γ-氨基丁酸的研究与应用进展
2020-07-14
(济南大学 生物科学与技术学院,山东 济南 250022)
γ-氨基丁酸(GABA)是一种天然的非蛋白质氨基酸,广泛分布在动植物、微生物中[1],由于物种不同,因此其所发挥的作用也不同。在哺乳动物体内,GABA是重要的抑制性神经递质[2],不仅参与多种代谢活动,还具有治疗癫痫[3]、预防肥胖、镇定安神[4]、健肝[5]、利肾[6-7]、促进乙醇代谢[8]、增强脑活力[9]、调节激素代谢[10]、降血压[11]等功效。GABA在植物中的主要作用是抵抗环境胁迫[12-13],在微生物中的主要作用则是提高微生物的耐酸能力[14]及孢子的萌发效率[15]。本文中主要从GABA的产生途径、谷氨酸脱羧酶(GAD)的分子结构及酶学性质、GABA的生产技术等几方面进行综述,以期能为其生产工艺的改进和完善提供参考。
1 GABA的产生途径
在生物体内,GABA产生途径有2种,分别是GABA支路和多胺降解途径[16]。其中GABA支路广泛存在于动、植物及微生物中,是生物体中产生GABA的主要途径。该支路是三羧酸循环的一条支路,三羧酸循环中的α-酮戊二酸经转氨产生L-谷氨酸(L-Glu),L-Glu在GAD的作用下脱羧生成GABA,在发挥生物学功能之后,可以通过GABA转氨酶转氨,进而氧化为琥珀酸,再进入三羧酸循环(见图1)。然而,当植物组织受到逆境环境刺激时,GABA支路产生的GABA不足,多胺降解途径作为GABA生产的辅助途径在植物体中出现。该途径是植物细胞内的腐胺、多胺在二胺氧化酶或多胺氧化酶的作用下经脱氨、脱氢生成GABA(见图2)[16-17]。

图1 γ-氨基丁酸支路

图2 多胺降解途径
GABA支路作为生物体中产生GABA的主要途径,GAD在该支路中是产生GABA的关键限速酶,因此在提升GABA产量中起到了关键作用。
2 谷氨酸脱羧酶
GAD是一种磷酸吡哆醛(PLP)类酶[18],是催化L-Glu脱羧生成GABA唯一的关键限速酶[19],对L-Glu具有高度专一性[20],对温度敏感[21],在pH大于6.0或小于2.0时[22]都会失活。
2.1 谷氨酸脱羧酶的分子结构
2.1.1 酶蛋白
