秀丽隐杆线虫模型在食品安全研究中的应用
2020-07-08夏程程钟晓凌张智源史文博宁吴柳志杰周梦舟
夏程程 杨 番 钟晓凌 李 琴 李 茜 张智源 史文博 徐 宁吴 茜 胡 勇 柳志杰 汪 超 周梦舟*
(1 湖北工业大学 工业发酵湖北省协同创新中心 湖北省食品发酵工程技术研究中心 武汉430068 2 武汉市环境检测中心 武汉430015 3 湖北省阿克瑞德检验检测有限公司 武汉430077)
随着人们生活水平的日益提高, 简单的温饱已不能满足绝大多数人的需求, 人们现在更关心食品的营养及安全问题。 由于食品安全关系人民群众的身体健康、生命安全及社会稳定,因此近年来都成为研究热点。由于生物进化的保守性,科学研究中常用模式生物来研究一些生命现象并得到一些信息和结论,从而类推到其它生物。常见的模式生物有果蝇、斑马鱼、小鼠等生物。然而,哺乳动物存在伦理性、试验周期过长和操作复杂等问题。因秀丽隐杆线虫(Caenorhabditis elegans)具有培育时间短、理化性质明显、遗传信息清楚、信号通路高度保守以及个体化差异小等优势, 而作为一种理想的模式生物被广泛应用在生命科学领域。在食品安全方面, 它可作为一种有效的手段去检测食品中的有毒物质。 线虫在食品安全领域主要作用归纳为:(1)环境毒理学评价;(2)研究食品危害源的毒理机制;(3)研究抑制食品危害的物质的保护机制以及对其的筛选。 随着包括RNAi 在内的全基因组筛选以及蛋白质组学等高通量筛选技术的发展, 可利用秀丽隐杆线虫进行毒性影响下体内生物活性分子筛选, 使得生物标志物体系的建立成为可能[1-4],相关技术还被应用在建立保护线虫抵抗病原菌的益生菌快速筛选模型等方面。现对秀丽隐杆线虫在食品安全领域中的应用做如下综述。
1 利用秀丽隐杆线虫作为模式生物的优势
秀丽隐杆线虫是一种在世界各地土壤中自由生长的小型线虫, 是主要以细菌为食的一种假体腔动物。 有典型线虫的基本结构,有口、咽、肠、性腺还有肌肉和各种系统,如神经系统、消化系统、排泄系统、生殖系统等。 线虫的寿命为2~3 周,在实验室环境温度在20 ℃左右时,线虫发育一代仅需3 d。 只需显微镜和铂丝挑虫器即可操作,用保存液和甘油混合放在-80 ℃冷冻保存,活化时将冷冻的线虫倒在有食物的NGM 培养基上即可。操作简便, 成本低以及试验周期短是秀丽隐杆线虫作为模式生物的主要优势。
秀丽隐杆线虫作为模式生物的另一大优势就是它的部分基因以及信号通路与人类的同源性高[5]。Sulston 绘制了世界上第一幅线虫的细胞谱系图,还发现了细胞凋亡途径, 并通过遗传突变和基因克隆手段克隆了十几个控制细胞凋亡的基因[6-8]。同期对人及哺乳动物细胞的研究发现, 秀丽隐杆线虫的细胞凋亡基因与人及哺乳动物的基因有高度的相似性, 所以线虫和人的细胞凋亡的控制机制和免疫机制是高度保守的。 线虫是第一个完成全基因测序的动物,在线虫的20 000 多个基因之中,有40%的基因和人类基因具有同源性。同时秀丽隐杆线虫还具有完备的发育和解剖学的特征以及复杂的遗传学特征。 秀丽隐杆线虫包含多种经典的神经递质如乙酰胆碱、 多巴胺、5-羟色胺、谷氨酸、γ-氨基丁酸和神经肽等。这些神经递质在神经元中的合成、 储存和代谢等过程都与哺乳动物具有高度相似性[9]。线虫的大多数离子通道基因同样也与哺乳动物具有同源性[10]。
