外源水杨酸对高温胁迫下金莲花幼苗生理及电阻抗参数的影响
2020-07-14刘金玉高月皓黄金硕
刘金玉 高月皓 黄金硕 张 芹
(河北农业大学园林与旅游学院,保定 071000)
近年来,温室效应不断加剧,全球气候变暖,我国各地区的平均气温逐年升高,尤其是夏季持续高温已经成为许多喜冷凉植物生长发育的制约因素。金莲花(TrolliuschinensisBunge.)为毛茛科(Ranunculaceae)金莲花属(Trollius)多年生草本植物,其花叶秀美,可用作观赏植物[1],花可以入药,也是我国重要的传统药用植物[2~3]。因其市场需求量大,近年来金莲花野生资源遭到严重破坏,处于濒危状态。因此开展金莲花引种繁殖,扩大栽培范围,对满足市场需求及保护野生金莲花资源具有重要意义。但金莲花性喜冷凉湿润、阳光较为充足的环境,最适生长温度为18~24℃[1,4],引种到北方低海拔地区栽培时幼苗越夏生长不良,因此,研究提高金莲花耐热性的措施成为栽培引种中亟待解决的问题。水杨酸(SA)作为一种外源调节物质,在提高植物抗逆性方面已有许多报道[5~11]。但有关金莲花的研究多集中在种子萌发、栽培管理、药用成分、医药制剂等方面[12~14],外源水杨酸对金莲花耐热性的影响尚未见报道。本试验以金莲花盆栽苗为材料,研究外施不同浓度的水杨酸对高温胁迫下金莲花幼苗生长及生理的影响,旨在为提高金莲花的耐热性提供理论和技术指导。另外,电阻抗图谱法(EIS)是由等效电路代表所测定植物器官样本,可快速获得植物基本生理学信息[15],在评测植物抗逆性方面更加便捷[16~19]。本研究中采用EIS结合生理指标评测金莲花的耐热性,为今后应用一种快速鉴定金莲花耐热性的方法提供依据,也为其他植物抗逆性鉴定提供参考。
1 材料与方法
1.1 试验材料与处理
试验于2017年3~9月份在河北农业大学实验基地进行。供试材料为当年生的金莲花幼苗,栽植于10 cm×10 cm的花盆中,每盆1株,栽培基质为草炭∶珍珠岩=3∶1,置于温室内进行统一常规管理。待植株生长状况较稳定时,选取长势一致、健壮无病虫害的金莲花幼苗进行试验。外源水杨酸(SA)设置4个浓度梯度(0.5、1.0、1.5、2.0 mmol·L-1),试验前首先喷洒蒸馏水清洁苗体,滤纸吸干水分,然后分别采用不同浓度的SA于上午8点喷施幼苗叶片正反两面,以喷施蒸馏水为对照,以叶片上液体欲滴为宜,连续喷施2 d,保证幼苗充分吸收。