鲟研究所和长江水产研究所采集的样本. 他们只是采用挖卵器采集性腺样本,并没有性腺的切片分析和性腺退化指标,更没有测量性腺脂肪、性腺重量、卵径等柯福恩等已测的指标,实际上无法判断性腺发育是否出现退化. 因此,这样的复核报告是不可靠的,只会误导管理部门.

表1 中华鲟性腺发育调查信息
因此,我们重点分析柯福恩等[1]和周春生等[2]的成果. 从文章发表的时间看,最早是周春生等(1984年4月20日收稿,1985年6月发表),柯福恩等发表于1985年7-8月. 周春生等的论文,重点是描述中华鲟的性腺发育各个阶段的分期特点,涉及性腺退化的内容为:“有些个体的性腺发育不正常”、“这些不正常现象是属于病态抑或退化, 尚有待进一步研究”. 但没有给出收集或切片的样本中有多少属于“性腺发育不正常”,忽视或回避了葛洲坝对性腺发育的影响和性腺退化的严重性. 该文的结论说:“被葛洲坝阻隔而不能到达长江上游产卵的中华鲟在坝下江段性腺能够继续发育, 达到成熟, 并在适当的环境中进行自然繁殖”. 这句话的意思是,葛洲坝下中华鲟的性腺基本上都能够发育成熟和自然繁殖,变相否定了葛洲坝对性腺发育的影响. 我们认为,葛洲坝下捕捞的样本中包含性腺成熟亲鱼,以及在葛洲坝下捕捞到幼苗或食卵鱼,只能说明葛洲坝下发生了产卵行为,也可以证实有新的产卵场,但不能外推葛洲坝下的亲鱼性腺都能发育成熟,更不能说明葛洲坝对中华鲟的性腺退化没有影响. 捕捞调查属于环境和目标 “非受控”条件下的“不严格”试验方法,外推结论要十分小心和慎重. 如果是“推测”,需要接受其他相关实验的验证.
柯福恩等(1985)[1]最早指出葛洲坝导致中华鲟发生性腺退化,他们测量了性腺脂肪、性腺重量、卵径和成熟系数等相关参数,给出了这些参数对应的Ⅲ期和Ⅳ期性腺的范围. 由于中华鲟亲鱼在长江中禁食,主要依靠性腺脂肪完成性腺发育. 传统上,长江渔民根据性腺中脂肪含量的多少把中华鲟分为“油子(含脂肪多,也称为脂鲟)”和“水子(含脂肪少或没有脂肪)”,说明脂肪消耗与性腺发育成熟度有关. 因此,他们通过性腺脂肪、性腺重量、卵径以及性腺发育状况来评估性腺发育和退化的方法是合理和有效的,其结论表明:(1)1984年4-5月和10-11月共获标本62尾,其中雌鱼28尾,71.43%性腺退化;雄鱼34尾,29.41%性腺退化. (2)1984年10月19日-11月2日获得的12尾雄鱼和10尾雌鱼中有6尾雄鱼和1尾雌鱼性腺发育成熟. 而1985年11月19日-22日获得的9尾雄鱼和6尾雌鱼全部性腺退化,说明11月中旬是中华鲟种群性腺发育是否全部退化的重要时间节点. 因此,柯福恩等(1985)[1]关于中华鲟性腺退化十分严重,产卵规模相当有限,若干年后资源要下降的结论是可靠的,以后也得到了大量证据的证明. 反观中科院水生所有关否定中华鲟性腺退化的方法和材料,是不充分和不可靠的.
我们认为,葛洲坝对中华鲟性腺发育的影响,在葛洲坝救鱼论证和实践中被长期忽视或轻视,给中华鲟保护带来严峻的后果. 因此,有必要提供更多的证据.
1.3 中华鲟性腺退化的其他证据
1)根据1981年中科院水生所和四川省水产所两家共同的解剖数据,我们利用葛洲坝下种群结构和数量的新成果[6-7]来推算中华鲟的性腺成熟率. 应该指出,在秋季捕捞的亲鱼中包含新、老股群,葛洲坝对新老股群的性腺发育的影响程度差异显著,当年的新股群就是第二年的老股群,两者存在转化关系. Wei等(2007)[5]把新、老股群混在一起进行分析的做法,会低估性腺退化率. 因此,本文性腺成熟率是指老股群中能够发育成熟的比例,产卵环境适宜时也指中华鲟的有效繁殖系数. 反之,老股群中性腺退化鱼所占的比例就是性腺退化率. 1981年的这些解剖数据收录在曹文宣等(1989)(第218页)[9]和胡兴祥(第7页)[10]的报告中:1981年10-11月,在宜昌和石首共解剖了139尾中华鲟,其中雄鱼62尾,有2尾鱼的精巢已经发育到IV期;雌鱼77尾,有6尾鱼的卵巢已经发育到IV期末,另外还有3尾产过卵. 因此,可以确认,139尾中华鲟有11尾是正常发育、性腺成熟的个体. 由前述1.1节的分析可以知道,1981年10-11月捕捞的139尾中包含新、老两个股群,我们只需要估算出老股群数量,就可以计算性腺成熟率或退化率.
1981年在葛洲坝下的捕捞量为1002尾. 坝下的资源量为1358尾,其中349尾1980年股群是葛洲坝截流拦在坝下的,1009尾是1981年股群(补充群体)[6-7]. 因此,假设捕捞具有随机性,这样,1002尾中的老股群为258尾(=1002×349/1358),新股群为744尾. 因此,可以计算出139尾解剖样本中老股群数量为36尾(=139×258/1002). 所以,1981年解剖的139尾中华鲟性腺成熟率为:30%(11/36). 换句话说,中华鲟的性腺退化率为70%. 这个估值,与我们给出的性腺退化率理论值75.8%比较接近[7].
2)危起伟[11]利用标记-重捕法估算了葛洲坝下中华鲟产卵场的繁殖群体数量,1996-1998年分别为426尾(雌鱼309尾,雄鱼117尾)、263尾(雌鱼191尾,雄鱼72尾)、320尾(雌鱼232尾,雄鱼88尾),3年平均336尾(雌鱼244尾,雄鱼92尾). 同时,根据采卵试验估算1996-1998年实际产卵雌鱼数量分别为94尾、18尾、154尾,平均89尾,占雌鱼总数244尾的36.5%,说明约有63.5%的雌鱼发生了性腺退化,没有参加繁殖活动. 为了估算中华鲟雄鱼的性腺退化率,根据1996-1998年雌雄性比2.65,雄鱼比雌鱼对环境变化更为敏感(这也为其他鱼类所证实),说明雄鱼比雌鱼的性腺退化更为严重,实际参加繁殖的雄鱼数量占雄鱼总数的比例为36.5%/2.65=13.77%. 因此,综合考虑1996-1998年实际参加繁殖的中华鲟雌鱼和雄鱼数量占中华鲟总量的比例(性腺成熟率)为 (36.5%+13.77%)/2=25%,退化率为75%.
3)长江水产研究所危起伟团队收集了1981-1995年捕捞的563尾中华鲟(其中雌鱼255尾,雄鱼308尾),对性腺发育分期和性腺发育状况进行了不完整的统计,其中性腺发育状况分为“normal”、“good”、“degeneration”、“不清”,我们把性腺分期和性腺发育信息完整的385尾中华鲟按照新、老股群进行分类和分析,发现新股群和老股群都出现性腺退化. 由于每年的样本数量很少(平均29尾),根据每年样本估算老股群的性腺退化率误差较大. 假设n年的样本新股群性腺退化率为An,样本老股群性腺退化率为Bn,因此,考虑新老股群的转化关系,估算n年老股群性腺退化率为Kn=1-(1-An-1)(1-Bn). 重新整理的老股群性腺退化率估算值和理论值如图5所示. 由于每年的样本量较少,估算值显得比较分散. 但性腺退化率的理论值在估算值范围内.