在食品安全研究领域中, 往往涉及到线虫的免疫调控相关的信号通路, 至少有3 个相对独立的信号传导调控路径在线虫的免疫调控和生长发育包括死亡中发挥重要作用[11-15]。线虫的这种免疫机制从线虫到哺乳动物再到人类都是高度保守的, 因为高等哺乳动物都是由简单的多细胞生物进化而来, 同时在进化过程中会增加新的附加作用,最后逐渐发育成完整的免疫系统[16]。 表1 列出了秀丽隐杆线虫中与免疫调控相关的主要信号通路,并且大多数都在人体中有同源基因。

表1 秀丽隐杆线虫中与人类同源的部分信号通路Table 1 Partial signaling pathways homologous to humans in C. elegans
1.1 DAF-2/DAF-16 信号途径
DAF-2/DAF-16 信号途径又称胰岛素信号通路。 胰岛素信号是通过Forkhead 家族的转录因子DAF-16 蛋白发挥着核心调控功能[26]。DAF-2 作为胰岛素受体同源物定位于细胞膜上, 它与胞外类似胰岛素配体结合后,导致PI3K(磷脂酰肌醇激酶)催化亚基同源物AGE-1 活化,AGE-1 产生第二信使进一步激活下游AKT/PKB 同源物丝氨酸/苏氨酸激酶AKT-1、AKT-2, 磷酸化DAF-16,从而抑制DAF-16 入核发挥转录调节作用。对DAF-2 突变研究发现, 许多抗菌基因LYS、SPP-1 等表达上调[27],不过对胰岛素通路的下游基因的研究表明DAF-2 和AGE-1 的免疫调控可能参与了其它通路[20]。 研究表明,DAF-16 通路并不是线虫免疫的关键通路,而是线虫的免疫防御机制,是免疫和繁殖中的联系通路。哺乳动物PI3K 的直源同系基因PDK-1 和胰岛素/IGF-1 受体InR 这两个基因在胰岛素/IGF-1 信号传导中起着关键作用,并且其它几种途径组分已经显示出可以调节果蝇、小鼠以及人的衰老, 这意味着该途径对衰老的影响是保守的[28-29]。
1.2 MAPK 信号通路
MAPK(促分裂原活化蛋白激酶)包含有3 条信号传导途径:分别是P38MAPK(细胞因子抑制的抗炎药物结合蛋白激酶)、ERK-MAPK (细胞外信号调节激酶)和JNK-MAPK(氨基末端激酶),这些途径从酵母到哺乳动物均高度保守。 这3 条途径分别对应于秀丽隐杆线虫的PMK-1、MPK-1[30]和KGB-1 信号途径,他们控制着线虫应对外界刺激和压力、 细胞增殖及凋亡等重要的生理活动[31-33]。
P38MAPK 通路为秀丽隐杆线虫免疫应答的关键组分,研究表明P38MAPK 通路是通过NSY-1→SEK-1→PMK-1 级联反应介导线虫的免疫功能,其中PMK-1 在抗病原菌的免疫应激中发挥重要作用[34]。 JNK 信号通路被认为是线虫应对外界各种应激诱导的信号转导的关键分子, 参与对温度、氧气和渗透压等变化的非生物应激作用。ERK同样是MAPK 家族的一员, 它的信号途径是涉及调节细胞生长、发育及分裂的信号网络的核心,遵循MAPKs 的三级酶促级联反应,Ras 作为其上游激活蛋白,Raf 作 为MAPKKK,MAPK/ERK激酶(MEK)作为MAPKK,ERK 即MAPK,即Ras-Raf-MEK-ERK 途径。 但是对上游蛋白及各种激酶的激活机制正在进一步研究中。