然后将处理过的幼苗置于PGX-3500-DN型人工气候箱进行6~72 h高温胁迫,温度设为昼38℃/夜30℃,湿度为50%~70%,光照强度为6 000 lx,之后常温25℃恢复生长4 d。每处理3次重复,每个重复10株幼苗。分别于处理6、12、24、48、72 h及恢复生长4 d后进行取样测定。
1.2 热害指数的测定方法
参照赵大球[20]、罗少波[21]的方法并结合金莲花自身在高温条件下的受损情况划分等级:
1级:植株正常生长,新叶叶缘稍有反卷,色泽正常;
2级:少于1/5的植株叶片反卷,反卷部分轻度枯萎,能够正常的生长;
3级:1/5~2/5的植株叶片枯萎,部分植株生长受到影响;
4级:2/5~3/5的植株叶片枯萎,生长受到严重抑制;
5级:3/5以上的植株叶片枯萎,茎叶明显萎缩,生长完全停止。
热害指数=∑(各级株数×热害等级)/(最高级数×总株数)×100%
(1)
1.3 指标测定
1.3.1 生理指标测定
叶绿素含量和可溶性糖含量参照李合生等[22]的方法测定;相对电导率参照白宝璋等[23]的方法测定;MDA含量参照张志良等[24]的方法测定;游离脯氨酸(Pro)含量和超氧化物岐化酶活性(SOD)测定参照高俊凤等[25]的方法。
1.3.2 电阻抗参数测定
经不同浓度的SA处理后,每个处理随机选取8片叶片,剪成4 mm×10 mm矩形片段,测厚仪(Mitu-toyo No.7331,JaPan)测定其厚度(精确到0.01 mm)。参照RePo等[26]的方法,用电阻抗仪(HP4284A,USA)测定42个频率下的EIS参数,利用RePo发展的LEVM8·06(Macdonald JR)软件将等效电路参数采用单一DCE模型进行拟合,得到胞外电阻率(re)、胞内电阻率(ri)、弛豫时间(τ)和弛豫时间分布系数(ψ)。
1.4 数据处理
采用Microsoft Excel软件制图,SPSS 17.0软件对数据进行方差分析和相关性分析。
2 结果与分析
2.1 外源SA对高温胁迫下金莲花幼苗热害指数的影响
