我们能拯救长江中正在消逝的鲟鱼吗?*
2020-07-17谢平
谢 平
(1:中国科学院水生生物研究所东湖湖泊生态系统试验站,武汉 430072)
(2:云南大学生态与环境学院高原湖泊生态与治理研究院,昆明 650500)
很多物理现象往往具有惊人的可预测性,如遥远星辰的摇晃和转动,这是一种美好而严格的科学. 但我们对生态学趋势的预测就没那么精准了,把握成千上万的物种交织在一起的现在就已经十分吃力了. 但种间关系的复杂与变异并不意味着物种不存在稳定的生理节律. 事实上,有些物种对生存环境精准性的生理追求十分悲催,不惜自我毁灭,不知是一种笨拙还是一种忠贞?长江中的鲟鱼就是这样一个悲壮的类群. 复杂性造就了精致性,而精致性却牺牲了可塑性.
鲟形目(Acipenseriformes)是软骨鱼类与硬骨鱼类之间的过渡类群,是鱼类中最濒危的类群,63%的种类处于极危(critically endangered)状态,另外有3个种可能已经绝灭,其余的30%也受到威胁或处于易危或濒危状态[1]. 鲟鱼种群数量的下降源自过度捕捞、河流建坝和水污染等[2].
1 古老的大型鱼类——鲟鱼
鲟形目包括鲟科(Acipenseridae,通俗英文名Sturgeon)、匙吻鲟科(Polyodontidae,通俗英文名paddlefishes)和一些灭绝的科. 鲟鱼是一种古老的鱼类——我国曾在距今2.5亿多年前的二叠纪地层(甘肃省肃北县)中发现过鲟形目鱼类化石,称之为中华原始软骨硬鳞鱼(图1),它兼有鲟形目鱼类、比耶鱼类和古鳕类的一些特征,可能是繁盛于古生代的古鳕类与自中生代开始繁荣的鲟形目鱼类之间的一个过渡类群[3].
在我国中生代地层发现过不少鲟形目鱼类化石,如北票鲟、燕鲟、原白鲟、辽鲟等[4]. 譬如,在我国的一些晚侏罗世地层中,发现有辽宁北票的北票鲟(Peipiaosteus)(图2)[5],河北丰宁的丰宁北票鲟(P.fenningensis)[6],辽宁凌源的刘氏原白鲟(Protopsephurusliui)[3].
鲟形目鱼类之所以被认为是原始鱼类,是因为它们的形态特征与其最早的化石记录相比并无太大改变,因此也有“活化石”之称[7-8]. 全世界的鲟科(Acipenseridae)鱼类共有27种,分为4个属——鲟属(Acipenser)、鳇属(Huso)、铲鲟属(Scaphirhynchus)和拟铲鲟属(Pseudoscaphirhynchus). 全世界的匙吻鲟科鱼类更少,仅有两属两种:一种是分布于长江的白鲟(Psephurusgladius),另一种是分布于密西西比河的匙吻鲟(Polyodonspathula). 早在20世纪初,Sowerby[9]就曾这样描述道:“扬子江白鲟是所有中国令人印象深刻的鱼类中的首位了. 扬子江白鲟属于匙吻鲟科鱼类,在世界上仅存2属2种,一种是中国的白鲟,另一种即是分布于北美密西西比河的匙吻鲟,两者皆属于鲟鱼类”.
