外源茉莉酸对稻瘟病菌引起褐点型坏死斑的水稻防御响应的影响
2020-07-07李春琴段桂花马笑晴刘文倩唐萍杨静
李春琴 段桂花 马笑晴 刘文倩 唐萍 杨静


摘要:【目的】解析外源茉莉酸(JA)對JA合成/响应、水杨酸(SA)合成/受体及防御相关基因表达的影响,为进一步揭示JA调控稻瘟病菌侵染引起褐点型坏死斑的水稻抗病作用机制提供基础数据。【方法】以接种稻瘟病菌菌株Y92-66b后形成褐点型坏死斑的地方中抗水稻月亮谷为研究材料,分别利用400 μmol/L JA和200 μmol/L JA抑制剂(IBU)外源处理接种稻瘟病菌后引起褐点型坏死斑(72 h)的水稻月亮谷,调查水稻稻瘟病症状,同时利用实时荧光定量PCR(qRT-PCR)检测JA合成/响应、SA合成/受体、防御和蔗糖/果糖合成酶基因的表达。【结果】外源JA有效减轻了水稻稻瘟病症状。同时,JA诱导水稻JA合成基因OsLOX1、OsOPR1、OsOPR7、OsJIMT、OsHPL3和OsAOS2在72 h时上调表达,至96和120 h时,只有OsAOS2、OsOPR7和OsLOX1上调表达,其余3个基因下调表达;外源JA未诱导OsLOX3表达;JA诱导水稻JA响应基因OsCOI1a、OsMYC2、OsJAZ1、OsJAZ9和OsbHLH35在72 h时上调表达,至120 h时,只有OsCOI1a和OsJAZ1上调表达,其余3个基因均下调表达;外源JA未诱导OsCOI1b和JiOsPR10表达。JA诱导SA受体基因OsNPR4在72 h时显著上调表达(P<0.05,下同),96 h时下调表达,至120 h时又上调表达但不显著(P>0.05)。JA诱导SA合成相关基因OsPAD4在72 h时显著上调表达,OsEDS1在120 h时显著上调表达。外源JA诱导防御基因OsPR5在72 h时开始上调表达,至96 h时显著上调表达;外源JA未诱导OsPR4a表达。外源JA诱导参与细胞壁合成的蔗糖合成酶基因OsRSUS1和果糖合成酶基因OsFRK-2表达,IBU则加重了水稻稻瘟病症状,同时抑制大部分JA合成/响应、蔗糖/果糖合成酶和防御基因的表达。【结论】外源JA主要通过诱导稻瘟菌引起的褐点型坏死斑的水稻JA合成/响应基因、参与细胞壁合成的蔗糖/果糖合成酶及防御基因表达参与水稻防御响应。
关键词: 水稻;稻瘟病菌;茉莉酸;防御响应;褐典型坏死斑
中图分类号: S435.111.41 文献标志码: A 文章编号:2095-1191(2020)05-1053-09
Abstract:【Objective】To analyze the effects of exogenous jasmonic acid(JA) on endogenous JA synthesis/response, salicylic acid(SA) synthesis/receptor and defense related gene expression were studied to provide reference for in-depth research on the JA regulation in rice defense mechanism during the formation of brown necrotic spots caused by Magnaporthe oryzae infection. 【Method】In this study, medium resistant rice cultivar Yuelianggu was inoculated with blast strain Y92-66b and treated with 400 μmol/L JA and 200 μmol/L JA depressor(IBU) during the formation of brown necrotic spots(72 h), the symptom of rice blast were investigated. And expression level of JA synthesis/response, SA synthesis/receptor, defense related and cell-wall synthesis relatedsucrose/fructose synthetase genes were detected by real-time fluorescence quantitative PCR(qRT-PCR). 【Result】Exogenous JA effectively reduced the symptoms of rice blast disease. Simultaneously, JA induced up-regulation of JA-synthesis genes of OsLOX1,OsOPR1,OsOPR7,OsJIMT,OsHPL3 and OsAOS2 at 72 h,only expression of OsAOS2,OsOPR7 and OsLOX1 were up-regulated at 96 and 120 h,and the other three genes were down-regulated. The expression of OsLOX3 was not induced by exogenous JA. Exogenous JA induced JA-response genes such as OsCOI1a,OsMYC2,OsJAZ1,OsJAZ9 and OsbHLH35 at 72 h,only OsCOI1a and OsJAZ1 were up-regulated at 120 h, and the other three genes were down-regulated. Exogenous JA did not induce expression of OsCOI1b and JiOsPR10. The SA receptor gene of OsNPR4 appeared significantly up-regulated at 72 h induced by exogenous JA(P<0.05, the same below), down-regulated at 96 h and up-regulated at 120 h but insignificantly(P>0.05). JA induced SA synthesis related gene OsPAD4 was significantly up-regulated at 72 h and OsSED1 was significantly up-regulated at 120 h.OsPR5, a defense gene induced by JA, was up-regulated at 72 h and significantly up-regulated at 96 h. Exogenous JA did not induce OsPR4a expression, but induced the expression of sucrose synthetase gene of OsRSUS1 and fructose synthetase gene of OsFRK-2, which were involved in cell wall synthesis. While IBU aggravated the symptoms of rice blast and inhibited expression of most JA synthesis/response, sucrose/fructose synthetase and defense genes. 【Conclusion】Exogenous JA mainly participate rice defense response by inducing expression of JA synthesis/response genes,sucrose/fructose synthetase and defense genes that are involved in cell wall synthesis during the formation of brown necrotic spots caused by M. oryzae infection.