由表1可以看出,昼38℃/夜30℃高温胁迫下,各处理组的金莲花幼苗受到不同程度的高温伤害。随胁迫时间延长,CK组的热害级别逐渐增加,胁迫72 h后,叶片开始枯萎,严重抑制生长,热害级别达到4级,热害指数也显著高于各SA处理组(P<0.05),恢复生长4 d后热害等级仍为4级,受伤害程度没有改善。各SA处理组对缓解高温伤害都有不同程度的效果,胁迫72 h后,4个SA处理中,0.5和1.0 mmol·L-1SA处理的热害等级最低,热害指数显著低于对照(P<0.05),分别比对照低26.16%和50.15%,但0.5 mmol·L-1SA处理恢复生长4 d后热害级别没有改善,而1.0 mmol·L-1SA处理恢复生长后热害等级为最低,降到2级,由此可见,喷施1.0 mmol·L-1的SA减轻高温对金莲花幼苗伤害的效果最好。

表1 外源SA对高温胁迫下金莲花幼苗热害指数的影响
2.2 外源SA对高温胁迫下金莲花幼苗叶绿素总含量的影响
如图1所示,CK和不同浓度SA处理的金莲花幼苗叶绿素总含量随着高温胁迫时间的延长呈现出缓慢下降的趋势。经不同浓度(0.5~2.0 mmol·L-1)SA处理后的叶片叶绿素总含量均比CK高且差异显著(P<0.05)。高温胁迫72 h后,0.5、1.0、1.5、2.0 mmol·L-1SA处理的金莲花叶片中的叶绿素含量分别比对照高21.75%、27.59%、20.44%、20.58%,1.0 mmol·L-1SA处理的叶绿素含量显著高于CK和其他处理,并且在整个胁迫过程中,1.0 mmol·L-1处理的金莲花叶片叶绿素含量始终高于其他处理,说明此处理对金莲花幼苗叶绿素降解的缓解效应好于其他浓度。
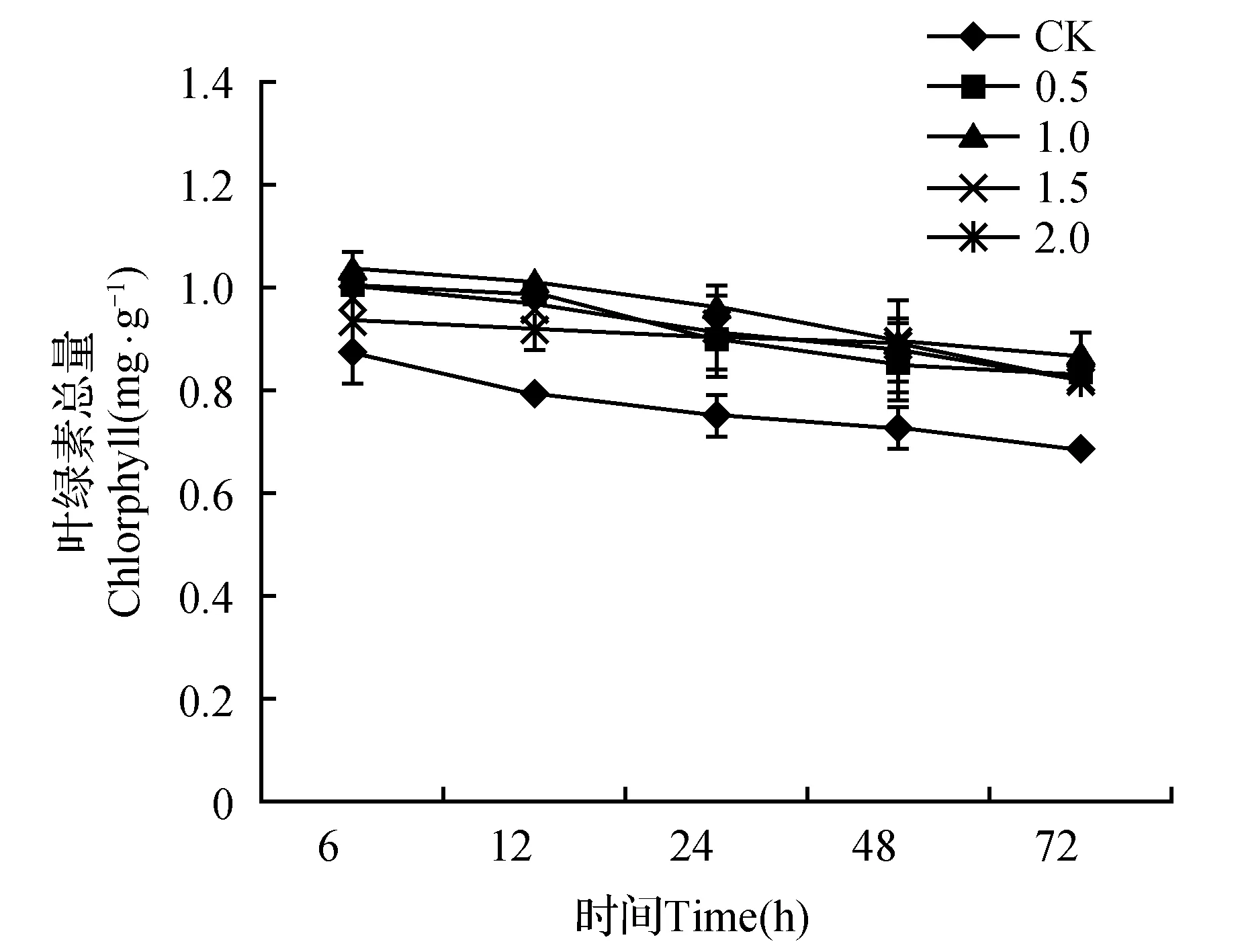
图1 外源SA对高温胁迫下金莲花幼苗叶绿素总含量的影响Fig.1 Effects of exogenous SA on total chlorophyll content of T.chinensis seedlings under high temperature stress
2.3 外源SA对高温胁迫下金莲花幼苗细胞膜稳定性的影响
由图2可以看出,高温胁迫条件下,CK和不同浓度SA处理金莲花幼苗的相对电导率均随处理时间的延长而升高。高温胁迫至72 h时,0.5和1.0 mmol·L-1SA处理均显著低于对照(P<0.05),两组处理分别比对照下降7.20%和9.70%。经1.5 mmol·L-1处理后电导率低于CK,但与CK组差异不显著,而经2.0 mmol·L-1处理后,其电导值显著高于CK(P<0.05)。由此可见,喷施0.5和1.0 mmol·L-1可以缓解高温胁迫,减轻高温对植物细胞膜结构的损伤。