根据线粒体基因组进行分子钟估算的结果,鲟形目约起源于3.62亿~4.14亿年前,鲟和匙吻鲟大约是在1.32亿~1.60亿年前分开的[10]. 通过线粒体细胞色素b基因分子钟的估算,鲟和匙吻鲟的分歧约发生在1.84亿年前,太平洋支和大西洋支的分歧时间约在1.21亿年前,黑龙江-松花江-乌苏里江流域的史氏鲟与长江流域的中华鲟和达氏鲟的分歧约发生于0.70亿年前,而中华鲟和达氏鲟的分歧时间约在1050万年前,密西西比河的匙吻鲟和长江的白鲟的分歧时间约在0.68亿年前(图3).
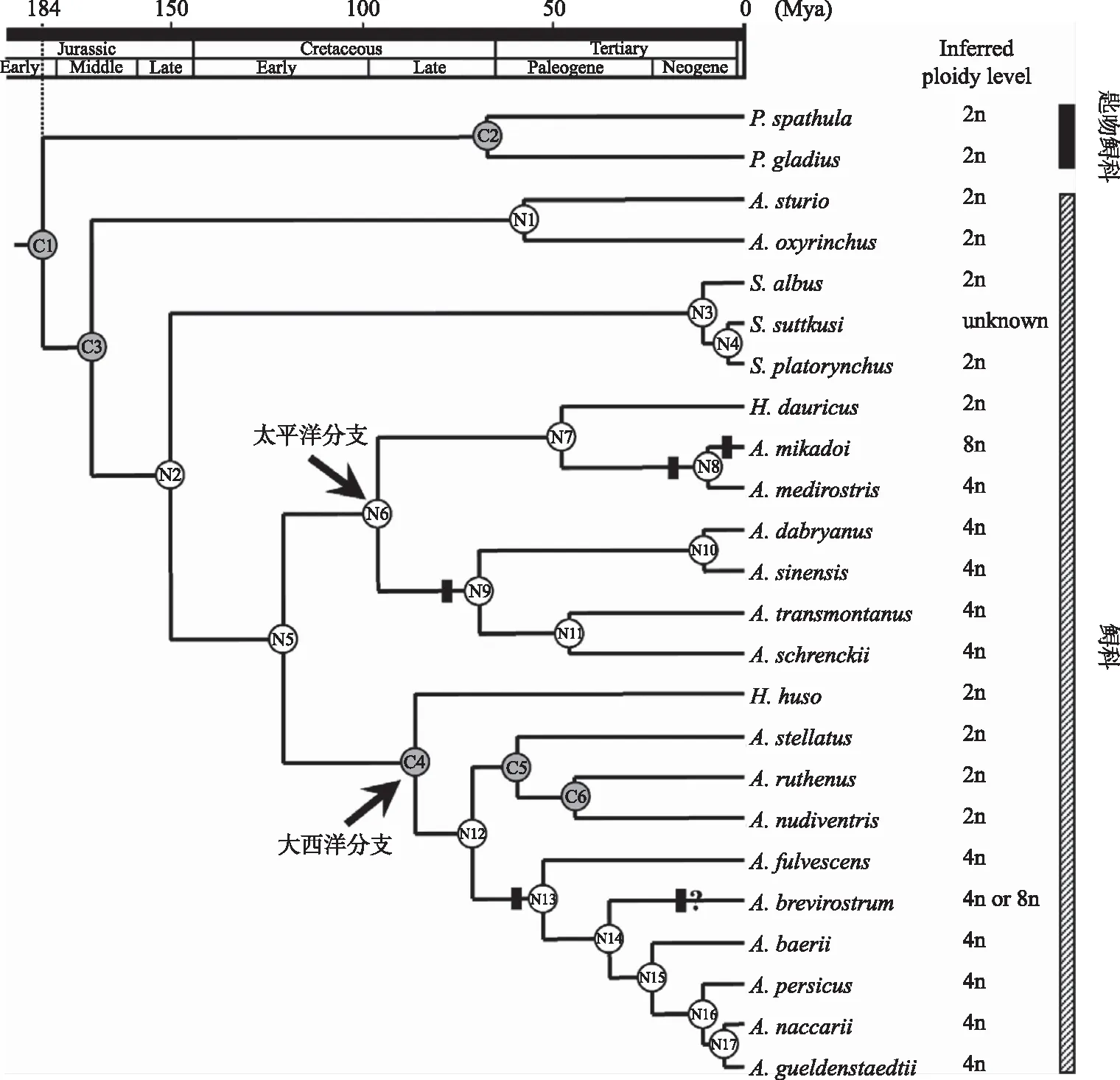
图3 根据线粒体细胞色素b构建的有根的内类群数(分支表示估算的分歧时间,C1~C6表示校验节点,N1~N17为感兴趣的时间节点,虚线表示鲟和匙吻鲟分歧时间(百万年)的后验平均值[10])
鲟鱼是典型的冷水性鱼类,寿命长,性成熟晚,体型大[11]. 譬如,欧洲鲟(Husohuso)可活118年,1827年曾经在伏尔加河河口捕获的一条雌鱼体重达到1571 kg!四川省的渔谚中有“千斤腊子万斤象”的说法,所谓“腊子”是指中华鲟,“象”则是指白鲟[12].
许多鲟鱼属于溯河产卵的底栖鱼类,在生殖季节需到河流的上游去产卵,但大部分时间在河流三角洲和河口地区摄食生长. 有些鲟鱼完全生活在淡水环境,而有些则主要栖息于沿海水域之中,虽然偶尔也能冒险进入深海.
鲟形目鱼类广泛分布于北美与欧亚大陆,通过分子系统树可分为两支——大西洋支和太平洋支(图4). 我国境内分布的野生鲟类有8种,分布于黑龙江、松花江、乌苏里江流域的史氏鲟(Acipenserschrenckii)、达氏鳇(Husodauricus)和库页岛鲟(Acipensermedirostris),分布于长江流域的中华鲟(Acipensersinensis)、达氏鲟(Acipenserdabryanus)和白鲟(Psephurusgladius),分布于新疆伊宁等地水域中的裸腹鲟(Acipensernudiventris),以及分布于新疆额尔齐斯河、布伦托海、博斯腾湖的西伯利亚鲟(Acipenserbaeri). 栖息于长江之中的中华鲟、达氏鲟和白鲟都已处于濒危状态,被列为国家Ⅰ级保护野生动物.
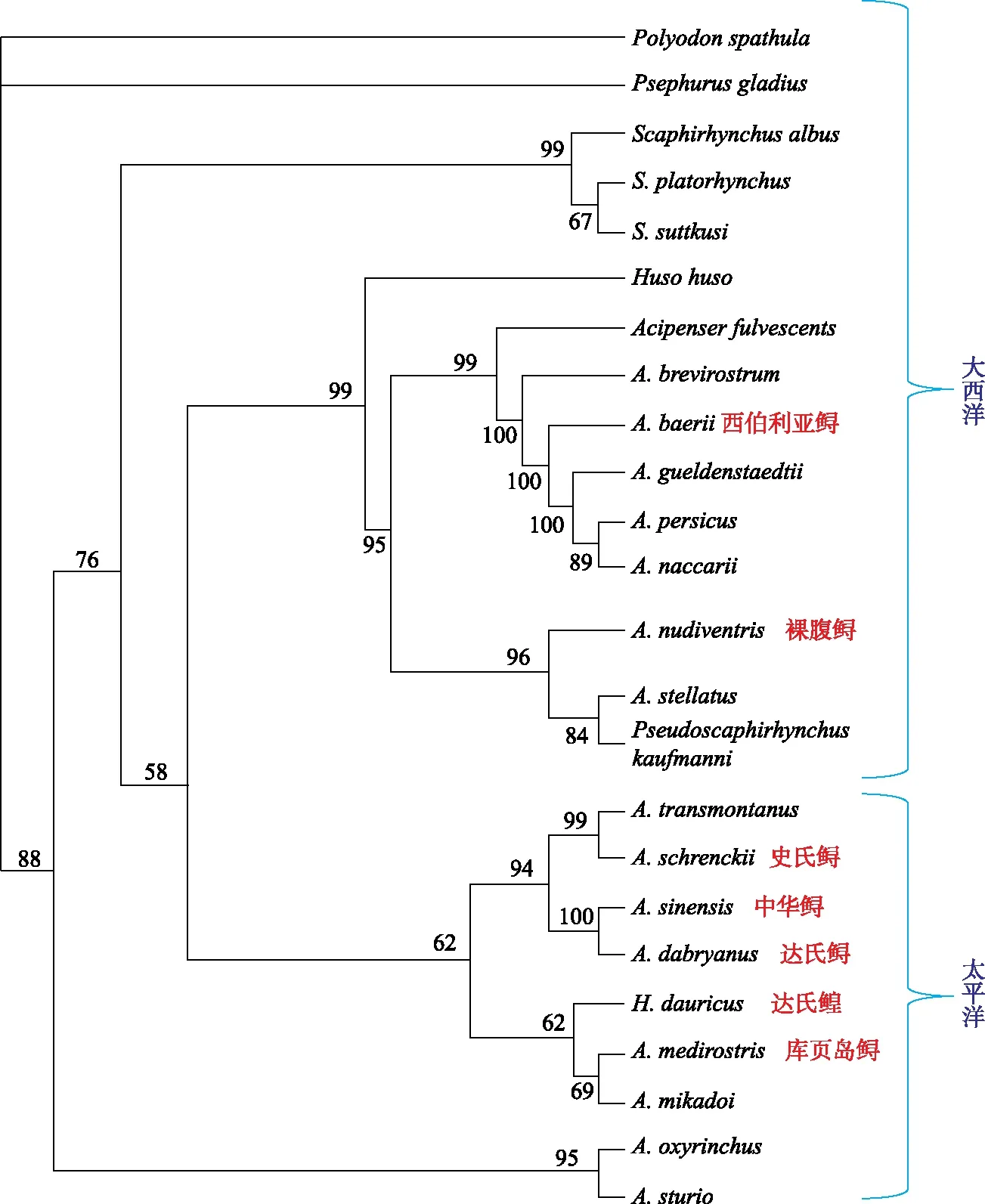
图4 根据线粒体细胞色素b、12S rRNA、细胞色素c氧化酶亚基Ⅱ、tRNA Phe、tRNA Asp、16S rRNA、NADH5和控制区基因序列(8个基因,4406个位点)构建的鲟形目鱼类的系统发育树,包含24种鲟鱼,其中7种(红色中文标识)分布于中国[13]