生物体中的GAD为胞内酶,一般由多个相同的亚基组成,不同物种GAD的亚基数量不同。研究表明,GAD是一种由酸胁迫诱导产生的胞内酶,大肠杆菌(Escherichiacoli)的GAD为六聚体[23],短乳杆菌(Lactobacillusbrevis)、酿酒酵母(Saccharomycescerevisiae)、红曲霉(Monascuspurpureus)的GAD均为二聚体[24-26],屎肠球菌(Enterococcusfaecium)的GAD为单体[27],它们亚基的结构基本相同,但是亚基的氨基酸组成会因物种不同而有差异[28]。GAD单个亚基里含有1个分子的PLP辅酶[29],此外,该亚基的多肽链大约有480个氨基酸残基[15],由N-端结构域、PLP结合区域以及C-端结构域3个部分组成(见图3),PLP结合区域存在一个可移动的催化环(图3中用虚线标出),当PLP与催化环的氨基酸残基结合时,催化环发生位移将PLP拉入PLP结合区域使GAD具有催化活性[30]。生物体可以通过PLP和催化环的解离与结合来调节GAD的活性[31]。
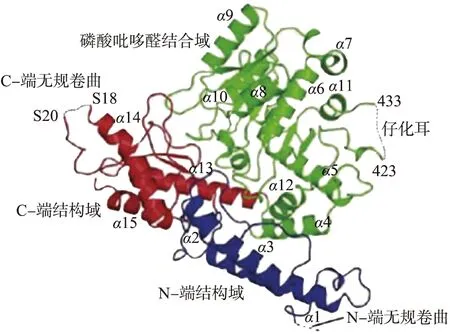
图3 谷氨酸脱羧酶单个亚基结构
齐育平等[32]以其实验室筛选出的短乳杆菌Lb-2菌株的互补脱氧核糖核酸(cDNA)为模板克隆了GAD基因。他们利用LaserGene序列工具软件、NCBI blast、ExPASy数据库及GOR Ⅰ软件对Lb-2菌株的GAD基因进行了分析预测,并利用LaserGene中的ORF Finder及相应软件分析预测了GAD基因的氨基酸组成及序列(见表1、图4),利用GOR I预测了GAD单个亚基的二级结构(见图5)。结果显示,该亚基共有468个氨基酸残基,α-螺旋、β-转角、延伸链、无规卷曲分别占总数的51.71%、8.12%、31.20%、8.97%;利用SWISS-MOD-EL软件预测了短乳杆菌Lb-2中GAD单个亚基的三级结构模型(见图6)。
2.1.2 活性中心
GAD活性中心包括PLP、GAD多肽链PLP结合区域的第276、464位的赖氨酸残基(Lys276、Lys464),第163位的谷氨酰胺残基(Gln163),第243位的天冬氨酸残基(Asp243),第245位的丙氨酸残基(Ala245),第275、465位的组氨酸残基(His275、His465),第463位的苯丙氨酸残基(Phe463)。PLP与Lys276通过共价键结合(见图7),形成一种棒状结构,PLP再以分子间氢键与活性中心其他氨基酸残基结合,形成稳定的结构支撑见图8[33]。