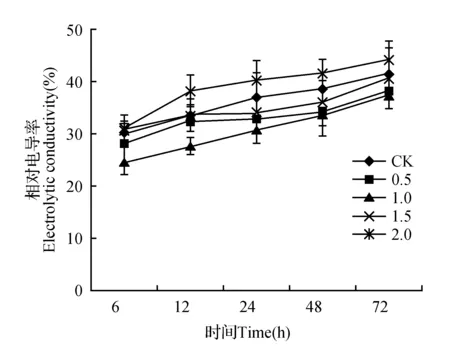
图2 外源SA对高温胁迫下金莲花幼苗相对电导率的影响Fig.2 Effects of exogenous SA on relative electric conductivity of T.chinensis seedlings under high temperature stress
如图3所示,随着高温胁迫的推进,CK和各SA浓度处理下金莲花幼苗叶片的MDA含量均逐渐上升,在所有处理中,CK的MDA含量随胁迫时间的延长上升的幅度最大,1.0 mmol·L-1SA处理组MDA含量上升的幅度最小。高温胁迫72 h后,0.5、1.0、1.5、2.0 mmol·L-1SA处理的金莲花叶片MDA含量分别比对照下降38.87%、57.26%、16.56%、34.56%,与对照差异显著(P<0.05),并且各SA处理组之间差异显著。1.0 mmol·L-1SA处理组在高温胁迫过程中MDA含量显著低于对照及其他处理(P<0.05)。由此可知,高温胁迫下,适宜浓度的SA能有效地减少MDA含量的积累,减轻高温对细胞膜脂过氧化的程度。
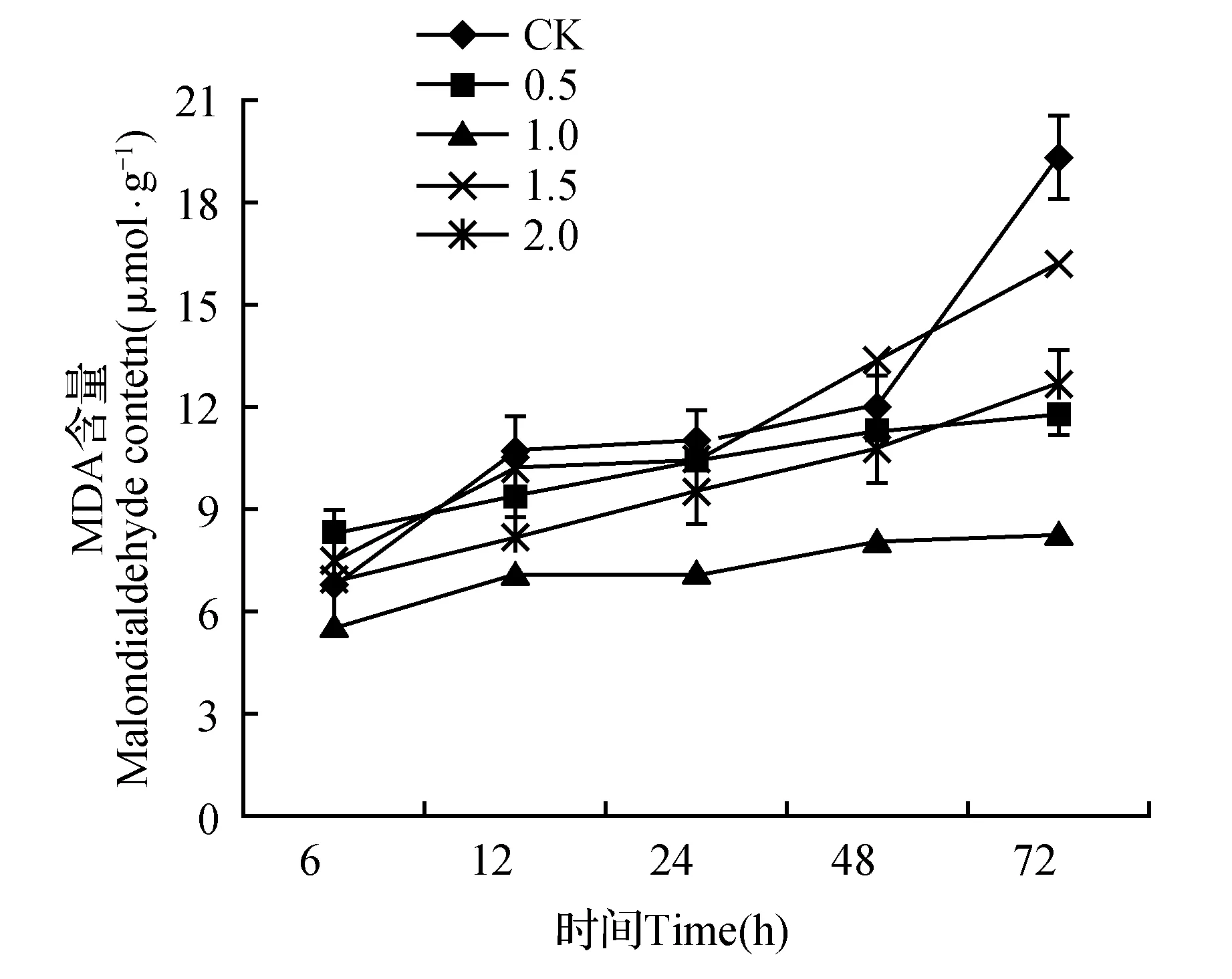
图3 外源SA对高温胁迫下金莲花幼苗MDA的影响Fig.3 Effects of exogenous SA on Malondialdehyder of T.chinensis seedlings under high temperature stress
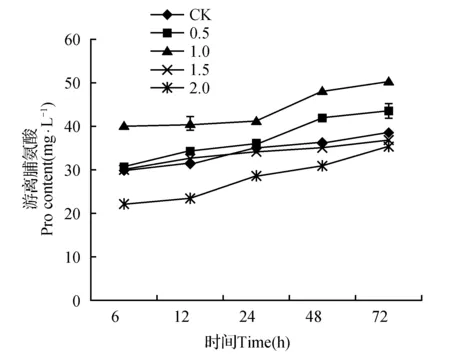
图4 外源SA对高温胁迫下金莲花幼苗游离脯氨酸的影响Fig.4 Effects of exogenous SA on Proline of T.chinensis seedlings under high temperature stress
2.4 外源SA对高温胁迫下金莲花幼苗渗透调节物质的影响
