外源褪黑素对盐胁迫下香椿种子萌发及幼苗生长的影响
2020-06-19刘珂张嘉欣杜清洁刘珂珂马勇斌丁新新肖怀娟
刘珂 张嘉欣 杜清洁 刘珂珂 马勇斌 丁新新 肖怀娟
摘 要: 為探究褪黑素对盐胁迫环境下香椿(Toona sinensis)种子萌发及幼苗生长的影响,以香椿为试材,采用清水处理为对照,利用NaCl模拟盐胁迫环境,分别用0、50、100、200、300 ?mol·L-1的褪黑素处理香椿种子及幼苗,测定种子萌发后的各项生理指标。结果表明,盐胁迫处理显著抑制了香椿种子的发芽率、发芽势、发芽指数和根长,以及幼苗叶片的叶长、叶宽。与盐胁迫处理相比,100 ?mol·L-1褪黑素预处理的香椿种子萌发和幼苗生长均得到显著改善,发芽率、发芽势、发芽指数分别显著提高了15.56%、18.89%、12.21%,叶长和叶宽提高了9.26、10.10 mm。盐胁迫导致香椿幼苗丙二醛含量和过氧化物酶活性显著高于对照,而超氧化物歧化酶活性显著低于对照。低浓度褪黑素预处理则显著降低了盐胁迫下丙二醛含量和过氧化物酶活性,提高了超氧化物酶活性。其中,提高香椿幼苗耐盐性的最适褪黑素浓度为100 ?mol·L-1,与盐胁迫相比,丙二醛含量和过氧化物酶活性分别降低了6.19 ?mol·g-1、 6 292.57 U·g-1,超氧化物酶活性提高了22.16 U·g-1。
关键词: 香椿;褪黑素;盐胁迫;萌发;生长
中图分类号:S644.4 文献标志码:A 文章编号:1673-2871(2020)05-053-06
Abstract: To explore the effect of melatonin on seed germination and seedling growth of Toona sinensis under salt stress. Toona sinensis was used as the test material, NaCl was used to simulate salt environment. Toona sinensis seeds and seedlings were treated with 0, 50, 100, 200 and 300 ?mol·L-1 melatonin, respectively, and water treatment was used as control. The root length, germination rate, germination potential, germination index, leaf length, leaf width, chlorophyll content, superoxide dismutase, peroxidase and malondialdehyde were measured. The results showed that the germination rate, germination potential, germination index, root length, the leaf length and the leaf width of seedling leaves were significantly inhibited by salt stress. Compared with salt stress treatment, seed germination and seedling growth of Toona sinensis were significantly improved after 100 ?mol·L-1 melatonin pretreatment, germination rate, germination potential, germination index, leaf length, leaf width were significantely improved 15.56%, 18.89%, 12.21%, 9.26 mm and 10.10 mm, respectively. Under salt stress, malondialdehyde content and peroxidase activity in Toona sinensis seedlings were significantly higher than those of control, while superoxide dismutase activity was significantly lower than that of the control. Malondialdehyde content and peroxidase activity under salt stress were significantly reduced by low concentration of melatonin pretreatment, while superoxide activity was increased. The optimum melatonin concentration to improve the salt tolerance of Toona sinensis seedlings was 100 ?mol·L-1, compared with salt stress treatment, the decrease of peroxidase activity and malondialdehyde were reduced 6.19 ?mol·g-1, 6 292.57 U·g-1, respectively, and superoxide dismutase was increased 22.16 U·g-1.