2 中华鲟——正在走向灭绝
中华鲟(图5)是一种海河洄游性鱼类,隶属于鲟形目鲟科鲟属. 中华鲟曾经分布于韩国西南部水域,日本九州西部水域,中国的黄河、长江、珠江、闽江、钱塘江等水域. 但现在除了葛洲坝下的长江中下游之外,中华鲟在其他河流都已绝迹.

图5 体型巨大的中华鲟(Acipenser sinensis)[14]
2.1 生活简史
中华鲟在长江上游出生,在大海中成长,最大个体曾有680 kg的纪录,寿命可达40龄. 中华鲟平时生活在黄海、东海、南海北部的浅海区域,以底栖动物(虾、蟹、软体动物、头足类、水生昆虫和小杂鱼等)为食,雄鱼最早8龄、雌鱼最早14龄才达初次性成熟[15-16]. 成熟个体需溯河洄游2500 km以上以抵达长江上游的金沙江(宜宾-屏山)河段方才进行产卵繁殖[17],那里水流湍急,底为砾石. 受精卵粘附于江底的岩石或石砾表面进行孵化.
中华鲟在秋季产卵. 亲鱼进入长江后要停留1年多时间,在此期间基本不摄食,依靠体内积累的脂肪提供繁殖过程中的能量消耗和性腺发育所需的物质,待性腺由Ⅲ期发育到Ⅳ期后,于翌年秋季10月中旬-11月中旬产卵,产后亲鱼迅速离开产卵场,降河入海肥育[18](图6). 因此,在长江中个体愈成熟,身体愈消瘦. 幼鱼长大到15 cm左右进入大海. 中华鲟在生殖洄游过程不进食而全靠消耗身体内的营养储备,其忍耐饥饿的能力是惊人的,在硬骨鱼类中亦极为罕见. 中华鲟并非每年产卵,根据对胸鳍第一鳍条磨片上的生殖标志的观察推测,中华鲟重复产卵的间隔期至少为5~7年[15].
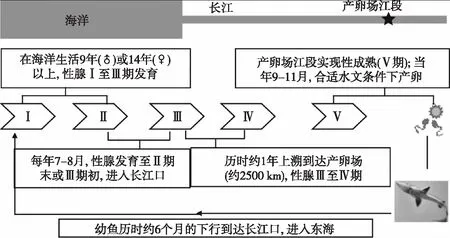
图6 中华鲟的生活史[19]
中华鲟产卵后,会有各种鱼类——圆口铜鱼(Coreiusguichenoti)、瓦氏黄颡鱼(Pelteobagrusvachelli)和铜鱼(Coreiusheterodon)等10多种底层鱼类——聚集在其播卵区,吞食黏附在河流底质上的受精卵[20]. 因此,解剖食卵鱼也是监测中华鲟自然繁殖的传统方法(图7).

图7 葛洲坝江段瓦氏黄颡鱼摄食中华鲟卵的情况(红圈中灰白色颗粒即为中华鲟卵)[21]
2.2 葛洲坝——切断了中华鲟的生殖洄游通道
早在1970s科学家就曾预言,“目前正在兴建的葛洲坝水利枢纽将阻隔亲鱼自中、下游上溯进行生殖洄游,对中华鲟的资源影响严重,如不采取适当措施,甚至将会造成长江中华鲟绝迹”[22].
据估算,在葛洲坝截流前,长江中中华鲟的有效补充量每年约1882尾,长江和海洋中育龄(雌:13~34龄,雄:8~27龄)总资源量为32260尾,其中雄鱼15310尾,雌鱼16950尾,每年长江中的资源量(捕捞后)约1727尾,占总资源量的比例约为5%[23].