如图4所示,随着高温胁迫时间的延长,各处理的金莲花叶片Pro含量均有不同程度的涨幅。在同一处理时间,随SA浓度的增加,Pro量先增加后减少,并在1.0 mmol·L-1SA处理达到峰值。高温胁迫72 h后,1.0 mmol·L-1SA处理的金莲花叶片Pro含量显著高于对照及其它处理(P<0.05),比CK提高30.48%。0.5 mmol·L-1SA处理的Pro含量显著高于CK(P<0.05),比CK提高12.47%。SA浓度为1.5~2.0 mmol·L-1时,Pro含量与CK无显著差异。由此可见,高温胁迫下,1.0 mmol·L-1SA处理可以显著提高金莲花叶片中Pro含量。
由图5可以看出,各处理的金莲花叶片可溶性糖含量在6~72 h高温处理过程中逐渐增加。在同一胁迫时间,可溶性糖含量随SA浓度的增加呈现先升高后降低的趋势,SA浓度为1.0 mmol·L-1时可溶性糖含量达到峰值。高温胁迫72 h后,0.5~2.0 mmol·L-1SA处理均显著高于对照(P<0.05),可溶性糖含量分别比对照提高27.40%、49.66%、33.56%、20.89%。其中,1.0 mmol·L-1SA处理显著高于对照及其他处理(P<0.05),因此,叶面喷施1.0 mmol·L-1SA能使金莲花幼苗叶片的可溶性糖含量保持较高的水平,调节植物组织渗透势平衡,有效减轻细胞膜的损伤。

图5 外源SA对高温胁迫下金莲花幼苗可溶性糖含量的影响Fig.5 Effects of exogenous SA on soluble sugar of T.chinensis seedlings under high temperature stress

图6 外源SA对高温胁迫下金莲花幼苗超氧化物歧化酶活性的影响Fig.6 Effects of exogenous SA on superoxide dismutase of T.chinensis seedlings under high temperature stress
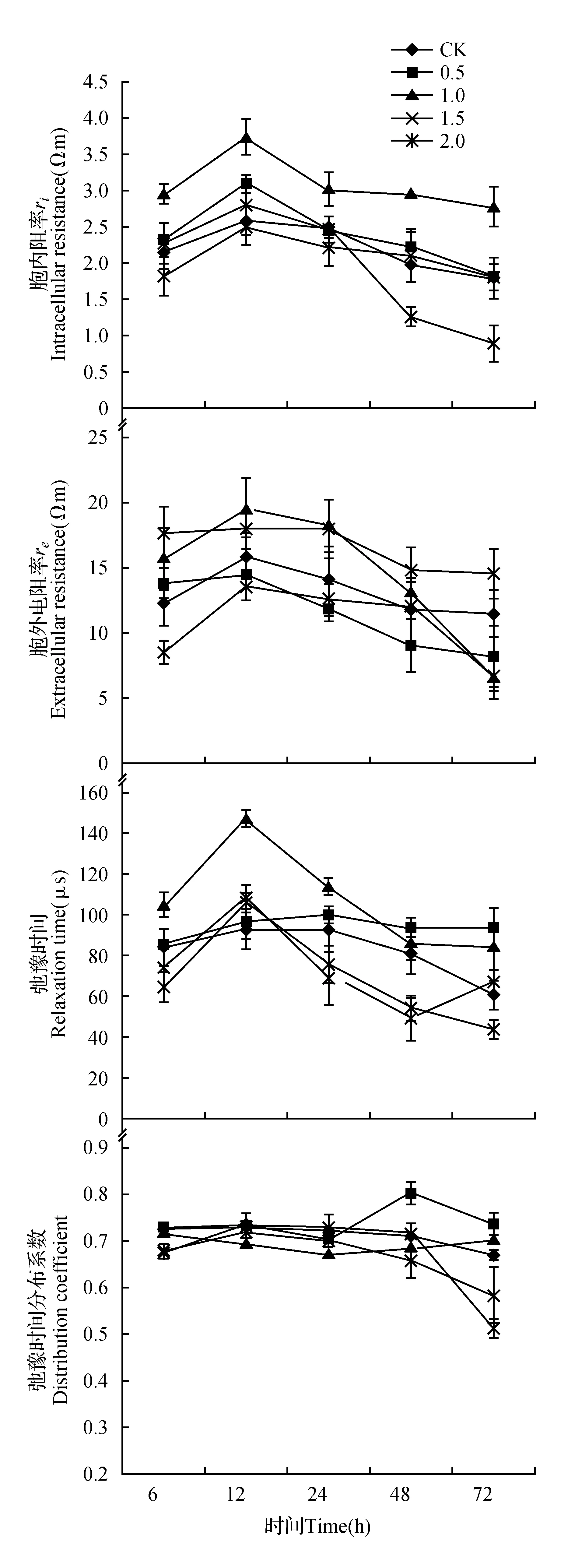
图7 外源SA对高温胁迫下金莲花幼苗电阻抗参数的影响Fig.7 Effects of exogenous SA on electrical impedance parameters of T.chinensis seedlings under high temperature stress