Key words: rice; Magnaporthe oryzae; jasmonic acids; defense response; brown necrotic spots
Foundation item: National Natural Science Foundation of China(31860483,31400073); Program for Innovative Research Team in Science and Technology in Universities of Yunnan(2019)
0 引言
【研究意义】稻瘟病菌属于半活体寄生真菌,能侵染水稻植株的各部位引起葉瘟、穗颈瘟、谷粒瘟等,是造成水稻减产的主要病害之一,每年可造成10%~30%的水稻产量损失(Nalley et al.,2016;曹慧娟等,2019)。目前,稻瘟病防治主要通过选育抗病品种及化学防治,生产上使用的抗病品种大多抗谱较窄,因而在水稻抗病育种实践上的应用价值不高(Dangl et al.,2013),加之稻瘟病菌毒性小种变异速度较快,抗病品种种植3~5年后易被新的毒性小种克服,导致抗性丧失(Hasan et al.,2017);化学防治一直是生产上广泛使用的防治措施,但随之带来的环境污染及稻瘟病菌毒性小种耐药性增强等问题严重限制了化学防治的持久大规模应用,也是最不可取的防治方法(Gurjar et al.,2012; Law et al.,2017)。因此,非常有必要寻找更加安全有效的稻瘟病菌绿色防控方法。【前人研究进展】目前农业生产上以植物激素在内的许多激发子已逐渐应用于植物病害防治,如外源茉莉酸(Jasmonic acid,JA)喷雾处理大麦等植物后显著提高了植株抗性(Oka et al.,2013;Cao et al.,2016)。Shang等(2011)报道水杨酸(Salicylic acid,SA)预先喷施烟草后再喷施JA,能有效防控烟草病毒病害,是一种绿色安全有效的防控方法。JA处理水稻后诱导芳樟醇等醇类抗菌物质合成(Taniguchi et al.,2014)及许多PR基因如OsPR1a、OsPR1b、OsPR2、OsPR5和OsPR10等上调表达,激活水稻免疫响应(Agrawal et al.,2000a,2000b;王云锋等,2018)。因此,JA在植物病害防治方面具有极佳的应用潜力。稻瘟病菌孢子在稻瘟病菌侵染循环中起核心作用,当稻瘟病菌孢子接触水稻叶片表面(0 h)后开始发芽(2 h)并形成附着胞(8 h),附着胞穿透寄主角质层侵入细胞(16~20 h),随后侵染菌丝长满侵入的第一个细胞(20~36 h),并向临近细胞扩展定殖(36~48 h)(Koga,1994),属于稻瘟病菌活体营养阶段;72 h时水稻开始显现褐点型坏死斑,随后进入死体营养阶段(Bhadauria et al.,2013)。由于部分半活体寄生真菌在侵染早期无法检测,加之田间状态下观察不到任何症状(García-Guzmán and Heil,2014),以至于无法确定最佳的防控时期,与上述半活体寄生真菌有所区别的是,稻瘟病菌侵染72 h时的水稻叶片上开始显现肉眼可见的褐点型坏死斑,为生产上制定稻瘟病防控措施提供了非常重要的契机。据此,本课题组前期利用外源JA处理稻瘟病菌侵染72 h时的高感水稻品种丽江新团黑谷,发现外源JA激活了高感水稻防御相关基因及JA途径相关基因上调,有效控制了稻瘟病发生,表明外源JA处理稻瘟病菌侵染72 h时的高感水稻是稻瘟病防控的最佳时间(王云锋等,2018)。