Key words: Toona sinensis; Melatonin; Salt stress; Germination; Growth
香椿(Toona sinensis)为楝科香椿属植物,原产中国,以食用嫩芽为主。香椿芽具有较高的营养价值,并且风味独特,散发一股浓郁香气,可以增强人的食欲。随着人们对身体健康的重视,香椿也成为高营养价值蔬菜的典型代表[1]。近年来,各地利用种子繁殖来实现香椿生产[2]。但由于栽培过程中化肥的过度使用和保护地栽培中土壤缺乏雨水冲刷等多方面原因,造成土壤盐渍化现象越来越多,已严重制约了香椿芽苗和成树生产的规模。因此,进行香椿种子及香椿幼苗的耐盐生理研究,对扩张香椿种植面积、有效利用盐渍化土壤和实现香椿高产有重大意义。
土壤中可溶性盐浓度过大,一方面会造成土壤水势过低,低于细胞水势时发生植物组织水分外渗,引起植物生理性缺水,另一方面由于单一离子含量较高,会产生单盐毒害并且影响植物对其他元素的吸收,最终导致植物细胞内的自由基代谢动态平衡被打破,活性氧(ROS)增加,破坏细胞膜结构,选择透性丧失,有毒物质积累,植物代谢紊乱,影响植物的生长发育[3-5]。姚侠妹等[4]用不同浓度的NaCl处理香椿幼苗,结果发现随着 NaCl 浓度提高,香椿幼苗叶长、叶宽和株高呈降低趋势,且影响香椿的光合作用,进而影响了香椿幼苗的生长。
褪黑素(MT)是广泛存在于动植物体内的一种低分子量(232.27)吲哚胺类化合物,1958年被LERNER在牛的松果体中首次提取出来[6]。MT作用于植物体的整个生长发育阶段,可以调控植物的营养器官和生殖器官发育以及调控光合效率和叶片衰老,除此之外,MT还可以增强植物对冷热胁迫、渗透胁迫、氧化胁迫、病菌胁迫的耐受性[7]。唐懿等[8]研究表明,MT可以提高低温弱光胁迫下茄子幼苗叶片的叶绿素含量,提高过氧化物酶(POD)、超氧化物歧化酶(SOD)、过氧化氢酶(CAT)活性,降低丙二醛(MDA)的含量,促进可溶性糖积累,提高茄子幼苗叶片对低温弱光胁迫的抵抗力等。目前,MT对大豆、茄子、苹果、水稻、紫苏、猕猴桃等[3,8-13]已有研究,鲜见MT对香椿的相关研究报道。因此,笔者以香椿为试材,采用种子阶段前期MT浸泡处理,幼苗阶段采用外源喷施MT预处理,测定盐胁迫下香椿种子发芽率、发芽势和香椿幼苗叶片的抗氧化酶活性等指标,探讨MT在香椿抵抗盐胁迫方面的生理机制,筛选出适宜的MT浓度,旨在为香椿栽培中利用MT提高作物耐盐性提供理论依据。
1 材料与方法
1.1 材料
香椿材料为‘红油香椿,产自燕山山脉地区,由河南农业大学园艺学院设施蔬菜栽培课题组提供;凹形定植棉规格为2.5 cm×2.5 cm×2.5 cm,水培槽规格为44 cm×28 cm×5 cm。试验于2019年2月27日至4月13日在河南农业大学园艺学院设施蔬菜实验室进行。