葛洲坝截流前中华鲟产卵场分布在金沙江下游和长江上游约800 km的江段,已报道的产卵场有16处以上. 1981年葛洲坝建成后,阻止了中华鲟亲鱼的生殖洄游,无法上溯到金沙江产卵的中华鲟,勉强在葛洲坝下形成了新的产卵场,主要是宜昌长航船厂至十里红江段,长度约为4 km;其次是十里红至胭脂坝江段,长度约为5 km[24]. 也有一种说法是葛洲坝截流后仅形成了1个新的产卵场,位于坝下消力池至庙嘴2 km范围内,主要集中在南岸笔架山江段的1 km范围内[18]. 很显然,中华鲟产卵场的规模大幅萎缩(图8). 紧靠葛洲坝枢纽的坝下江段新形成的中华鲟产卵场被认为对延续中华鲟的自然种群会起到至关重要的作用[24]. Deng等[25]甚至认为,葛洲坝对中华鲟不是一个威胁. 但Xie[26]预测,三峡大坝建成后,可能毁坏这个唯一的产卵场. 不幸的是,2013、2015、2017年均未在葛洲坝下产卵场发现中华鲟产卵. 中华鲟自然繁殖的终结已近在咫尺!
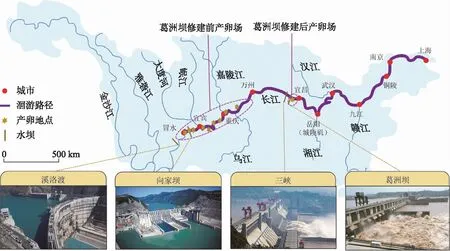
图8 长江中华鲟产卵场分布示意图[27]
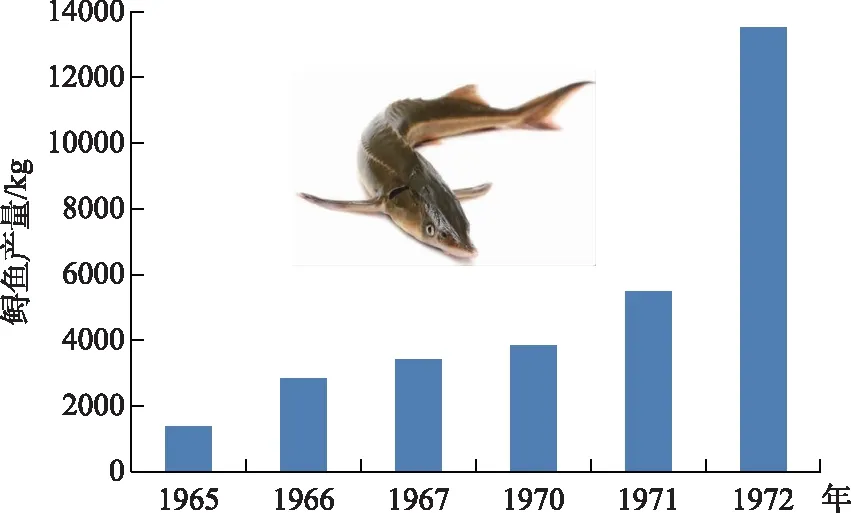
图9 宜宾渔业社1965-1972年期间的鲟鱼产量(数据来源:文献[22])
上溯到金沙江产卵的中华鲟,曾经是一种重要的经济鱼类,在长江上游的产量每年大约20000~25000 kg,譬如,在宜宾地区每年繁殖季节可形成一个捕捞中华鲟的旺季(图9). 葛洲坝截流后的1981-1982年,宜昌和宜宾等地的渔民大量捕杀中华鲟亲鱼,数量达2000余尾,给中华鲟种群带来很大的伤害[12]. 1983年以后,国家明令禁止了对中华鲟的商业性捕捞. 据黄真理等[28]最近的估算,1981年1月葛洲坝截流导致1980年鱼群被葛洲坝阻隔在上游的数量为660尾,下游数量为349尾,1981年坝下中华鲟捕捞量1002尾.
在过去的半个世纪,中华鲟的繁殖群体经历了急速的衰退过程[29]. 据估计,1970s,长江中的中华鲟繁殖群体数量(捕捞后)尚有1727尾,2005-2007年期间下降到了203~257尾,到了2010年只剩数十尾(图10). 中华鲟已被列入《世界自然保护联盟》(IUCN)2012年濒危物种红色名录ver 3.1——极危(CR).
2.3 梯级大坝——压死骆驼的最后一根稻草
三峡蓄水对温度过程产生了影响:以2004年刚蓄水的寸滩站(位于三峡水库顶部,是三峡水库的入口控制站)和宜昌站(位于三峡大坝下游42 km 处,是三峡水库的出口控制站)为例,1-2月份水温偏高,3-5月份水温偏低,7-12月又显著偏高(图11).
水温的变化被认为对中华鲟的产卵产生了重要影响:历史资料表明,葛洲坝建成前,中华鲟产卵活动大部分发生在10 月,在21次产卵中只有3 次发生在11月;葛洲坝建成后至三峡蓄水的2003 年以前,中华鲟在大部分年份有两次繁殖活动,产卵时间集中在10月中下旬;2003年以后,中华鲟在大部分年份只有1 次产卵活动,且产卵时间明显推迟(图12).
从2013年溪洛渡蓄水开始,梯级水坝强化了秋季水温的增加,导致2013年开始中华鲟停止产卵[27]. 上游大坝对下泄水温度的改变进而对中华鲟产卵的影响日益受到关注[27,32-38]. 王鲁海和黄真理[36]认为,如果通过梯级水库调度等措施在10月22日之前将葛洲坝下产卵场的水温降低到18~20℃,中华鲟野生种群数量可维持在葛洲坝截流后的水平. 但依笔者之见,下泄水温升高导致中华鲟在坝下产卵推迟,并不是导致中华鲟衰退的决定性因素,因为虽然推迟产卵,但毕竟还是能产卵.