【本研究切入点】虽然前期研究已明确JA在高感水稻与稻瘟病菌强致病菌株互作体系中能有效控制稻瘟病的发生,但关于外源JA在抗病水稻与稻瘟病菌强致病菌株互作体系中对水稻抗瘟性及JA合成/响应和防御体系的影响尚未开展相关研究。【拟解决的关键问题】以外源JA和JA抑制剂IBU(Ibuprofen)处理接种稻瘟病菌后褐点型坏死斑形成时期(接种后72 h,下同)的地方抗病水稻月亮谷,解析外源JA对JA合成/响应、SA合成/受体、防御和蔗糖/果糖合成酶基因表达的影响,以明确JA对月亮谷抗瘟性及防御体系的影响,为进一步揭示JA调控稻瘟病菌侵染引起褐点型坏死斑的水稻抗病作用机制提供基础数据。
1 材料与方法
1. 1 试验材料
供试水稻品种为月亮谷(月亮谷是云南元阳县种植了上百年的传统地方中抗稻瘟病品种,本课题组早期研究发现该品种具有很多优良的农艺性状基因),供试稻瘟病菌株为Y92-66b(强致病性菌株),均由云南生物资源保护与利用国家重点实验室保存提供。
1. 2 试验方法
1. 2. 1 水稻育苗及稻瘟病菌产孢培养 播种前将水稻土和腐殖土按2∶1混合均匀,灭菌后装于育苗盘中备用。水稻种子先用1.5%次氯酸钠消毒5 min,清水冲洗3~5遍,置于28 ℃恒温培养箱中催芽至露白后播种于育秧盘中。水稻长至三叶一心期(约21 d)时用于稻瘟病菌接种试验。
配制PDA培养基(马铃薯200 g、葡萄糖20 g、琼脂粉15 g、蒸馏水1000 mL)用于稻瘟病菌株活化。挑取保存的Y92-66b菌种于PDA固体培养基上,置于28 ℃恒温培养箱中活化培养,将活化菌丝移至PDA液体培养基中培养4~5 d有大量菌丝产生。将菌丝涂布于西梅汁培养基(西梅汁40 mL、酵母提取物1 g、乳糖5 g、琼脂粉15 g、蒸馏水定容至1000 mL,pH 6.5)上,置于28 ℃光照培养箱(12 h黑暗和12 h光照交替)中培养至培养基表面长满菌丝,刮去培养基表面的菌丝后再持续光照培养5 d,即可产生大量分生孢子。
1. 2. 2 稻瘟病菌接种及JA和IBU处理接种稻瘟病菌后引起褐点型坏死斑的水稻 无菌水制备孢子悬浮液,显微镜下调节孢子浓度为1×105个/mL。孢子悬浮液喷雾于三叶一心期的水稻苗,将接种后的水稻苗置于培养箱中黑暗保温(28 ℃)保湿(95%以上)24 h。于喷雾稻瘟病菌接种后0、24、48、72、96和120 h分别取样,每个样品进行3次生物学重复,每次重复6株水稻苗。
JA浓度配制按王云锋等(2018)的方法,JA溶于100%乙醇中配制成300 mmol/L的母液,再用灭菌ddH2O稀释为400 ?mol/L的工作液。IBU溶于100%乙醇配制成300 mmol/L的母液,再用灭菌ddH2O稀释为100、200和300 ?mol/L的工作液。为确定IBU的浓度,利用100、200、300和400 ?mol/L的IBU分别处理接种稻瘟菌72 h的水稻,以无菌水处理接种稻瘟菌72 h的水稻为对照(CK),观察水稻发病症状和调查病情指数。将400 ?mol/L JA和200 ?mol/L IBU分别喷雾处理接种稻瘟病菌引起褐点型坏死斑的水稻,于JA和IBU喷雾处理后72、96和120 h分别取样。每个样品进行3次生物学重复,每次重复6株水稻苗。于接种7 d时进行病害调查,病级分级標准参照许志刚(2002)的方法。
病情指数=∑(各级病叶数×各级代表值)/(调查
总叶数×最高一级代表值)×100