2.5 外源SA对高温胁迫下金莲花幼苗超氧化物歧化酶(SOD)活性的影响
如图6所示,各处理组金莲花幼苗叶片的SOD活性随高温胁迫时间延长均呈现出先上升后下降的趋势,并在胁迫24 h时达到峰值。同一胁迫时间内,SOD活性随SA浓度的增加先升高后降低,1.0 mmol·L-1SA处理SOD活性最高。在高温胁迫72 h时,1.0 mmol·L-1SA处理SOD活性显著高于对照及其他处理组(P<0.05),比对照高22.31%。经0.5、1.5 mmol·L-1SA处理后,SOD活性与对照差异不显著。当SA浓度为2.0 mmol·L-1时,SOD活性显著低于CK(P<0.05)。由此可见,经适宜SA浓度处理后,可使金莲花幼苗SOD活性增强,而浓度过高时幼苗SOD活性明显降低,说明高浓度SA会抑制SOD活性。
2.6 外源SA对高温胁迫下金莲花幼苗电阻抗参数的影响
EIS参数可以反映植物组织细胞的物理化学变化[27],如图7所示,各处理组的金莲花在高温胁迫过程中,胞外电阻率(re)、胞内电阻率(ri)和弛豫时间(τ)等电阻抗参数的变化趋势均呈现出先增大后减小,而不同处理的弛豫时间分布系数(ψ)变化趋势各不相同。
在6~12 h的高温胁迫下,各处理胞内电阻率(ri)和胞外电阻率(re)均呈现上升趋势,热胁迫12 h后,各处理胞内电阻率(ri)和胞外电阻率(re)均呈现下降趋势。高温胁迫72 h后,0.5、1.0 mmol·L-1SA处理的胞内电阻率(ri)值高于CK但无显著差异,1.5、2.0 mmol·L-1SA处理的胞内电阻率(ri)值低于CK,1.5 mmol·L-1SA处理与CK差异不显著,2.0 mmol·L-1SA处理显著低于CK(P<0.05)。高温胁迫至72 h后,0.5、1.0、2.0 mmol·L-1SA处理的胞外电阻率(re)均低于CK且达到显著性差异(P<0.05),而1.5 mmol·L-1SA处理后的胞外电阻率(re)显著高于CK和其他处理(P<0.05)。
在整个胁迫过程中,各处理的弛豫时间均呈现出先升高后降低的趋势,CK、1.0、1.5、2.0 mmol·L-1SA处理均在胁迫12 h时达到峰值,0.5 mmol·L-1SA处理在24 h达到峰值。高温胁迫至72 h后,经0.5、1.0 mmol·L-1SA处理后的弛豫时间(τ)均显著高于CK和其他处理(P<0.05),两者差异不显著。1.5 mmol·L-1SA处理的弛豫时间(τ)显著低于CK(P<0.05),2.0 mmol·L-1SA处理与CK差异不显著。