黄真理(私信交流)认为,葛洲坝修建前,正常繁殖期在9月中旬-11月中旬(2个月),葛洲坝修建后推迟至10月中旬-11月中旬(1个月)产卵,梯级水坝修建后停止了产卵. 这里面,阻隔后性腺发育推迟37天,梯级水坝后适宜环境条件(以水温为主)从10月中旬推迟到11月底或12月初. 中华鲟在长江不摄食,性腺发育推迟后,如果推迟时间超过容许时间上限(种群水平上为11月15日),性腺就会吸收退化. 因为,中华鲟的性腺发育和脂肪储量,不容许长期等待,什么时候环境条件适合,什么时候繁殖. 就像女人生孩子,也有容许时间的上限,过了42周就有生命危险. 因此,是长江梯级水坝导致了中华鲟的可能灭绝. 要解决其中关键问题,如性腺退化、温度抑制繁殖行为等问题.
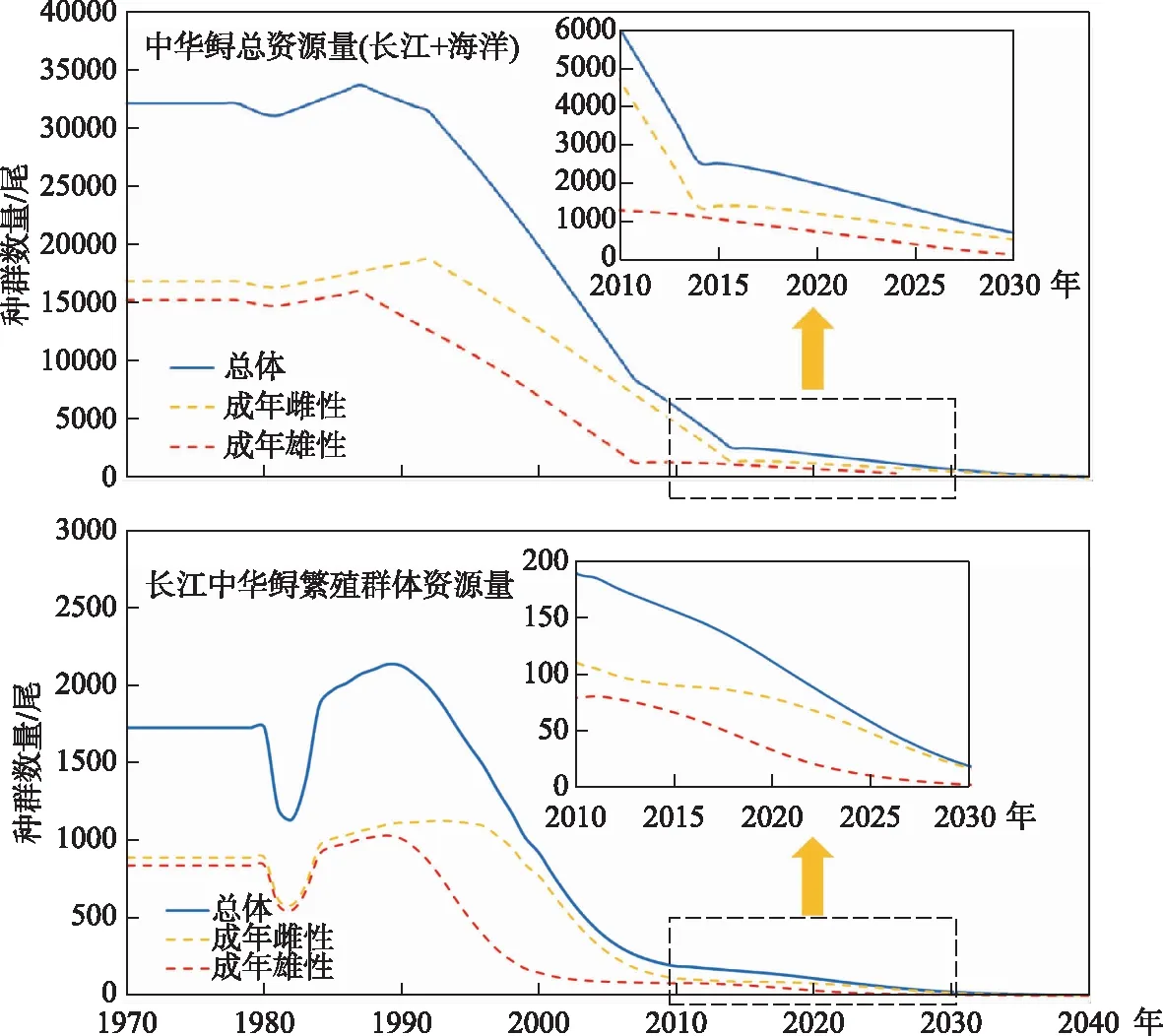
图10 自1970s以来中华鲟繁殖群体在长江中的衰退过程[27]
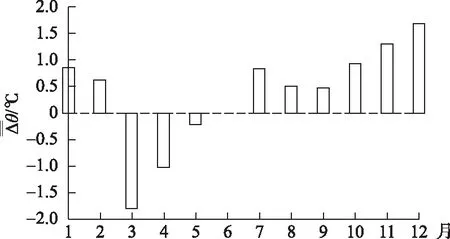
图11 寸滩站与宜昌站2004年月平均水温差与蓄水前两站多年月平均水温差之差的变化[30]
中华鲟产卵时间推迟为何就会招来灭顶之灾呢?这实在难以令人信服. 笔者认为,如果繁殖群体足够大,这应该不会成为问题. 但事实上,三峡截流前,中华鲟的种群数量(如果按IUCN的估算数据)就已衰减到不足1970s的4%,大势已去,温度变化最多不过是压死骆驼的最后一根稻草罢了!