1. 2. 3 水稻JA合成/响应、SA合成/受体及防御相关基因表达分析 水稻基因表达分析参照王云锋等(2018)的方法,并稍作修改。采用TransZol UP(北京全式金生物技术有限公司)试剂盒提取水稻总RNA,利用反转录试剂盒TransScript All-in-One First-Strand cDNA Synthesis SuperMix for qPCR(One-Step gDNA Removal)-AT341(北京全式金生物技术有限公司)将RNA反转录合成cDNA。根据JA合成/响应、SA合成/受体及防御相关基因的核酸序列设计引物,引物名称及序列见表1。采用TransStart Top Green qPCR SuperMix-AQ131(北京全式金生物技术有限公司)试剂盒进行实时荧光定量PCR(qRT-PCR),反应体系20 ?L:上、下游引物各1 ?L,cDNA模板1 ?L,荧光染料10 ?L,ddH2O补足至20 ?L;扩增程序:95 ℃预变性3 min;95 ℃ 20 s,60 ℃ 20 s,进行45个循环。60 ℃升高到98 ℃获取溶解曲线。试验重复3次。
1. 3 统计分析
所有数据采用IBM SPSS Modeler 17.0进行处理,运用SigmaPlot 10.0作图。
2 结果与分析
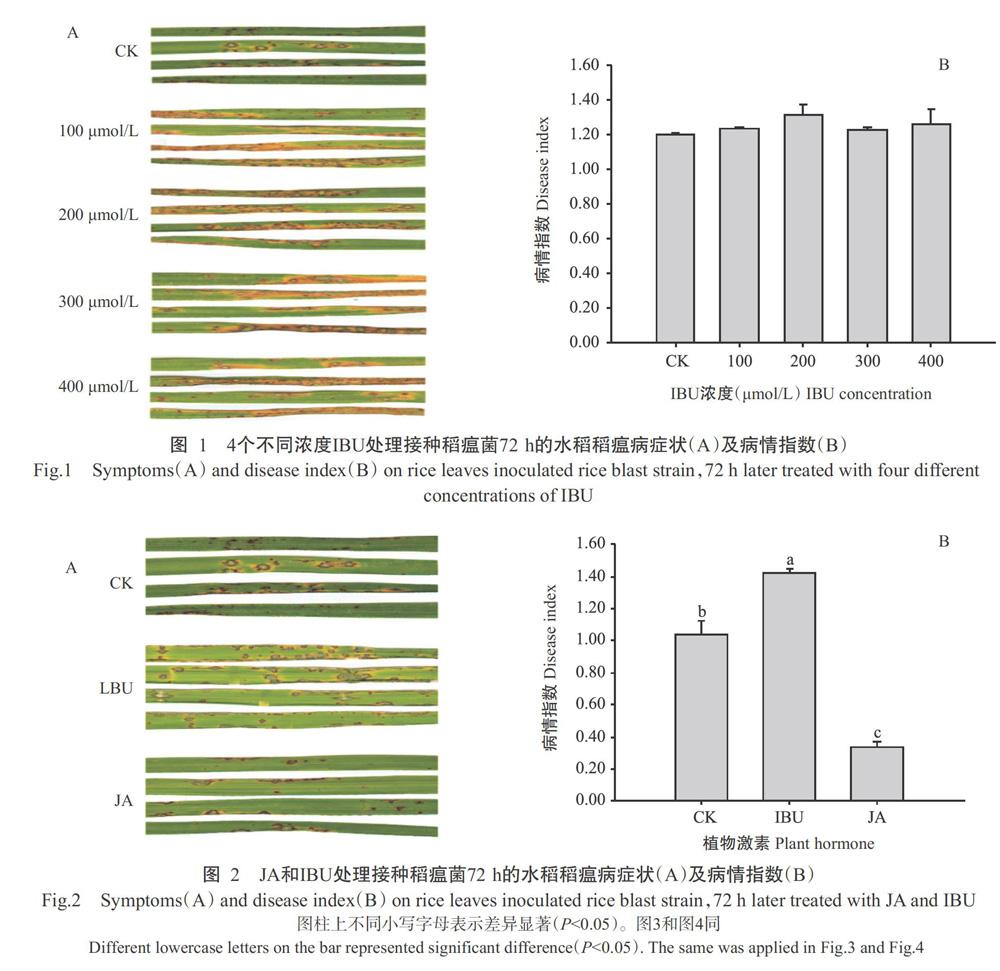
2. 1 不同浓度IBU处理接种稻瘟菌后引起褐点型坏死斑的水稻稻瘟病症状调查结果
调查结果显示,4个IBU浓度处理接种稻瘟病菌72 h时的水稻稻瘟病症状较对照严重(图1-A),100、200、300和400 ?mol/L IBU处理接种稻瘟菌72 h时的水稻病情指数分别为1.23、1.32、1.22和1.26,均高于对照的病情指数(1.20),但差异不显著(P>0.05,下同),而4个IBU浓度处理受侵染水稻间的病情指数差异也不显著(图1-B)。依据4个IBU浓度处理受侵染水稻的稻瘟病病情指数,选取200 ?mol/L IBU开展后续试验。
2. 2 JA和IBU处理接种稻瘟菌后引起褐点型坏死斑的水稻稻瘟病症状调查结果
200 ?mol/L IBU处理接种稻瘟病菌72 h的水稻稻瘟病症状较对照和JA处理的受侵染水稻稻瘟病症状严重(图2-A),病情指数为1.42,显著高于对照(1.04)和JA处理(0.34)(P<0.05,下同),表明IBU处理加重了接种稻瘟菌72 h时的水稻稻瘟病症状,而JA则减轻受侵染水稻稻瘟病症状。
2. 3 JA合成/响应基因在JA和IBU处理接种稻瘟菌后引起褐点型坏死斑水稻中的表达分析结果
利用qRT-PCR分析200 ?mol/L IBU和400 ?mol/L JA处理接种稻瘟菌72 h的水稻内源JA合成基因OsLOX1、OsLOX3、OsJMT1、OsOPR1、OsOPR7、OsAOS2和OsHPL3,以及JA响应基因OsCOI1a、OsCOI1b、OsJAZ1、OsJAZ9、OsMYC2、JiOsPR10和OsbHLH35的表达情况。以无菌水处理接种稻瘟菌72 h的水稻为对照。结果表明,外源JA诱导72 h的水稻JA合成相关基因OsLOX1(图3-A)、OsJIMT(图3-C)、OsOPR1(图3-D)、OsOPR7(图3-E)、OsAOS2(图3-F)和OsHPL3(图3-G)上调表达,至96和120 h时,仅OsAOS2、OsOPR7和OsLOX1上调表达,其余3个基因OsORP1、OsJIMT和OsHPL3则下调表达;未诱导OsLOX3(图3-B)表达。