1.2 方法
1.2.1 种子处理及萌发试验 挑选籽粒饱满、大小一致的香椿种子用75%的酒精溶液消毒30 s后,无菌水冲洗3次,每次2 min。室内回干到种子初始含水量。回干后的种子浸泡在MT溶液中进行预处理。MT溶液浓度分别为0、50、100、200、300 ?mol·L-1(分别为处理A、B、C、D、E),浸泡时间为12 h。再次在室内回干到种子原始含水量,在直径为90 mm的培养皿中放2層无菌滤纸,每皿添加5 mL的100 mmol·L-1 NaCl溶液为发芽溶液。对照(CK)采用清水对种子进行预处理,过程同其他处理,且以清水为发芽溶液。种子在人工气候室(RLD-380E-4)中进行萌发,培养条件为温度20 ℃,相对湿度70%,黑暗培养。采用完全随机处理,每个处理3次重复,每重复为30粒种子。培养期间每隔3 d补充发芽溶液1 mL。
为选择适宜盐浓度的发芽溶液,首先进行盐处理试验,将未被MT溶液处理过的香椿种子在不同浓度(0、50、100、200、300 mmol·L-1)NaCl溶液中进行发芽,测定种子的发芽率和发芽势,最后选定刚好能够抑制香椿种子萌发的盐胁迫处理浓度为100 mmol·L-1 。
1.2.2 发芽率、发芽势、发芽指数和芽长的测定 在处理第5天统计各处理的发芽数,计算发芽势。处理第10天统计各处理的发芽数,计算发芽率及发芽指数。用20 cm直尺测量种子处理10 d后的芽长。发芽势、发芽率和发芽指数的计算参照李平平等[2]的方法。
1.2.3 幼苗培养及处理 挑选籽粒饱满、大小一致的香椿种子,温汤浸种后放入直径为150 mm的培养皿中(放2层无菌滤纸),每皿添加10 mL的蒸馏水,在温度20 ℃、相对湿度70%的光照培养箱暗光培养中催芽。待种子露白达80%后播于凹形定植棉中进行水培培养。待香椿幼苗子叶展平后从培养箱中取出,使用水培槽进行水培,放在人工气候室培养。环境条件设置:昼/夜光周期为16 h/8 h,光强为10 000 lx,昼/夜温度为23 ℃/18 ℃。
待幼苗2叶1心时,随机选取120株幼苗平均分为5组,各喷施0、50、100、200、300 μmol·L-1的外源MT(分别为处理A、B、C、D、E),连续喷施3 d,然后在营养液中添加100 mmol·L-1的NaCl。采用喷施清水+正常营养液作为对照(CK)。添加NaCl 7 d后,测定各项指标。试验设计为完全随机处理,每个处理设3次重复,每个重复24株。
1.2.4 形态指标的测定 盐胁迫处理7 d后,各处理随机选取3株植株,用数显游标卡尺(DL91150)测量植株叶片的长和宽。