研究表明这3 种MAPK 途径之间可相互作用,例如Kim 等[35]发现JNK 中的MEK-1 的MAPK激酶(MAPKK)和VHP-1 蛋白激酶磷酸酶(MKP)对线虫生物应激也有影响,并且也能调节PMK-1基因。 JNK 和P38MAPK 通路都是通过调节TTM-1 和TMM-2 基因抵御毒素[36]。P38 和ERK 信号通路间的双向信号能协同促进组织的修复,JNK 信号通路和ERK 信号通路存在负的相互作用[37-38]。
1.3 TGF-β 信号通路
TGF-β(转化生长因子)信号通路在成熟有机体和发育中的胚胎都参加了细胞生长、细胞分化、细胞凋亡和细胞动态平衡等其它细胞功能。 线虫中TGF-β 的同源基因为DBL-1。TGF-β 信号通路主要是由DBL-1 编码的蛋白和AMA/DAF-4 受体复合物结合磷酸化后激活下游SAM 家族蛋白来调控的[39]。 TGF-β 家族的其它成员包括TIG-2、UNC-129、DAF-7, 其功能多与神经和发育有关。线虫中对TGF-β 的免疫调控研究有助于我们了解动物中的致病菌以及对致病菌的抵抗机制[40-42]。
2 食品中安全危害因素以及秀丽隐杆线虫在相关研究中的应用
2.1 食品中化学物质危害因素
食品中化学危害包括食品包装材料、重金属、天然毒素和农药残留等,广泛存在于日常食品中,当剂量超过一定值,就可能导致食品安全问题。食品中的化学危害一直以来都是研究热点。
2.1.1 食品包装材料 世界上广泛使用的增塑剂邻苯二甲酸二(2-乙基己基)酯(DEHP)与其替代物邻苯二甲酸二乙酯(DEP)被应用于玩具、食品包装材料、医用血袋和胶管、乙烯地板和壁纸、清洁剂、润滑油以及个人护理用品(如指甲油、头发喷雾剂、香皂和洗发液)等数百种产品中,有研究表明[43],DEPH 对无脊椎动物有一定的急性毒性。Tseng 等[44]使用秀丽隐杆线虫为体内模型来评估邻苯二甲酸酯诱导的神经毒性和可能的相关机制, 通过抗氧化剂坏血酸预处理显著降低了DEHP 暴露线虫的ROS 水平, 改善了运动和热敏行为缺陷,以及保护了神经元的损伤,表明邻苯二甲酸酯对线虫的神经毒性作用中氧化应激起关键作用。 Pradhan 等[45]利用秀丽隐杆线虫研究了DEHP 和DEP 毒性以及致毒机理。 基因表达分析显示,在线虫L1 至青年期阶段期间DEHP 和DEP暴露改变了参与脂质代谢和应激反应的基因表达情况。 与脂质代谢有关的基因, 包括FASN-1,POD-2,FAT-5,ACS-6 和SBP-1 以及卵黄蛋白原上调, 应激反应有关基因中,CED-1,WAH-1,DAF-21 和GST-4 上调,而CTL-1,CDF-2 和热休克蛋白(HSP-16.1,HSP-16.48 和SIP-1)下调。 脂质染色显示在DEHP 暴露下, 线虫的脂质含量显著增加,生殖能力和寿命下降。
自从双酚A(BPA)被禁止应用在所有食品包装材料中[46]以来,与BPA 结构高度相似的双酚S作为替代品广泛应用于各种食品包装材料中。 但是BPS 同样也有一定的危害,CHEN 等[47]发现BPS 和BPA 能引起秀丽隐杆线虫生殖系统损伤,增加生殖细胞的凋亡,导致胚胎死亡率上升,但是和BPA 的作用途径不完全相同。低浓度的BPS 可能通过促进ROS 的过度产生加速线虫衰老,从而显著缩短线虫寿命, 因此得出结论双酚S 并不能很好替代双酚A。