外源JA诱导72 h时的水稻JA响应基因OsCOI1a(图3-H)、OsJAZ1(图3-J)、OsJAZ9(图3-K)、OsMYC2(图3-L)和OsbHLH35(图3-M)上调表达,至120 h时,只有OsCOI1a和OsJAZ1上调表达,其余3个基因OsMYC2、OsJAZ9和OsbHLH35均下调表达;未诱导OsCOI1b(图3-I)和JiOsPR10(图3-N)表达。IBU则抑制上述基因表达(图3-A~图3-N)。因此,外源JA处理接种稻瘟病菌72 h的水稻能诱导水稻植株体内多个JA合成/响应基因表达,而IBU对上述基因表达的影响与外源JA的相反,即IBU抑制外源JA激活基因的表达。
2. 4 SA合成/受体、防御相关、蔗糖/果糖合成酶基因在JA和IBU处理接种稻瘟菌后引起褐点型坏死斑水稻中的表达分析结果
进一步分析200 ?mol/L IBU和400 ?mol/L JA处理接种稻瘟菌72 h的水稻中SA合成基因OsEDS1和OsPAD4、SA受体基因OsNPR4、参与细胞壁合成的蔗糖合成酶基因OsRSUS1和果糖合成酶基因OsFRK-2及防御基因OsPR4a和OsPR5的表达情况,结果发现JA处理的OsPR5(图4-B)在72 h开始上调表达,至96 h时显著上调表达;OsRSUS1(图4-C)在120 h时显著上调表达;OsFRK-2(图4-D)在72、96和120 h时显著上调表达;OsNPR4(图4-G)在72 h时显著上调表达,但96 h时抑制其表达,至120 h时又上调表达但不显著;OsPAD4(图4-F)在72 h时显著上调表达;OsEDS1(图4-E)在120 h时显著上调表达。IBU处理的水稻中除OsEDS1(图4-E)、OsPAD4(图4-F)在72 h时上调表达外,其余5个基因(OsNPR4、OsRSUS1、OsFRK-2、OsPR4a和OsPR5)均下调表达。外源JA未诱导OsPR4a(图4-A)上调表达。表明外源JA能诱导OsPR5、OsRSUS1和OsFRK-2上调表达,而IBU抑制JA激活基因的表达。
3 討论
JA作为多个植物激素的核心组分,参与多个植物激素信号共同调节植物生长发育和胁迫响应(Ahmad et al.,2016;Wasternack and Strand,2016;Yang et al.,2019)。外源施用JA提高植物抗性的研究已有较多报道(Taniguchi et al.,2014;Cao et al.,2016)。本课题组前期研究发现外源JA处理稻瘟病菌强致病菌株接种高感水稻丽江新团黑谷72 h(即褐点型坏死斑开始显现)时稻瘟病症状明显减轻(王云锋等,2018)。在此基础上,本研究进一步发现JA处理强致病菌株接种地方抗病水稻月亮谷72 h时稻瘟病症状也显著减轻,而IBU处理的结果相反,推测外源JA可能激活水稻JA合成和防御响应。进一步分析发现,外源JA诱导JA合成基因OsLOX1、OsOPR7、OsAOS2和JA响应基因OsCOI1a、OsJAZ1、OsJAZ9上调表达,表明外源JA激活了JA合成及其信号通路。本研究检测到JA诱导OsLOX1上调表达而未诱导OsLOX3上调表达,由于OsLOX1和OsLOX3基因结构相似且均参与病原菌防御响应(Maria and Singh,2012),表明JA可能在调控OsLOX1参与JA合成及防御响应中发挥作用。外源JA诱导96和120 h的OsAOS2显著上调表达,但未诱导这两个时间点的OsHPL3上调表达,由于OsAOS2和OsHPL3竞争性地与同一底物结合合成12,13-环氧—十八碳三烯酸(He et al.,2017),表明外源JA主要诱导OsAOS2和OsOPR7参与稻瘟病菌侵染水稻后期内源JA合成(Tani et al.,2008)。