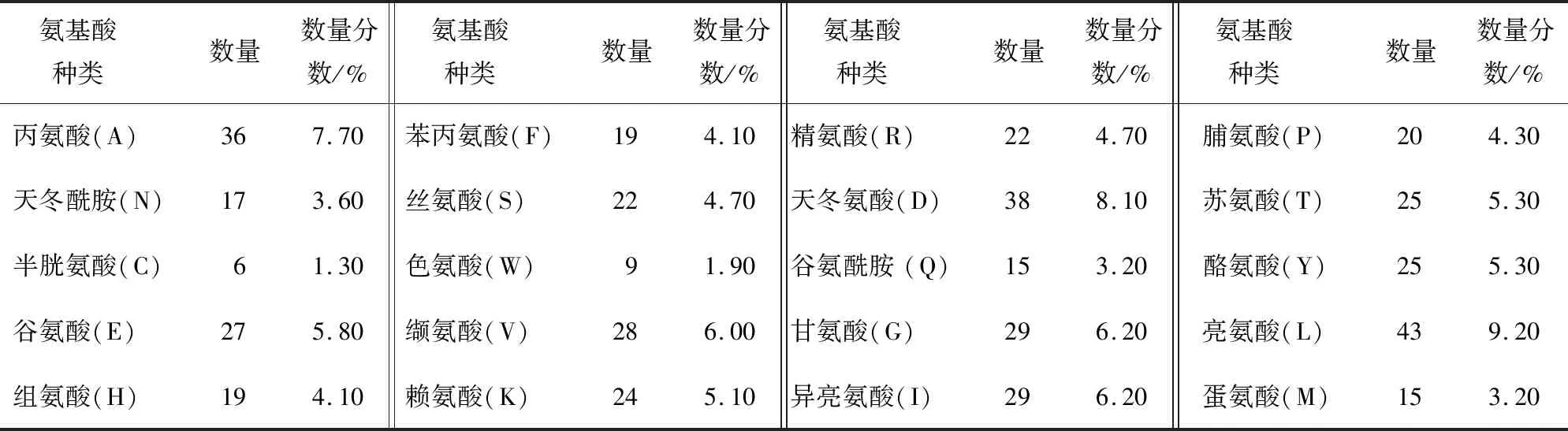
表1 短乳杆菌Lb-2中谷氨酸脱羧酶的氨基酸组成

图4 短乳杆菌Lb-2中谷氨酸脱羧酶基因及氨基酸序列
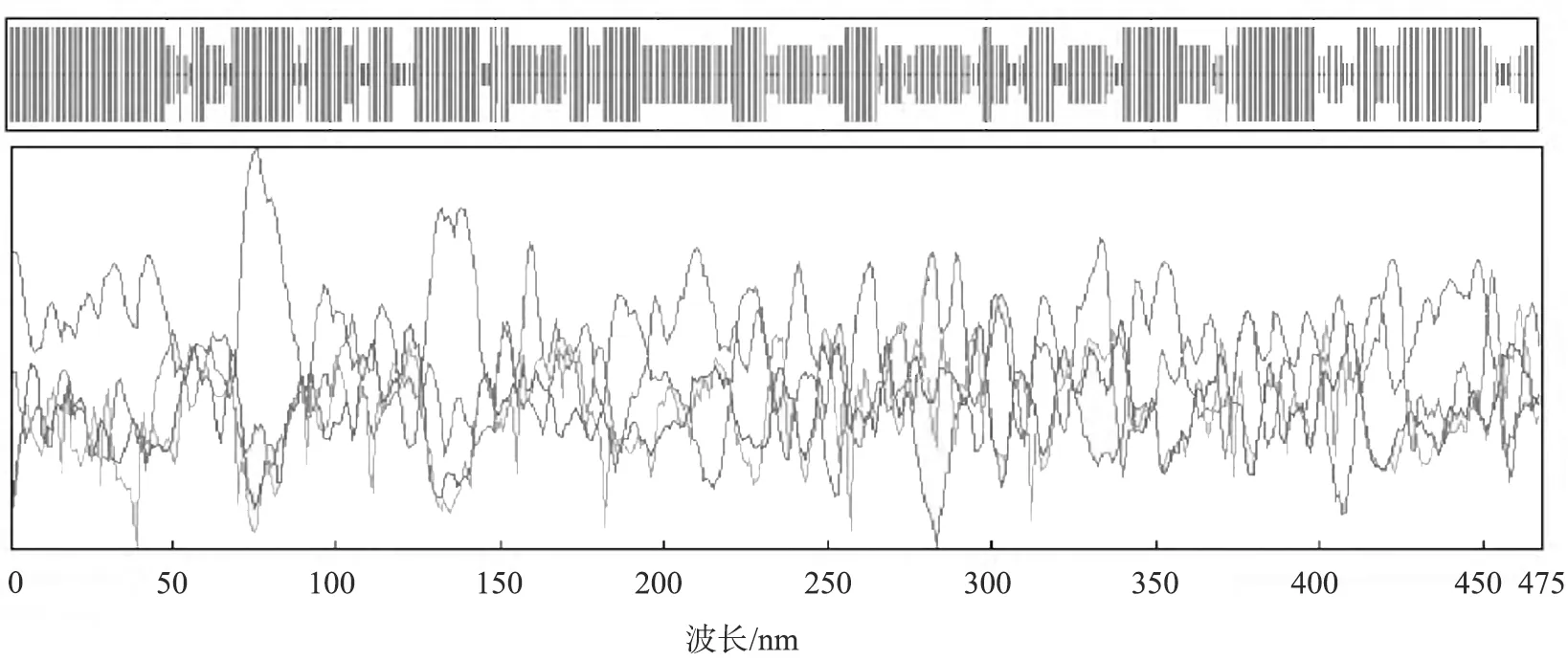
图5 GOR I预测的短乳杆菌Lb-2的谷氨酸脱羧酶二级结构图谱
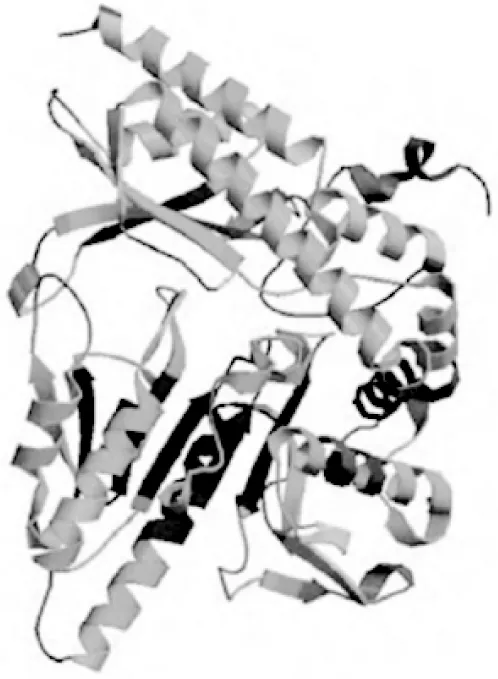
图6 短乳杆菌Lb-2的谷氨酸脱羧酶三级结构模型
当L-Glu或其盐进入GAD的活性中心时,PLP与Lys276结合的共价键断裂,L-Glu分别以共价键与PLP、Lys276结合,进行α-脱羧反应。PLP-Lys276棒状结构对pH敏感,光谱学性质差异很大,会影响L-Glu在活性中心的结合,从而影响GAD的活性。相比之下,Thr466与活性中心的PLP结合,则会“锁住”活性中心,阻止L-Glu及其盐进入活性中心,从而降低GAD的活性[34]。
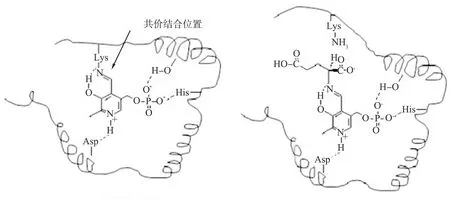
Asp—天冬氨酸残基;Lys—赖氨酸残茎;His—组氨酸残基。图7 磷酸吡哆醛、谷氨酸与谷氨酸脱羧酶活性位点中赖氨酸残基的结合方式

His275、His465—第275、465位组氨酸残基;Lys276、Lys464—第276、464位赖氨酸残基;Phe463—第463位的苯丙氨酸残基;Gln163—第163位的谷氨酰胺残基;Asp243—第243位的天冬氨酸残基;Ala245—第245位的丙氨酸残基。图8 磷酸吡哆醛与相邻氨基酸残基之间的分子内相互作用
2.2 酶促反应
GAD的辅酶是PLP[35],α-脱羧反应的最适宜pH为4.0~5.0[36],最适宜反应温度为40~50 ℃[37]。GAD可以催化进入其活性中心的L-Glu或其盐发生不可逆的α-脱羧反应[38]。L-Glu进入菌体内,与GAD活性中心的PLP结合形成醌类结构,在pH为3.5~6.0时,在转醛亚胺的作用下形成Schiff碱,之后L-Glu羧基上的1个电子进入吡啶环,Schiff碱发生质子化,L-Glu脱去1个分子的CO2得到H+,α-脱羧反应完成,L-Glu脱羧成为GABA。