1.2.5 生理指标的测定 盐胁迫处理7 d后取样测量叶绿素等生理指标。其中,叶绿素含量采用无水乙醇与丙酮混合法测定[14];超氧化物歧化酶(SOD)、过氧化物酶(POD)的活性与丙二醛(MDA)含量采用张海波[15]的方法测定。
1.3 数据处理
试验数据采用IBM SPSS Statistics 24.0软件进行单因素方差分析,用LSD法对比各处理间差异。
2 结果分析
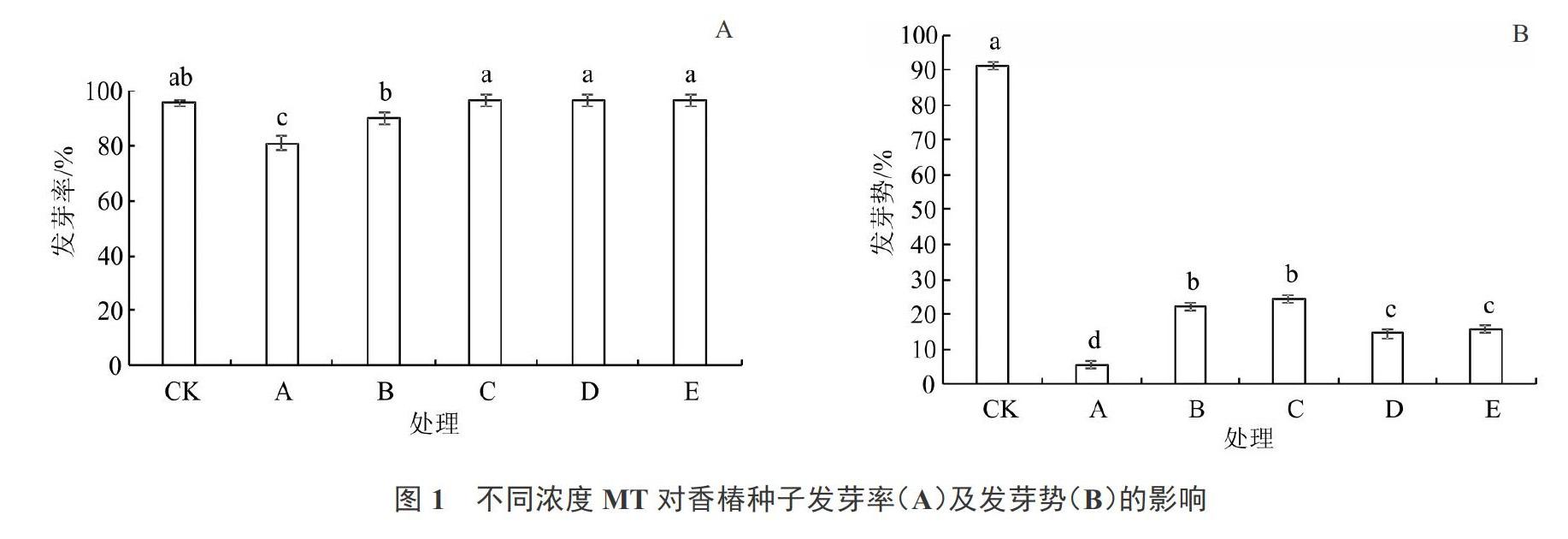
2.1 不同浓度MT对香椿种子发芽率及发芽势的影响
由图1可知,与CK相比,盐胁迫处理下香椿种子发芽率和发芽势显著降低,MT预处理可显著缓解盐胁迫处理对发芽率和发芽势的抑制作用。盐胁迫处理下,处理B~E(50、100、200、300 ?mol·L-1 MT预处理)的种子发芽率分别比处理A(无MT预处理)增加了8.89%、15.56%、15.56%、15.56%,均与CK无显著差异(图1-A)。同时,处理B~E使盐胁迫下种子发芽势(图1-B)由处理A的5.56%分别显著提高到了22.22%、24.45%、14.44%、15.56%。这说明MT预处理可显著提高盐胁迫下香椿种子的发芽率和发芽势,其中50和100 ?mol·L-1的MT预处理效果最佳。
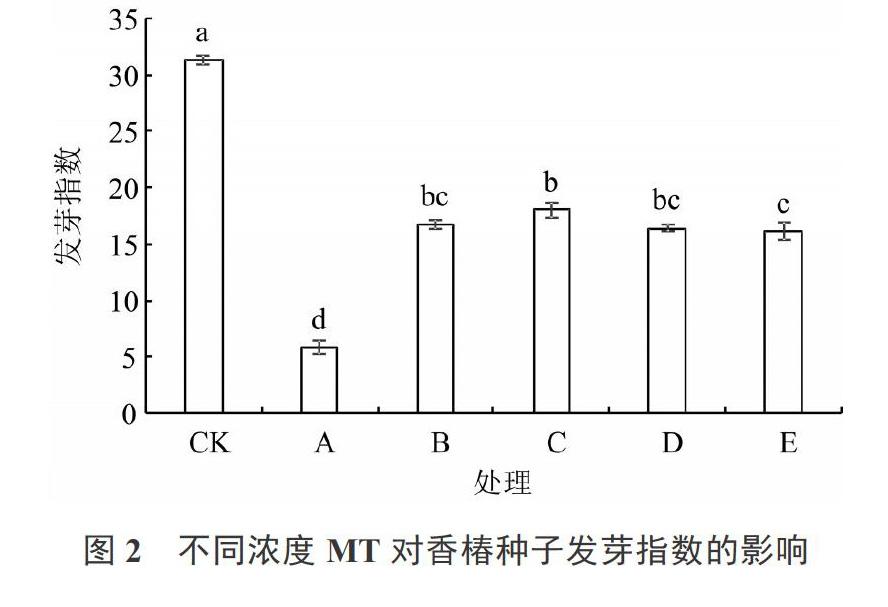
2.2 不同浓度MT对香椿种子发芽指数的影响
由图2可知,盐胁迫下处理A香椿种子的发芽指数较CK降低了81.47%,达显著水平。盐胁迫处理下,与无MT预处理相比,MT预处理均大幅促进了种子的发芽指数,其中,处理C(100 ?mol·L-1 MT预处理)的发芽指数最高,达到CK的57.52%,其次为处理B、D、E,分别为CK的53.49%、52.47%、51.52%。
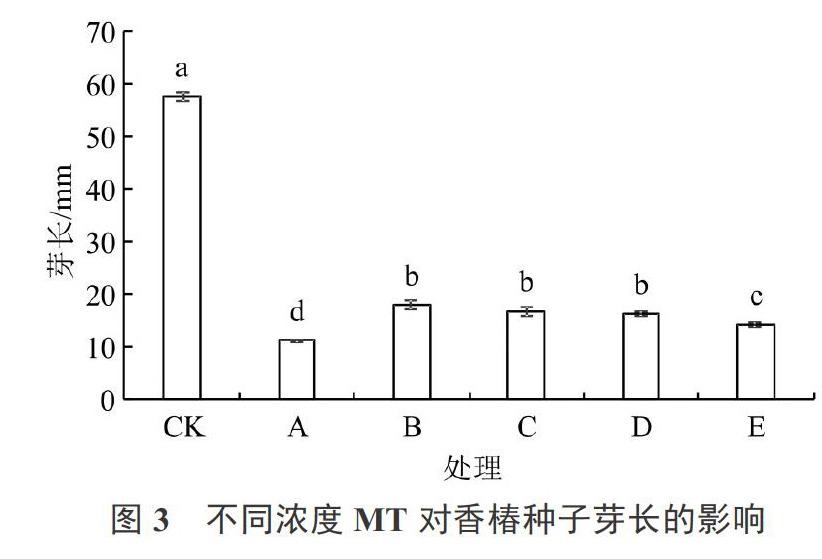
2.3 不同浓度MT对香椿种子芽长的影响
由图3可以看出,盐胁迫处理也显著抑制了香椿种子的芽伸长。同盐胁迫下无MT预处理(处理A)相比,香椿种子的芽长在MT预处理下显著增大,其中,处理B~D之间无显著差异,且显著高于处理E。说明低浓度MT预处理能显著缓解盐胁迫对香椿种子芽伸长的抑制作用,但高浓度MT的缓解效果较差。
2.4 不同浓度MT对香椿幼苗叶长和叶宽的影响