有研究表明,JA诱导OsCOI1a与OsJAZ1和OsJAZ9结合而提高水稻对干旱和盐等非生物逆境的耐受性(Wu et al.,2015;Fu et al.,2017),本研究中JA诱导OsCOI1a、OsJAZ1和OsJAZ9上调表达,表明外源JA也可能同时提高水稻对干旱和盐害等非生物逆境的抗性。OsCOI1b主要参与调控叶片衰老(Yuan and Zhang,2015),外源JA和IBU处理的水稻中OsCOI1b的表达无明显变化,结合侵染水稻稻瘟病症状调查结果,本研究认为外源施用JA并未引起水稻叶片衰老。OsEDS1是受病原菌和SA诱导的防御响应基因(Wiermer et al.,2005;Ke et al.,2019),本研究检测到JA处理的水稻中OsEDS1在120 h时显著上调表达,而OsPAD4只在72 h时上调表达,IBU处理的水稻中OsEDS1在72 h时上调表达,表明OsEDS1在72 h时的表达不受JA诱导。外源JA对SA受体基因OsNPR4表达无显著影响。综上所述,外源JA可能调控OsLOX1、OsOPR7、OsAOS2、OsCOI1a、OsJAZ1和OsJAZ9参与水稻内源JA合成和防御响应。
病原菌、SA、JA、斑蝥素(Cantharidin,CN)和茵多杀(Endothall,EN)能诱导OsPR5表达(Rakwal et al.,2001)。OsPR4a主要受干旱诱导表达(Wang et al.,2011),本研究鉴定到外源JA诱导OsPR5在72和96 h时上调表达,对OsPR4a表达无影响,而IBU抑制两个防御基因表达,因此外源JA诱导OsPR5表达参与水稻防御响应。
细胞壁相关激酶在植物防御响应病原真菌侵染方面具有重要作用(Delteil et al.,2016)。其中,蔗糖合成酶的作用之一是引导UDP-葡萄糖合成纤维素等细胞壁多糖(Haigler et al.,2001),果糖合成酶则是磷酸化果糖进入细胞质后成为细胞壁合成的组分,特别是在木质素和导管的细胞壁发育中起作用(Stein and Granot,2018)。本研究中外源JA诱导蔗糖合成酶基因OsRSUS1在120 h时显著上调表达,果糖合成酶基因OsFRK-2在72、96和120 h时显著上调表达,表明外源JA通过诱导蔗糖/果糖合成酶基因表达在参与受侵染水稻后期细胞壁重建中发挥作用。
4 结论
外源JA主要通过诱导稻瘟菌引起褐点型坏死斑的水稻JA合成/响应基因、参与细胞壁合成的蔗糖/果糖合成酶基因及防御相关基因表达参与水稻防御响应。
参考文献:
曹慧娟,张瑾瑾,杜艳,齐中强,俞咪娜,刘永锋. 2019. 稻瘟病菌转录因子研究进展[J]. 江苏农业学报,35(6):1493-1500. [Cao H J,Zhang J J,Du Y,Qi Z Q,Yu M N,Liu Y F. 2019. Research progress of transcription factors in the rice blast fungus[J]. Jiangsu Journal of Agricultural Sciences,35(6):1493-1500.]