各处理的弛豫时间分布系数(ψ)变化趋势不同,CK、1.5和2.0 mmol·L-1SA处理随时间延长呈先上升后下降的趋势,0.5、1.0 mmol·L-1SA处理则呈先下降后上升的趋势。高温胁迫72 h后,0.5、1.0 mmol·L-1SA处理的弛豫时间分布系数(ψ)与CK无显著差异,1.5、2.0 mmol·L-1SA处理的弛豫时间分布系数(ψ)低于CK和其他处理且达到显著水平(P<0.05),两者无显著差异。
2.7 SA处理下金莲花幼苗各指标的相关性分析
由表3可知,热害指数与相对电导率呈极显著正相关(P<0.01),与叶绿素总量、可溶性糖含量和胞外电阻率(re)呈显著负相关(P<0.05)。胞内电阻率(ri)与Pro含量呈正相关,且相关系数达到显著水平(P<0.05)。弛豫时间(τ)与Pro含量和SOD活性呈极显著正相关(P<0.01)。相对电导率与Pro含量和SOD活性呈显著负相关(P<0.05)。

表3 SA处理下各指标相关系数
注:*在0.05水平上显著相关;**在0.01水平上极显著相关
Note:*significantly correlated at the level of 0.05;**significantly correlated at the level of 0.01
3 讨论
高温胁迫下,热害指数是评价植物对高温逆境的抗性强弱和反映植物受热伤害程度最直接的指标[28~30]。本研究结果表明,喷施适宜浓度的SA能够降低高温胁迫下金莲花幼苗的热害指数,提高其耐热性,缓解高温伤害。这与曹淑红[31]等对百日草耐热性、万正林[32]等对番茄抗高温效果的研究结果一致。叶绿素含量是对高温胁迫反应的敏感性生理指标[33],本研究中表明热害指数与叶绿素含量呈显著负相关,随着胁迫时间延长各处理的叶绿素含量呈明显下降趋势,但喷施适宜浓度的SA显著减缓了叶绿素的降解,在一定程度上反映了耐热能力的提高。