GAD在微生物不同的生长周期中表达量差异很大,稳定生长期表达量最多,而指数生长期几乎不表达[39]。激活GAD的方式很多,骤冷、骤热、组织损伤、低氧、缺水、高渗透压等逆境协迫和添加激活剂都可以使GAD活性增大[40],GAD的激活剂有很多[41],以Ca2+为代表的金属离子激活剂最为常见[42],同时Cu2+、Ag+、Zn2+、Mg2+、Mn2+、Li+、Ba2+等金属离子也可以使GAD活性增强[43-45]。
3 GABA生产及应用
GABA生产方法主要有3种,分别是化学合成法、植物富集法、微生物发酵法,它们在实际生产中的应用各不相同。本文中将对各种生产方法进行详细论述。
3.1 化学合成法
化学合成法生产制备GABA的工艺有很多,主要可以分为以下3种:其一,碱性条件下,2-吡咯烷酮(C4H7NO)发生开环反应,再经过脱色、重结晶、提纯得到GABA[46];其二,高温下,邻苯二甲酞亚氨钾(C8H4KNO2)和γ-氯丁氰(C4H6ClN)反应,之后经过回流、结晶、提纯得到GABA[47];其三,γ-丁内酯(C4H6O2)和氯化亚砜(SOCl2)反应,经过氯化、酰化、氨解、皂化合成GABA[48]。以上工艺生产的GABA纯度高;但生产过程能耗高,对温度要求严格,污染环境,安全性差,成本高,不符合绿色生产的理念,因此这些生产工艺不被业内所推崇。
3.2 植物富集法
植物富集法是通过植物组织的应激代谢来富集GABA[49-50]。通常植物体内富集的GABA含量较低,分离提纯很难。例如经过富集处理的GABA茶中GABA的质量分数仅有0.401%[51],发芽的糙米中GABA的质量分数仅有0.042%[52],因此植物富集法不适用于大规模生产GABA,只适用于GABA富集产品的开发,例如米类、豆类、麦类等粮食及其他经济作物在经过富集后,可以直接加工成食品、功能性饮料以及牲畜的饲料[53-55]等。该生产方法只需分离纯化GABA,就可以满足当今消费者的需求,也是粮食深加工的一个潜在领域[56]。
3.3 微生物发酵法
微生物发酵法是利用微生物体内的GAD将L-Glu或其盐脱羧转化成GABA[57]。该方法具有专一性高、设备简单、环保、成本低、生产菌种多样且易获得等优点,适用于大规模产业化生产[58]。常见的能够产生GABA的菌种如表2所示,大肠杆菌发酵液中的GABA质量浓度最高达到了297 g/L,乳酸菌发酵液中GABA质量浓度达到了76.36 g/L,而酿酒酵母、红曲霉、屎肠球菌GABA的产量很低,所以目前常用的GABA生产菌种是大肠杆菌和乳酸菌。卫生部公布的食品新资源目录中,生产GABA的菌种是乳酸菌,因此大肠杆菌仅能作为饲料级GABA的生产菌种。