2.4 葛洲坝下的产卵场早就失去了意义
中华鲟的最大寿命可达40龄,最大个体可达560 kg[18]. 在葛洲坝截流前的1972-1976年,金沙江下游产卵场中华鲟成熟个体雄性最小为9龄,雌性最小为14龄,初次性成熟雄性为9~22龄,雌性为14~27龄[39]. 通过繁殖群体的年龄统计来看,雌性的平均年龄比雄性高出5年左右,暗示雌性寿命可能明显高于雄性.
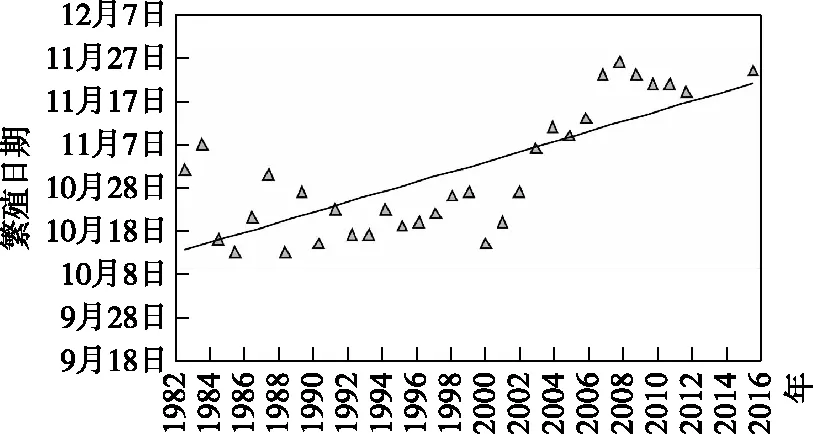
图12 中华鲟的自然繁殖时间[31]
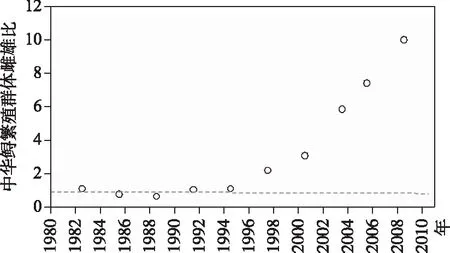
图13 葛洲坝截流之后坝下宜昌江段中华鲟产卵群体的雌雄性比变化[31]
葛洲坝截流发生于1981年1月4日,当年5月23日大坝关闸蓄水,因此1981年秋季繁殖的中华鲟才是葛洲坝截流后出生的后代. 譬如,只有在1996 年秋季采集的15 龄样本才是1981年出生的[29].
从图13不难看出,葛洲坝截流之后的新生繁殖群体补充严重缺乏,这可由雌雄之间寿命的差异来解释,即先行死亡的雄性导致了繁殖群体雌雄性比的不断攀升. 这其实是中华鲟自然种群进入灭绝状态的先兆!
2.5 宁可玉碎,不能瓦全
葛洲坝下并不是中华鲟理想的产卵之地,虽然每年都有为数不多的个体(如30~40尾)在那里产卵繁殖,但是很多被拦在坝下的亲鱼其性腺退化(雄性约30%,雌性约70%),有一部分亲鱼向坝上硬闯,撞得头破血流伤重而亡(据统计,1981-1985年期间在宜昌江段共捡到死鲟146尾,每年平均30尾左右),还有一部分亲鱼未产卵而直接降河入海[18].
葛洲坝工程局中华鲟人工繁殖研究所从1983年起,每年都向长江放流中华鲟苗种,人工繁殖放流被视为拯救中华鲟的主要措施[24]. 据统计,1983年以来的30多年间,中华鲟人工繁殖研究所每年都向长江投放鲟苗20万~50万尾,以增加长江的种群数量. 但中华鲟的种群数量并未因人工放流而增加,估计人工放流对野生种群的贡献不足10%[40-41].
为了拯救中华鲟,先后建立了3个自然保护区:江苏省东台市中华鲟自然保护区(省级,2000年)、上海市长江口中华鲟自然保护区(市级,2002年)和湖北省宜昌市中华鲟自然保护区(国家级,2004年). 2015年9月28日,农业部发布了《中华鲟拯救行动计划(2015-2030年)》.
令人遗憾的是,无论是通过人工繁殖放流还是建立自然保护区,都未能阻挡中华鲟的日益衰退,这个古老的鲟鱼似乎已经卷入了难以逃脱的灭绝漩涡!

图14 达氏鲟[14]
3 达氏鲟——危危可及
达氏鲟(Acipenserdabryanus),又名长江鲟、沙腊子,隶属于鲟形目鲟科鲟属,仅分布于长江中、上游,尤其以宜昌与宜宾之间的干流江段为多,在上游金沙江、沱江、嘉陵江等支流和洞庭湖亦有记载[22]. 达氏鲟是我国的特有物种,属淡水定居型鲟种. 达氏鲟可能是中华鲟的陆封型类群(landlocked form)[42].
3.1 生活简史
达氏鲟(图14)喜欢在较暗的底层缓流水体中活动,常栖息于8~10 m的江河浅水区,在岩礁、沙底、卵石区域游弋与觅食. 冬季在缓流的河道中越冬,立春后当江水回升至12℃以上时开始溯水到上游干支流摄食与繁殖[43].
达氏鲟雄性4~7龄、雌性5~8龄达性成熟,一般体长0.8~1.0 m,体重5~10 kg. 每年春季(3-4月)和秋季(11-12月)产卵,适宜水温为16~19℃,绝对怀卵量6万~13万粒[39]. 性成熟个体上溯至长江上游(重庆至宜宾江段)繁殖,卵粘着在石砾滩底上发育,最适孵化温度为18~20℃[44]. 与中华鲟和白鲟相比,达氏鲟体型要小得多,记载的最大个体也才16 kg[18].