高温胁迫下,细胞膜系统是受到热伤害的首要部位,细胞膜透性和MDA含量是反映细胞质膜损伤程度的重要指标[34]。本研究中相关性分析表明热害指数与相对电导率呈显著正相关,喷施适宜浓度(0.5和1.0 mmol·L-1)的SA处理后相对电导率和MDA含量显著低于CK,这与李同根[35]等对皖贝母,张月雅[36]等对东北对开蕨的试验结论相似,说明喷施适宜浓度的SA能够有效缓解高温胁迫对细胞质膜的伤害。另外,游离脯氨酸和可溶性糖这些重要的渗透调节物质会在植物遭受胁迫时及时做出响应,调节渗透势来抵抗逆境,SOD活性的提高有利于防止膜脂过氧化,维持细胞膜稳定性[37],本研究也表明Pro含量和SOD活性与相对电导率呈显著负相关,可溶性糖含量与热害指数呈显著负相关,喷施适宜浓度(1.0 mmol·L-1)的SA能提高金莲花幼苗叶片的Pro和可溶性糖含量,增加SOD活性,孙军利[38]、杨岚[39]等研究水杨酸对高温胁迫下葡萄、铁皮石斛耐热性影响的也有类似的结果,这说明外源的SA可能通过调节金莲花幼苗叶片的SOD活性、Pro和可溶性糖含量等指标影响细胞膜的稳定性,从而提高金莲花幼苗抵御高温的能力。
植物能够自我调节来适应或抵抗逆境造成的伤害,利用EIS参数可以记录植物细胞电阻的变化,测定植物细胞在胁迫下的生理变化[40],这种测定方法更加高效便捷。本试验结果表明,弛豫时间(τ)与Pro含量和SOD活性呈极显著正相关,高温胁迫下,1.0 mmol·L-1SA处理金莲花幼苗胞外电阻率(re)、胞内电阻率(ri)及弛豫时间(τ)均高于对照,可能是喷施适宜浓度的水杨酸能有效降低细胞质膜透性,减缓膜脂过氧化,提高渗透调节物质含量,从而显著增加胞内胞外黏稠度,增强耐热性,这与相关性分析热害指数与re呈显著负相关,ri与Pro含量呈显著正相关的结果一致。因此,re和ri等电阻抗参数可作为金莲花对高温逆境反应的评价指标,通过测定EIS参数可以了解高温胁迫对金莲花幼苗组织膜透性的影响和热害程度。
综上所述,外源水杨酸能在高温环境中有效地降低金莲花幼苗的热害等级和热害指数,降低细胞质膜透性,减缓膜脂过氧化,缓解叶绿素降解,提高渗透调节物质含量和SOD活性,从而提高金莲花的耐热性,缓解高温伤害。电阻抗参数能有效地表示金莲花对高温的反应,可作为快速估测高温胁迫下金莲花幼苗组织内部生理变化的一种方法,从而检测高温伤害。在栽培生产中,夏季高温环境下对金莲花幼苗叶片适宜喷施1.0 mmol·L-1的SA,以缓解高温对金莲花幼苗的伤害。