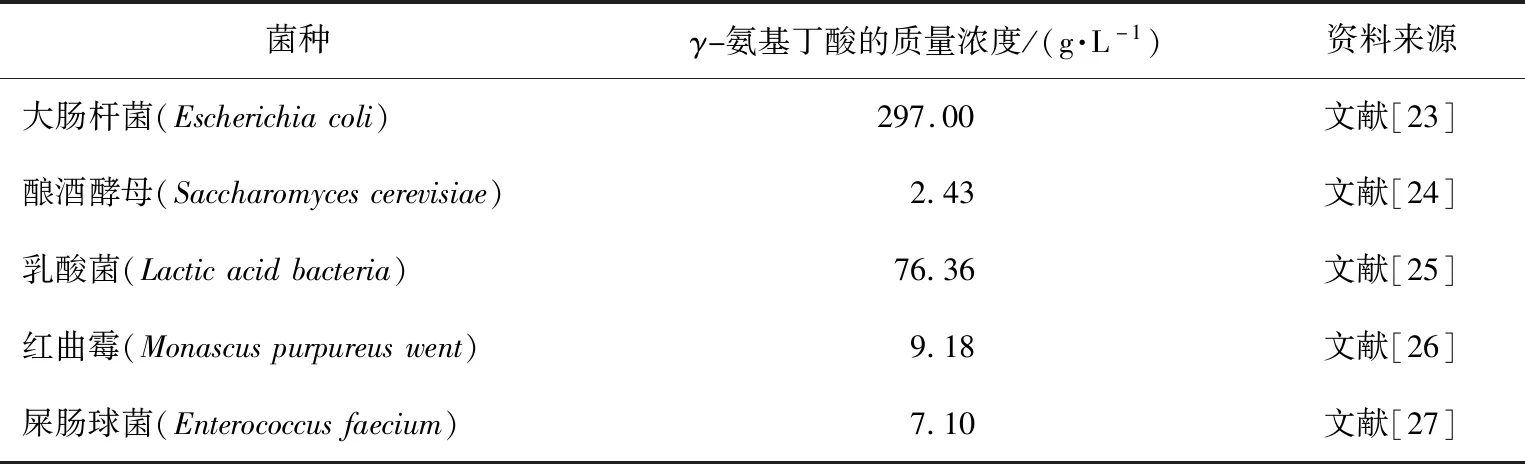
表2 能够生产γ-氨基丁酸的菌种及其产量
毕金丽等[59]分别对大肠杆菌BL21、DH5a的GAD基因进行了定点突变,构建了突变型转基因工程菌。在突变型与野生型大肠杆菌具有相同酶活力的前提下,突变型GAD具有更广泛的pH(3.4~6.2)适应能力,这样的突变型GAD将更加符合工业生产的需要,可以有效减缓GABA发酵液对生产设备的腐蚀,在饲料添加剂等行业生产实践意义重大。李超等[60]发现在持续高温的夏天,在肉鸡的每千克日粮中如果加入50 mg的GABA,可以调节肉鸡的内分泌机制[61],增强免疫力,降低肉鸡的直肠温度,减缓肉鸡的热应激反应,从而提高肉鸡的存活率和鸡肉品质。陈强等[62]发现如果在母猪的每千克饲料中添加60 mg GABA,经对比分析,添加GABA组比未添加GABA组母猪的泌乳量增加了32.85%,仔猪成活率增加了23.35%,而GABA对母猪整个的生殖周期无显著影响,这也为GABA饲料的进一步应用提供了参考。
乳酸菌作为最常用的菌种,长期用于乳制品、泡菜等食品行业[63-64],被归类为“一般公认安全菌种” (GRAS级),是理想的GAD基因和细胞工厂来源[65-66]。汪祥燕等[67]利用纸层析法在泡菜中筛选出一株短乳杆菌Y-3,该菌种发酵液中GABA的质量浓度达到了12 g/L。黄俊[25]利用短乳杆菌hjxj-08119进行了小试生产,经4次补料,发酵72 h后,发酵液中GABA的质量浓度达到了76.36 g/L。
以上这些研究都表明,与其他2种方法相比,微生物发酵生产GABA更具有开发前景,也更值得深入研究。
4 结论
GABA分布广泛,功效众多,GABA支路是其产生的主要途径。GABA支路中生产GABA的关键限速酶是GAD,该酶是由一个或多个相同的亚基组成,每个亚基的结构以及活性中心都基本相同。GAD可以通过激活剂以及逆境刺激激活使GABA的产生量增加。GABA的生产方法主要有3种,化学合成法生产GABA安全性差、成本高,植物富集法生产的GABA产量低、分离提纯困难,而通过微生物发酵法生产的GABA不仅产量高,而且安全性好,应用前景广阔,因此,研究开发GAD活力更高、生长周期更短、GABA产量更高的安全菌种以及更加简便的GABA分离提取设备,是产业化生产GABA的研究热点,具有重要的应用价值。