2.1.2 重金属 随着工业科技的发展, 重金属对食品安全的影响越来越大, 自然界中一般广泛存在有一定浓度的重金属。 但是随着人类开采重金属,随意排放污水等活动,很多重金属进入了生态系统,经过时间积累,以及食物链的富集,最终在生物体内聚集,通过氧化作用,破坏生物体正常生理代谢活动,最后给人体健康造成巨大影响。Yang[48]发现0.4 μmol/L 以上的镉暴露可显著抑制细胞增殖,并具有剂量效应。
在20 世纪九十年代,Williams 和Dusenbery[49]首次研究了Ag,Hg,Cu,Cd 和As 等可以导致哺乳动物致死的14 种金属和非金属盐对线虫的毒性作用,测定了24,48,72 和96 h 的半致死剂量。 自此以后很多研究开始使用线虫作为模式生物进行重金属评估。 Boyd 等[50-51]通过对秀丽隐杆线虫的研究表明Cd,Hg 和Pb 暴露均会导致秀丽隐杆线虫数目显著下降。 WANG 等[52]发现重金属镉通过促进形成ROS(反应性氧类物质)和损害抗氧化系统、DNA 修复、RNA/蛋白质合成、细胞的分化和增殖来对生物体造成严重的威胁。 为了研究秀丽隐杆线虫在严重镉胁迫下的信号传导途径, 通过对照RNA-Seq 野生型和pmk-1 突变体在Cd 胁迫条件下进行的转录组分析,发现PMK-1 促进了镉胁迫动物中转录因子SKN-1 / Nrf 和DAF-16 的靶基因的表达, 其中包括分子伴侣蛋白或免疫蛋白的基因。
砷中毒所涉及的机制尚不明确, 研究认为砷可以改变多条细胞通路包括改变生长因子, 抑制细胞周期检查点蛋白和DNA 修复, 影响DNA 甲基化,削弱免疫监视,破坏氧化剂的平衡,增加氧化应激等[53]。
Anthony 等[54]通过秀丽隐杆线虫发现三价砷和亚砷酸盐可以诱导线粒体活性氧的产生, 抑制参与能量代谢的酶,并在体外诱导有氧糖酵解,提示砷可能通过影响代谢功能来诱导疾病的产生。此外, 亚砷酸盐暴露还会引起严重的线粒体功能障碍。 游牧[53]以秀丽隐杆线虫为模式生物,建立了基于秀丽隐杆线虫的抗砷胁迫模型。 试验发现热激处理可以显著提升线虫对砷的耐受度, 其机理是热激处理激活了胰岛素先天的免疫调节功能,与胰岛素途径关键位点HSF-1 和DAF-16 有关,热激处理促进DAF-16 核质转运的时间和比率,提高抗胁迫能力,并且具有显著的时间效应。通过对HSF-1 和DAF-16 基 因下游HSP-4、HSP-6、HSP-16.2、HSP-60 和HSP-70 5 种热休克蛋白的表达情况分析, 发现线虫对砷暴露抗性涉及内质网和线粒体的蛋白质折叠过程, 并可能与机体的氧化应激途径有关。 游牧[53]还利用秀丽隐杆线虫多种突变品系, 发现辣椒素通过胰岛素途径对砷胁迫造成的氧化应激损伤具有保护作用, 具体分子通路可能为: 辣椒素抑制胰岛素信号, 活化DAF-16/FOXO 和SKN-1, 两者独立平行发挥作用,共同提高抗氧化酶如GST-4、SOD-3 和GCS-1 的表达,提高线虫氧化还原能力。 通过对线虫这种模式生物的运用,建立了抗砷胁迫模型,探讨了胰岛素途径先天免疫调控功能对砷毒害的保护作用和分子机制。