达氏鲟的食物主要是底栖无脊椎动物(摇蚊幼虫、蜻蜓幼虫、蜉蝣幼虫和蛹、蚬、寡毛类等),也摄食水生植物碎屑、藻类(硅藻和丝状藻类)和腐殖质等,有少数个体捕食鱼苗和幼鱼[22].
3.2 种群的衰退与濒危
达氏鲟的自然种群并不大,虽然它曾是长江上游的重要经济鱼类,但1980s以来,达氏鲟的种群数量已十分稀少. 1984-2010年的统计数据显示,每年平均仅有十多尾达氏鲟被误捕,特别是1995年以来,在葛洲坝下就再也没有捕捞到过[45-46]. 由此判断,达氏鲟在葛洲坝下游江段可能已经绝迹,目前仅零星分布于四川境内的长江上游江段.
导致达氏鲟种群衰退的原因有过度捕捞和水利工程等. 譬如,在沱江的内江河段,1950s仅有渔船500艘,到1985年增加到2000艘. 又如,在岷江乐山河段,流刺网不分昼夜密集作业. 1981年建成的葛洲坝和2003年建成的三峡大坝大大压缩了达氏鲟的生存空间,迫使它们只能生活于坝上江段,而2008年建成的向家坝正好在达氏鲟产卵场的中间,其巨大影响不言而喻.
1988年达氏鲟被列为国家Ⅰ级保护野生动物. 1996年,世界自然保护联盟(IUCN)将达氏鲟评定为极危(critically endangered, CR)物种. 据称,自2000年以后再没有监测到达氏鲟的自然繁殖,达氏鲟的野生种群基本绝迹,人工保种的野生个体仅存几十尾. 2018年5月16日,农业农村部发布了《长江鲟(达氏鲟)拯救行动计划(2018-2035年)》. 人工放流依然延续着人们企图恢复达氏鲟野外种群的梦想,如2019年5月21日,中国水产科学研究院长江水产研究所在长江上游的宜宾江段放流了60尾达氏鲟亲本、1000尾2龄达氏鲟.
4 白鲟——已经绝灭?
白鲟(Psephurusgladius)隶属于鲟形目匙吻鲟科白鲟属,主要分布于四川宜宾以下至河口的长江干流和支流(沱江、岷江、嘉陵江)以及大型通江湖泊(洞庭湖、鄱阳湖)中,也见于黄河、钱塘江和甬江,东海和南海亦有分布[18].
早在1862年,英国“扬子江上游探险队”的托马斯·布莱基斯顿在湖北咸宁市附近记述了白鲟:“体长3英尺,嘴部呈钟形,向外突出,鼻子或触角伸出头部12英寸”,并绘有白鲟整体及部分的素描图[47].
4.1 生活简史
白鲟是中国最大的淡水鱼类,个体长度一般在2 m左右,重量在25 kg以上. 白鲟个体的最大记录体重为908 kg[54],体长为7 m,是世界淡水鱼类体长的最高记录.
白鲟(图15)分布于长江、钱塘江以及黄海和东海[48]. 在长江干流,白鲟栖息于中下层,偶亦进入沿江大型湖泊中,大的个体多栖息于干流的深水河槽,幼鱼则常到支流、港道,甚至到长江口的半咸水区觅食.

图15 已经绝灭的世界上最长的淡水鱼——白鲟(Psephurus gladius)[14]
白鲟为凶猛性鱼类,成鱼和幼鱼均以鱼类为食:在重庆、涪陵解剖的8尾鱼中均以铜鱼为食,在安庆解剖的5尾鱼主要摄食鲚鱼以及少数虾、蟹[22].
白鲟初次性成熟年龄较迟,雄鱼为5~6龄,雌鱼为7~8龄. 每年春季(3-4月)溯江产卵,产卵场位于四川省泸州以上,集中于江安县附近的长江河段和宜宾柏树溪附近的金沙江河段. 成熟亲鱼多栖息在水流较急、水较深、底质多为岩石或鹅卵石的河段[49]. 白鲟产沉性卵,产卵水温为18~20℃. 7~15龄雌性白鲟的绝对繁殖力为17.8万~78.8万粒(平均为35.9万粒)[50].
4.2 种群的衰退与濒危
在上游繁殖的白鲟一部分滞留在上游干支流内生长,另有一部分漂流到中下游,有的一直抵达长江口崇明附近觅食. 葛洲坝枢纽兴建后,在长江上游生活的白鲟仍能繁殖,但在中下游成长的白鲟无法上溯到上游产卵场,滞留于葛洲坝宜昌江段的白鲟虽然能够发育成熟,但由于亲鲟数量很少,未观察到在坝下江段的自然繁殖;在上游繁殖的一部分白鲟仔、幼鱼可以通过泄水闸漂流下坝,因此,在1983-1988年期间,每年均可在崇明收集到白鲟幼鱼,但数量呈逐渐减少的趋势[24].
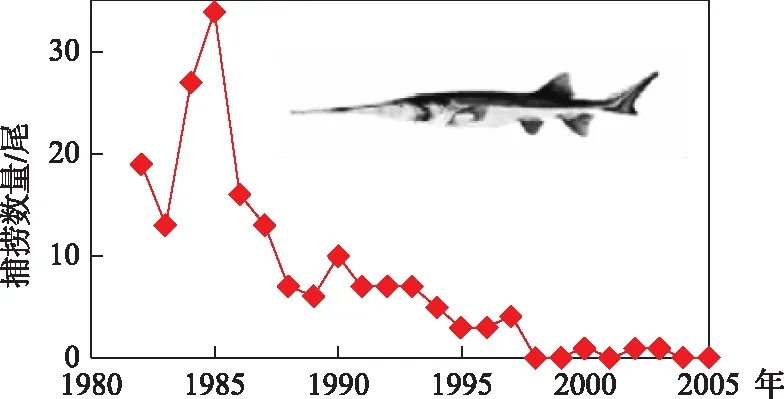
图16 1981-2005年期间葛洲坝下的白鲟捕捞数量[51]
白鲟的产量曾达25 t左右,但1980s以来,种群数量大幅下降(图16). 白鲟已很难捕获到,1992年在长江上游重庆江段捕获了2头,2002年在长江下游江苏南京江段捕获到一尾雌鱼,2003年在长江上游的宜宾江段捕获了一尾长3.5 m的雌鱼. 之后就再也没有见到过白鲟. 据估计,白鲟于1993年功能性灭绝(functional extinction),2005-2010年期间完全灭绝[51].