由图4可以看出,盐胁迫明显抑制了香椿幼苗植株的长势,MT预处理可以在不同程度上缓解盐胁迫对幼苗长势的抑制。香椿幼苗叶长和叶宽均以盐胁迫处理(处理A)下最小,显著低于CK;以处理C(100 ?mol·L-1 MT预处理)最大,分别为处理A的1.28倍和1.37倍,且显著高于CK;处理D(200 ?mol·L-1 MT预处理)均显著高于盐胁迫处理,但叶宽与CK无显著差异;处理B(50 ?mol·L-1 MT预处理)均显著高于盐胁迫处理,且显著低于CK;处理E(300 ?mol·L-1 MT预处理)均显著高于盐胁迫处理,且显著低于CK。表明50、100 和200 ?mol·L-1 MT预处理都对缓解盐胁迫下香椿幼苗叶片长度和宽度作用显著,其中100 ?mol·L-1 MT预处理的效果最好。
2.5 不同浓度MT对香椿幼苗叶绿素含量和MDA含量的影响
由图5-A可以看出,各处理间香椿幼苗叶绿素含量无显著差异,表明盐胁迫和MT预处理均不影响香椿幼苗的叶绿素代谢。由图5-B可以看出,与CK相比,盐胁迫处理下MDA含量大幅升高,为CK的1.55倍。MT预处理均使MDA含量较盐胁迫处理显著下降,但处理B、处理D和处理E(50、200、300 ?mol·L-1 MT预处理)的MDA含量显著高于CK,处理C(100 ?mol·L-1 MT预处理)的MDA含量则显著低于CK。表明MT预处理均能缓解盐胁迫下香椿幼苗MDA的积累,其中100 ?mol·L-1 MT预处理的作用最好。
2.6 不同浓度MT對香椿幼苗POD和SOD含量的影响
由图6-A可知,盐胁迫处理下POD活性为CK的1.65倍,差异达显著水平。MT预处理则不同程度地抑制盐胁迫下POD活性的升高。与盐胁迫处理相比,处理B~E的POD活性分别降低了30.37%、43.82%、36.89%和34.17%。由图6-B可知,香椿幼苗SOD活性在盐胁迫下较CK显著下降。处理B和处理C(50和100 ?mol·L-1 MT预处理)可显著缓解SOD活性的降低。与盐胁迫相比,处理D和处理E(200和300 ?mol·L-1 MT预处理)的SOD活性有所下降,其中处理E的SOD活性与盐胁迫的差异显著。上述结果表明,MT可抑制香椿幼苗的POD活性;低浓度的MT可促进SOD活性的增加,高浓度的反而抑制SOD的活性。
3 讨论与结论
土壤中盐离子含量对植物生长、发育和产量形成具有很大影响,过高的盐含量可破环植物体内的新陈代谢过程。MT作为一种植物生长调节剂,对植物的生长发育有重要调节作用,研究MT对盐胁迫下植物种子萌发及幼苗生长的影响是非常重要的。前人研究结果表明,MT预处理可显著缓解盐胁迫对黄瓜、玉米和棉花种子的抑制作用[16-18]。本研究结果也表明,在盐胁迫下MT预处理过的香椿种子发芽率、发芽势、发芽指数和根长均较无MT预处理的有所提高,且100 ?mol·L-1 MT预处理下各指标均达显著水平。张海军等[16]研究发现,外源褪黑素处理能够明显提高氯化钠胁迫下黄瓜种子中SOD、POD和CAT活性,清除活性氧,降低膜质过氧化程度,从而缓解盐胁迫对种子造成的抑制作用。转录组和miRNA组证据也表明,MT预处理能显著提高盐胁迫下玉米种子的萌发过程中POD、CAT、SOD等抗氧化酶的表达[17]。