研究者发现线虫的JNK 和P38 MAPK 信号通路都参与了对重金属毒素的抵御, 任何一条通路中的关键基因PMK-1、KGB-1 敲除都会使线虫对毒素极其敏感, 并且这两条通路都是通过调节TTM-1 和TTM-2 基因抵御重金属毒素。研究表明秀丽隐杆线虫MEK-1 基因在抗重金属方面发挥重要作用,MEK-1 信号途径MLK-1/MEK-1/KGB-1 介导对重金属铜、镉的抗性作用,在重金属的刺激下,KGB 表达上调[55]。同时SEK-1、MEK-1 对PMK-1 和KGB-1 的激活作用与VHP-1 对它们的抑制作用在细胞内达到平衡, 协调生物体内的稳定性[56]。
2.1.3 自然毒素 自然界中存在许多以食品的天然组分形式存在的自然毒素,由于毒性巨大,且与食品容易混淆,因此对人体的健康威胁巨大。自然毒素主要分为:有毒蛋白质类、有毒氨基酸类、生物碱类、蘑菇毒素、木藜芦烷毒素、毒甙和酚类衍生化物。
蓖麻毒素是一种核糖体失活蛋白(RIP),由能够每分钟失活1 500 个核糖体的异二聚体多肽组成,可中断蛋白质合成并导致细胞死亡。传统的蓖麻蛋白检测方法使用基于抗体的免疫测定和酶连接免疫吸附测定, 但是这些方法需要使用产生更多抗体的高等动物,但是这些动物一般都受保护,而且存在伦理问题。 Demant 等[57]利用秀丽隐杆线虫研究出一种可以量化蓖麻毒素的最佳生物测定方法,并选择出最佳测定毒素的线虫品系。
青颤毒素(Penitrem A, PA)是由几种陆地和少量海洋青霉菌物种产生的卤化吲哚生物碱毒素, 发现可以防止PA 毒性效应的天然产物对食品安全至关重要。 Goda 等[58]利用秀丽隐杆线虫和大鼠模型同时评估AST(虾青素)和DHA(二十二碳六烯酸)对PA 诱导的毒性的体外保护活性。 最后发现DHA 和AST 处理有效抵消了PA 的毒性作用并使大多数生化参数(谷氨酸、天冬氨酸、血清中多巴胺、血清素和肾上腺素)正常化。 并表示DHA 和AST 主要是通过抵消PA 诱导的BK 通道拮抗活性来抑制PA 对线虫的毒性,说明DHA 和AST 可以作为有用的食品添加剂来预防和减少PA 食品引起的毒性。
上述食品中化学危害因子涉及到的与线虫免疫防御相关的信号通路如图1 所示, 当食品的化学危害因子如食品包装材料中的有害物质以及重金属对秀丽隐杆线虫产生刺激时, 影响线虫中脂质代谢和应激反应的基因的表达,产生活性氧,降低了线虫的生殖能力以及寿命。 线虫中的JNK 通路和P38MAPK 通路都是通过调节TTM-1 和TTM-2 基因抵御毒素。KGB-1 为抗金属的关键基因, 同时SEK-1、MEK-1 对PMK-1 和KGB-1 的激活作用与VHP-1 对它们的抑制作用在细胞内达到平衡。而辣椒素可通过抑制胰岛素信号,活化DAF-16/FOXO 和SKN-1, 两者独立平行发挥作用, 共同提高抗氧化酶如GST-4、SOD-3、GCS-1的表达, 提高线虫氧化还原能力来抵抗重金属对线虫的影响。
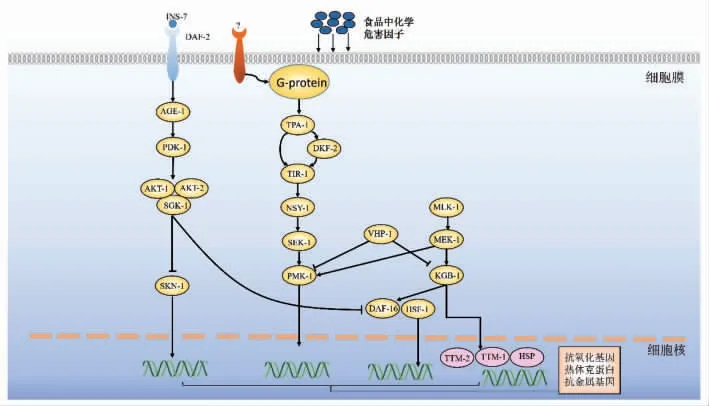
图1 秀丽隐杆线虫中与食品化学危害因子相关的部分免疫防御信号通路示意图Fig.1 Schematic diagram of some of the immune defense signaling pathways associated with chemical hazards in foods of Caenorhabditis elegans