王云锋,王长秘,李春琴,刘林,李晓杰,李晓疆,杨静. 2018. 稻瘟病菌侵染时水稻防御体系对外源茉莉酸的响应分析[J]. 南方农业学报,49(7):1324-1331. [Wang Y F,Wang C M,Li C Q,Liu L,Li X J,Li X J,Yang J. 2018. Response of rice defense system to exogenous jamonisic acids during rice blast strain infecting rice[J]. Journal of Southern Agriculture,49(7):1324-1331.]
许志刚. 2002. 普通植物病理学[M]. 第3版. 北京:中国农业出版社:319-335. [Xu Z G. 2002. General plant pathology[M]. The 3rd Edition. Beijing:China Agriculture Press: 319-335.]
Agrawal G K,Jwa N ,Rakwal R. 2000a. A novel rice(Oryza sativa L.) acidic PR1 gene highly responsive to cut,phytohormones,and protein phosphatase inhibitors[J]. Biochemical & Biophysical Research Communications,274(1): 157-165.
Agrawal G K,Rakwal R,Jwa N S. 2000b. Rice(Oryza sativa L.) OsPR1b gene is phytohormonally regulated in close interaction with light signals[J]. Biochemical & Biophy-sical Research Communications,278(2): 290-298.
Ahmad P,Rasool S,Gul A,Sheikh S A,Akram N A,Ashraf M,Kazi A,Gucel S. 2016. Jasmonates: Multifunctional roles in stress tolerance[J]. Frontiers in Plant Science,7:813.
Bhadauria V,Banniza S,Vandenberg A,Selvaraj G,Wei Y. 2013. Overexpression of a novel biotrophy-specific colleto-trichum truncatum effector,ctnudix,in hemibiotrophic fungal phytopathogens causes incompatibility with their host plants[J]. Eukaryotic Cell,12(1): 2-11.
Cao J J,Li M Y,Chen J,Liu P,Li Z. 2016. Effects of MeJA on Arabidopsis metabolome under endogenous JA deficiency[J]. Scientific Reports,6:37674.
Dangl J L,Horvath D M,Staskawicz B J. 2013. Pivoting the plant immune system from dissection to deployment[J]. Science,341(6147):746-751.
Delteil A,Gobbato E,Cayrol B,Estevan J,Michel-Romiti C,Dievart A,Kroj T,Morel J B. 2016. Several wall-associa-ted kinases participate positively and negatively in basal defense against rice blast fungus[J]. BMC Plant Biology,16:1-10.
Fu J,Wu H,Ma S,Xiang D,Liu R,Xiong L. 2017. OsJAZ1 attenuates drought resistance by regulating JA and ABA signaling in rice[J]. Frontiers in Plant Science,8:2018.
García-Guzmán G,Heil M. 2014. Life histories of hosts and pathogens predict patterns in tropical fungal plant diseases[J]. New Phytologist,201(4):1106-1120.
Gurjar M S,Ali S,Akhtar M,Singh K S. 2012. Efficacy of plant extracts in plant disease management[J]. Agricultu-ral Sciences,3(3): 425-433.
Haigler C H,Ivanova-Datcheva M,Hogan P S,Salnikov V V,Hwang S,Martin K,Delmer D P. 2001. Carbon partitioning to cellulose synthesis[J]. Plant Molecular Biology,47(1-2):29-51.
Hasan N,Rafii M Y,Abdul R H,Nusaibah S A,Mazlan N,Abdullah S. 2017. Genetic analysis and identification of SSR markers associated with rice blast disease in a BC2F1 backcross population[J]. Genetics and Molecular Research. doi:10.4238/gmr16019280.