1983年白鲟被列为国家I级野生保护动物,1996年,IUCN将白鲟评定为极危物种. 水利工程对生殖洄游的阻断、过度捕捞以及长江鱼类资源量的下降可能是导致白鲟濒危的重要因素.
5 为什么长江的鲟鱼都岌岌可危?
鲟鱼虽然古老,但却十分稀少. 譬如,全世界的鲤形目鱼类有3000多种,而全世界的鲟形目鱼类却只有约27种. 这表明鲟鱼的分化能力极其有限,也预示其生态位的特殊性,以及极为有限的生态可塑性. 鲟鱼的珍贵已是不争的事实(图17).

图17 1994年发行的鲟鱼(鳇、中华鲟、白鲟和达氏鲟)邮票
虽然在长江中的3种鲟鱼的生活史特征或多或少都有些差别(表1),但繁殖方面却有惊人的相似之处:产卵场都在上游相似的江段,都是激流环境,底质特征都是多砾石,都产沉性·粘性卵,虽然产卵时间不同——达氏鲟和白鲟在春季,而中华鲟在秋季. 三种鲟鱼寻求在上游的急流环境中产卵,难道是因为能从那里得到什么欢乐?这应该是一种自然选择的结果,即它们嗜好的可能不是激流,而是急流环境中的砾石,因为它们产出的卵都需粘附在石块上或沉落于石缝中去孵化,这可能最有利于安全孵化与幼鱼成活,就被固化成了秉性. 而在葛洲坝下,鲜有江段能满足这样的条件,因此,自1980s初葛洲坝建成以来,鲟鱼就迎来了灭顶之灾. 虽然中华鲟在葛洲坝下形成了新的产卵场,但其规模却大大萎缩,现在看来,这个新的产卵场未必就那么适合中华鲟.

表1 长江中3种鲟鱼生活史特征的比较
水坝对长江鲟鱼的致命打击无容置疑[56],从地质历史演化的角度看亦是如此[57],因为适应并不是万能的,不然,历史上就不会出现物种的绝灭,这与物种自身的适应能力、时间以及环境的变幅等诸多因素均有关系. 成熟的中华鲟并不需要在长江中摄取食物(虽然它在浅海的索饵场或多或少也遭受到了破坏),因此,中华鲟种群的衰退可能只能归因于生殖洄游的阻隔. 它们无法在葛洲坝上游长期生存下去,因为它们的幼鱼必须回到大海中去摄食与肥育. 而在葛洲坝下游,又没有它们适宜的产卵之地. 因此,笔者悲观地预测,中华鲟的灭绝决不会久远,这个古老的物种已无回天之力.
因为达氏鲟无需到大海中去肥育,从理论上来说,只要上游的产卵场存在以及饵料充足,它就可能在上游水系中残存下来. 但因为上游的个体无法到中下游及其通江湖泊中摄食,达氏鲟的生存空间已被大大压缩. 白鲟属于咸淡水双向迁徙型,早期幼鱼需要在海洋生活(河口捕捞到幼鱼),幼鱼生长到一定阶段回到长江. 因此,一方面葛洲坝对它的影响会很显著,另一方面又由于它体型更大,且是肉食性,因此,灭绝的风险更大. 事实上,现在很难寻觅白鲟的身影,被认为已经功能性灭绝了.
鲟鱼体型巨大,一个完整的生活周期往往需要巨大的空间,特别是有些环境的不可替代性往往会成为它们的致命之处,这种关键环境的破坏往往会导致灭顶之灾,如中华鲟体型巨大,其生活周期需洄游数千里,而且还需到金沙江的激流中去繁殖,将卵粘附于砾石或掉落于砾石缝隙之中. 其他两种鲟鱼也选择了类似的产卵环境.
水温、水流以及砾石……哪一个都有可能成为这些古老鲟鱼的繁殖启动信号,也许是它们的组合,但这些环境信号的一致性所表征的产卵场却不是那么轻易能够重组的,因为它是在漫长的自然历史中,鲟鱼适应气候、地质过程以及河流地貌不断演变的产物,藉此,中华鲟形成了对河流自然进程的特殊感知,并如此根深蒂固以致无可替代. 因此,葛洲坝与三峡大坝的阻隔,使中华鲟永远丧失了适宜的产卵场所. 白鲟的生存空间似乎也是从上游一直延伸到河口,只是不像中华鲟那样必须到大海中去摄食生长,因而但大坝的阻隔也使它们索饵肥育的空间大大压缩. 达氏鲟由于体型相对较小,也主要在中上游索饵生长,因而可能是在3种鲟鱼中生存状态相对较好的一种,或许是最有可能在上游环境中残存下来的鲟鱼.
这是一条延绵数千公里并通向海洋的河流,从古至今都是所有物种共享着的,河首河尾,河滩河心,此处彼处……在这个壮观的剧院中,它们用各自的音符弹奏着一首共同的河流之歌,荡漾在无尽的河谷之中,即便我们倾心地观察与思考,也未必能完全理解它. 中华鲟和白鲟的乐曲就谱写在大海与金沙江之间的穿越中,保持着几乎不变的生死韵律——觅食、洄游、繁殖和死亡——世世代代,生生不息……在苦难面前,它们没能屈服,其结果,曲终鲟散正在悄然而至,感知绝技即将化为寓言!