2.2 食品中微生物
食品中的微生物危害包括细菌、病毒、寄生虫和真菌等方面的危害。 食品中的微生物危害涉及范围广,食品在加工、运输、贮藏和销售等过程中都可能被微生物及其毒素污染, 导致食品卫生质量下降,对人体健康产生危害。在研究食品微生物危害方面利用秀丽隐杆线虫作为模式生物的研究中,主要与真菌及致病性细菌有关。
2.2.1 真菌 真菌毒素是一类由真菌产生的有毒次级代谢产物, 通过污染食品进入人类和动物体内引发癌症以及不孕等症状。因其分布广泛,难以避免,故对人类的健康有很大的影响。真菌主要的共同毒性归结为细胞毒性和致DNA 损伤,而不同真菌又有其各自特点。 真菌毒素的毒理机制主要分为类雌性激素效应、氧化损伤、染色体畸变和诱发细胞凋亡。
黄曲霉毒素是由黄曲霉和曲霉菌产生的一种真菌毒素, 天然存在于各种动物饲料和人类食物中,特别是在谷物、坚果和饲料中易被发现[59]。 黄曲霉毒素B1(AFB1)是已知的真菌毒素中毒性最强和致癌性最强的物质。Leung 等[60]利用秀丽隐杆线虫研究黄曲霉毒素B1 对细胞色素酶P450 系列(CYP)的影响, 发现其会导致DNA 突变细胞衰亡。 这是公开报道秀丽隐杆线虫可用于真菌毒素毒性研究的第一篇文章, 自此之后秀丽隐杆线虫被广泛地应用于真菌毒素研究中。研究表明AFB1主要通过其代谢物和DNA 形成加合物阻断DNA的复制和转录,导致DNA 的损伤。除了致癌作用,AFBI 还会在人和哺乳动物中产生其它严重的毒性作用,比如生长延缓,免疫控制以及生殖功能障碍。 Eduardo 等[61]使用秀丽隐杆线虫作为体内模型,着重研究AFB1 对生长和繁殖以及DNA 损伤和生殖细胞凋亡的毒性作用。结果表明,将秀丽隐杆线虫暴露于AFB1 会引起浓度依赖性生长和繁殖抑制,核DNA 损伤和生殖细胞凋亡这些毒性效应之间存在着潜在的相互关系,而且DNA 损伤为中心枢纽, 通过关键信号通路导致生长抑制和生殖细胞凋亡,后者进一步抑制生殖。
脱氧雪腐镰刀菌烯醇(DON)是由某些感染禾谷类作物的镰孢属物种产生的次级代谢产物。DON 能引起拒食、呕吐、胃肠道病变、免疫抑制和家畜缺乏肌肉协调等症状, 又称呕吐毒素。 DON降低了作物产量以及影响了牲畜和人类健康。Gowrinathan 等[62]研究表明DON 可以抑制秀丽隐杆线虫野生型N2 和突变体品系AU1 后代数目,而且抑制作用与DON 暴露浓度有明显剂量依赖关系,染毒浓度越大,后代数目越少。 而且这种情况还存在于镰刀菌类真菌毒素中, 因此推测镰刀菌类真菌毒素对线虫存在生殖毒性。
Yang 等[63]选取秀丽隐杆线虫为模式生物,探索真菌毒素对秀丽隐杆线虫的毒性效应, 深入研究了以玉米赤霉烯酮(ZEN)为代表的真菌毒素的毒性效应与线虫代谢的相关性, 并分析其毒理机制。研究发现真菌毒素会抑制线虫的体型、寿命和后代数, 导致线虫性腺臂和产卵器缺陷以及行为能力缺陷。 微阵列结果显示过半基因受ZEN 调控表达有异常, 其中DAY-17、COL-121、HCH-7、NLP-9 显著差异表达, 分别与线虫表皮胶原蛋白合成、生殖细胞生产和免疫应激功能相关。ZEN 使DPY-31、DAF-2、DAF-16 标记基因在荧光线虫品系中强烈表达,显示了ZEN 对表皮胶原蛋白的下调作用和DAF-2insulin-like 信号通路的上调作用。 推断ZEN 的致死性、发育毒性、生殖毒性、神经毒性与线虫表皮胶原蛋白合成信号通路以及DAF-2insulin-like 信号通路相关。 该研究构建了一种有效评价真菌毒素毒性效应的秀丽隐杆线虫模型, 可快速筛选毒性评价指标并在分子水平分析基因表达与毒性效应相关性,阐明了毒理机制。