He X,Li L,Xu H,Xi J,Cao X,Xu H,Rong S,Dong Y,Wang C,Chen R,Xu J,Gao X,Xu Z. 2017. A rice jacalin-related mannose-binding lectin gene, OsJRL,enhances Escherichia coli viability under high salinity stress and improves salinity tolerance of rice[J]. Plant Biology,19(2):257-267.
Ke Y G,Kang Y R,Wu M X,Liu H B,Hui S G,Zhang Q G,Li X H,Xiao J H,Wang S P. 2019. Jasmonic acid-involved OsEDS1 signaling in rice-bacteria[J]. Rice,12:25.
Koga H. 1994. Hypersensitive death,autofluorescence,and ultrastructural changes in cells of leaf sheaths of susceptible and resitant near-isogenic lines of rice(Pi-zt) in relation to penetration and growth of Pyricularia oryzae[J]. Canadian Journal of Botany,72:1463-1477.
Law J W F,Ser H L,Khan T M,Chuah L H,Pusparajah P,Chan K G,Goh B H,Lee L H. 2017. The potential of streptomyces as biocontrol agents against the rice blast fungus,Magnaporthe oryzae(Pyricularia oryzae)[J]. Frontiers in Microbiology,8:3.
Maria S S,Singh V K. 2012. LOX genes in blast fungus (Magnaporthe grisea) resistance in rice[J]. Functional & Integrative Genomics,12(2):265-275.
Nalley L T,Tack J,Barkley A,Jagadish K,Brye K. 2016. Quantifying theagronomic and economicperformance of hybrid and conventional rice varieties[J]. Agronomy Journal,108(4):1514.
Oka K,Kobayashi M,Mitsuhara I,Seo S. 2013. Jasmonic acid plays a negative role in resistance to tobacco mosaic virus in tobacco[J]. Plant & Cell Physiology,54(12): 1999-2010.
Rakwal R,Agrawal G K,Agrawal V A. 2001. Jasmonate,salicylate,protein phosphatase 2A inhibitors and kinetinup-regulate OsPR5 expression in cut-responsive rice(Oryza sativa)[J]. Jorunal of Plant Physiology,158:1357-1362.
Shang J G,Yu S S,Gu F Q,Xu Z Y,Zhu L F. 2011. A mobile guide system framework for museums based on local location-aware approach[C]. 2011 international conference on computer science and service system(CSSS),1935-1940.
Stein O,Granot D. 2018. Plant fructokinases: Evolutionary,developmental,and metabolic aspects in sink tissues[J]. Frontiers in Plant Science. doi:10.3389/fpls.2018.00339.
Tani T,Sobajima H,Okada K,Chujo T,Arimura S,Tsutsumi N,Nishimura M,Seto H,Nojiri H,Yamane H. 2008. Identification of the OsOPR7 gene encoding 12-oxophytodienoate reductase involved in the biosynthesis of jasmonic acid in rice[J]. Planta,227(3): 517-526.
Taniguchi S,Hosokawa-Shinonaga Y,Tamaoki D,Yamada S,Akimitsu K,Gomi K. 2014. Jasmonate induction of the monoterpene linalool confers resistance to rice bacterial blight and its biosynthesis is regulated by JAZ protein in rice[J]. Plant,Cell and Environment,37:451-461.
Wang N,Xiao B,Xiong L. 2011. Identi?cation of a cluster of PR4-like genes involved in stress responses in rice[J]. Journal of Plant Physiology,168(18):2212-2224.
Wasternack C,Strnad M. 2016. Jasmonate signaling in plant stress responses and development-active and inactive compounds[J]. New Biotechnology,33(5):604-613.
Wiermer M,Feys B J,Parker J E. 2005. Plant immunity: The EDS1 regulatory node[J]. Current Opinion in Plant Bio-logy,8(4):383-389.
Wu H,Ye H Y,Yao R F,Zhang T,Xiong L Z. 2015. OsJAZ9 acts as a transcriptional regulator in jasmonate signaling andmodulates salt stress tolerance in rice[J]. Plant Science,232: 1-12.
Yang J,Duan G H,Li C Q,Liu L,Han G G,Zhang Y L,Wang C M. 2019. The crosstalks between JA and other plant hormone signalings highlight the involvement of JA as a core component in plant response to biotic and abio-tic stresses[J]. Frontiers in Plant Science,10:1349.
Yuan Z,Zhang D B. 2015. Roles of jasmonate signalling in plant in?orescence and?ower development[J]. Current Opinion in Plant Biology,27:44-51.
(责任编辑 麻小燕)