图5 中华鲟性腺退化率的估算值与理论值[7]
4)葛洲坝截流前,中华鲟雌鱼的怀卵量较大(平均64万粒/尾),雄鱼精子活力较高,寿命较长. Xiao等[12]报道了中华鲟研究所为了繁殖捕捞的中华鲟野生亲鱼的精子活力(图6a). 可以看出,葛洲坝截流后,反映中华鲟精子活力的寿命和快速活动期是逐步减少的,1990s下降显著. 刘鉴毅等[13]发现中华鲟繁殖力显著下降,与葛洲坝截流前相比,1998-2004年期间,中华鲟雌鱼的平均怀卵量只有葛洲坝截流前1976年的一半,雄鱼相对怀精量也有所下降. 郑跃平[14]进一步发现1973-1976、1982-1984、1998-2004、2005-2006年4个时期中华鲟精子活力逐步衰退的事实(图6b). 精子活力显著下降,反映出中华鲟精子质量的持续下降,必然导致受精率大幅降低和后代数量的减少. 中华鲟繁殖能力的下降,对于中华鲟自然种群的恢复非常不利,直接说明中华鲟性腺退化的严重性.

图6 葛洲坝修建后中华鲟精子活力逐步衰退[12,14]
2 结论
本文针对葛洲坝下中华鲟性腺退化这一存在争议、长期忽视、对野生群体存亡关系重大的问题,利用我们提出的创新理论——MDM模型、性腺发育模型和种群模型,通过对相关文献和观点的重新分析和评价,给出中华鲟性腺发育严重退化的若干证据. 我们的研究表明:(1)长江水产研究所柯福恩等(1985)[1]对葛洲坝下中华鲟性腺退化的工作是扎实的,提出的性腺退化的测量指标和评价方法是恰当的,他们关于中华鲟性腺退化十分严重,产卵规模相当有限,若干年后资源要下降的结论是可靠的,也得到了1990s以来中华鲟持续衰退事实的证明. 反观中科院水生所有关否定中华鲟性腺退化的方法和材料,是不充分和不可靠的. (2)葛洲坝对中华鲟的性腺发育和繁殖产生了严重影响,阻隔效应导致中华鲟的性腺退化率为75%左右. (3)中华鲟属于生命周期长和生殖年龄高的鱼类,建坝对性腺发育和繁殖活动的影响,将在建坝10年以后导致种群数量的减少. 在葛洲坝救鱼过程中,除了中科院水生所外,来自不同研究机构和专家1985年以后多次指出了葛洲坝对中华鲟性腺发育影响严重,为什么没有深入研究相关对策?中科院水生所及其相关专家,是葛洲坝救鱼中被决策认可的权威机构和专家,为什么轻率否定中华鲟性腺退化?性腺退化被长期忽视或轻视,对中华鲟保护产生了严重影响.
致谢:感谢长江水产研究危起伟团队提供的1981-1995年的捕捞数据资料. 特别致谢长江水产研究所的老所长柯福恩先生及其团队,他们是中华鲟性腺退化的“吹哨人”.
潜在利益冲突声明:本文作者不存在相关利益冲突,包括但不限于从利益相关方获得可能会影响本文公正性和客观性的经费资助;本文作者之一黄真理,1993-2012年在原国务院三峡工程建设委员会办公室工作,参与组织协调三峡工程相关的生态与环境保护工作.