2.2.2 致病性细菌 食品致病菌是指可以引起食物中毒,或者以食品为传播媒介的致病性细菌,是食品安全问题的重要源头。 常见的食品致病菌有痢疾杆菌、鼠疫杆菌、致病性大肠杆菌、沙门氏菌、霍乱弧菌和猪丹毒杆菌等,对人类的健康有很大影响。
产肠毒性大肠杆菌(ETEC)是一类引起人和幼畜腹泻的主要病原菌。 Zhou[64]使用秀丽隐杆线虫建立了ETEC 致死线虫模型和保护线虫抵抗ETECCJG280 的乳酸菌快速筛选模型, 并挑出保护效果最好的一株玉米乳杆菌来研究益生菌保护线虫的机理。 发现LB1 对线虫的保护作用与抑制ETECJG280 肠毒素有关,并且发现活的LB1 能显著提高MAPK 和DAF-16 信号通路中关键基因及抗菌肽基因的表达水平,抑制肠毒素基因的表达,显著减少肠毒素克隆的致死作用。
Bolz 等[65]对感染鼠疫杆菌的秀丽隐杆线虫进行研究发现 PMK-1/P38MAPK 通路在宿主的先天免疫应答中发挥重要的作用。 肠道沙门氏菌感染秀丽隐杆线虫导致性腺细胞的程序性死亡,推断是菌体表面的LPS(lipopolysaccharide)和PMK-1 共同作用的结果。 RNAi 实验显示PMK-1 信号途径位于CED-9 上游[66]。 通过感染绿脓杆菌的秀丽隐杆线虫筛选出对病原菌敏感性增强(enhanced suscepttbility to pathogen,ESP)表型的突变株, 分析发现对应MAPK 途径中的SEK-1 和NSY-1 的基因ESP-2 和ESP-8 介导了秀丽隐杆线虫对病原菌的抗性作用[34]。 在铜绿假单胞菌感染线虫过程中,TPA-1 和DKF-2 (蛋白激酶D)激活PMK-1,诱导宿主防御基因的表达[67]。铜绿假单胞菌感染导致神经系统中的胰岛素样肽(INS-7)的产生,可激活DAF-2 并下调肠中DAF-16 的活性[68-69],胰岛素受体DAF-2 通过激酶级联抑制下游转录因子DAF-16, 该激酶级联包括磷酸肌醇3-激酶(AGE-1),磷酸肌醇依赖性激酶(PDK-1,AKT-1,AKT-2)和血清/糖皮质激素调节激酶1(SGK-1)[70]。 DAF-2 或AGE-1 中的突变体在喂食革兰氏阳性菌后,激活了DAF-16,延长了线虫的寿命,增强了线虫对死亡的抗性[71]。 粪肠球菌[72-73]通过产生活性氧簇介导对线虫的致死作用, 秀丽隐杆线虫通过DAF-2/DAF-16 信号途径与热休克因子HSF-1 之间的相互作用,调节CLT-1(过氧化氢酶)、CLT-2 和SOD-3(超氧化物歧化酶)等抗氧化基因的表达, 通过表达的这些酶分解活性氧簇[74-76]。
当粘质沙雷氏菌(Serratia marcescens)感染线虫时,溶菌酶基因(LYS-1、LYS-7 和LYS-8)以及凝集素基因表达均上调,且LYS-8 受DBL-1 正调控[77]。 Mallo 和Ewbank[78-79]的研究发现,当突变株dbl-1(DBL-1 基因被敲除)被铜绿假单胞菌(Pseudomonas aeruginosa)和粘质沙雷氏菌感染时,和野生型相比寿命明显减少。 Thomsen 等[80]利用秀丽隐杆线虫建立了李斯特菌的毒理分析模型。
秀丽隐杆线虫对致病菌的防御主要涉及前文提到过的线虫免疫相关的3 条信号通路以及一些抗菌肽基因, 而且各种信号通路之间有一定的相互作用,共同维持细胞内的平衡。 例如PMK-1 和DAF-16 都参与对病原菌的抵抗作用,PMK-1 的过量表达能增加DAF-2 突变引起的病原菌抗性作用[81]。另外,MAPK 和TGF-β 途径都参与抗绿脓杆菌的抗性作用[82]。 说明不同的信号途径间在抗致病菌方面可能通过相互作用, 整体上协调细胞免疫防御机制。
线虫的免疫系统以及抗微生物相关信号通路的信号通路图如下[83-84]:

图2 线虫的免疫系统以及抗微生物因子相关信号通路示意图Fig.2 Schematic representation of nematode immune system and antimicrobial signaling pathways
如图, 秀丽隐杆线虫自身的免疫防御机制主要如下:食品中微生物危害因子刺激下,TGF-β 通路的DBL-1 基因编码的蛋白与SMA/DAF-4 受体复合物结合磷酸化后激活下游SAM 家族蛋白,使得溶菌酶基因和凝集素基因均上调; 胰岛素信号通路上游神经样肽INS-7 激活DAF-2,抑制下游DAF-16 入核来发挥转录调控作用,上调了抗菌基因LYS、SPP-1 的表达并且和HSF-1 相互作用,调节抗氧化基因, 使其表达的蛋白分解活性氧簇。P38MAPK 通路主要是通过PMK-1 基因调控下游基因编码的神经样肽、 抗菌肽以及自噬基因的表达。 同时MAPK 中的JNK-1 信号途径中MEK-1对PMK-1 也有调控作用。
3 总结和展望
秀丽隐杆线虫因其具有结构简单、周身透明、生命周期短、 个体特异性小以及遗传模式保守等优点,已经被广泛应用于各个领域。在食品安全研究中,建立了许多线虫致死模型、线虫筛选模型和线虫抵抗模型。 研究了各种外界危害因子对线虫的毒性机理, 线虫自身的免疫防御的调控机制以及对有害物质起抑制作用的物质对线虫的保护作用机理, 充分说明了线虫作为模式生物在这个领域的重要性。不过,尽管线虫与人类基因具有高度保守性, 但是线虫和人类在生理以及病理方面差异较大,线虫不具备完整的循环系统、呼吸系统和获得性免疫系统, 所以它不完全能代替其它哺乳动物进行研究。由于线虫存活温度的限制,所以缩小了可研究的病原菌范围。此外,线虫强大的解毒系统,限制了一些调节宿主防御的化合物的发现。秀丽隐杆线虫模型在某些筛选模型中, 还需要后续的临床试验[85]。 同时我们应当正确认识秀丽隐杆线虫在食品安全研究中的优势和不足, 进行周密的试验方案设计, 才能充分发挥线虫模型的真正价值, 使秀丽隐杆线虫在更多领域发挥其不可或缺的作用。
秀丽隐杆线虫今后在食品领域的应用方向,可以借鉴保护线虫抵抗ETEC JG280 的乳酸菌快速筛选模型,利用线虫模型的快速筛选性,进行体内生物活性分子筛选, 也可以从食品中筛选出对人体有潜在危害的物质。 秀丽隐杆线虫这种模式生物的各种优势,在21 世纪,可能与生物信息学相结合,发挥更重要的作用。