笔者发现,MT预处理可缓解盐胁迫对香椿幼苗长势的抑制作用,降低盐胁迫下香椿幼苗叶片脂质过氧化物MDA的积累。Mittler[19]指出,高等植物在受到氧化胁迫时,会自身形成一套主要包括SOD、POD、CAT和APX等酶促系统的完整抗氧化体系链条。本研究中,低浓度MT预处理可提高幼苗叶片中SOD活性。SOD可清除植物体内的超氧阴离子,使植物体内的活性氧含量减少。因此,MT可通过调控SOD活性来减缓盐胁迫下膜脂过氧化对香椿细胞的伤害,促进香椿幼苗的生长,与前人研究结果一致[13, 20-21]。POD作为一个重要的抗氧化酶,可清除细胞内ROS,但同时也参与木质素的合成,增加木质化程度,可以作为组织老化的一种生理指标[22-23]。本研究中MT预处理后的香椿幼苗在盐胁迫下的POD活性显著降低,以100 ?mol·L-1 MT预处理下降低幅度最大。这表明MT预处理主要通过提高香椿幼苗叶片中SOD的活性来清除盐胁迫诱导产生的ROS,减缓了组织老化进程。
综上所述,盐胁迫下香椿种子的萌发以及幼苗的生长受到抑制,MT可有效缓解盐胁迫对香椿种子和幼苗的损害。MT浓度为100 μmol·L-1时效果最好,为后续香椿种子萌发和幼苗生长的进一步研究提供理论参考和技术支持。
参考文献
[1] 贾韶千.香椿的食用价值和市场前景[J].食品安全导刊,2016(15):82-83.
[2] 李平平,苗秀琴,武军杨,等.壳聚糖在香椿种子萌发及幼苗生长上的生理效应[J].西北林学院学报,2011,26(6):44-47.
[3] 刘月,寇从贤,付桂萍,等.褪黑素对大豆幼苗盐害的缓解效应及机理研究[J].中国油料作物学报,2017,39(6):813-819.
[4] 姚侠妹,偶春,王群群,等.盐胁迫对香椿幼苗生长和光合特性的影响[J].阜阳师范学院学报(自然科学版),2019,36(1):36-39.
[5] DAVIES K J.An overview of oxidative stress[J].IUBMB Life,2000,50(4/5):241-244.
[6] 陆炀,燕志,要志宏,等.褪黑素的应用[J].江苏调味副食品,2018(1):7-9.
[7] 张杰,顾红,程大伟,等.植物褪黑素研究[J].江西农业学报,2018,30(6):39-48.
[8] 唐懿,余雪娜,何娟,等.外源褪黑素对低温弱光胁迫下茄子幼苗生理特性的影响[J].湖南农业大学学报(自然科学版),2017,43(3):257-261.
[9] WANG P,YIN L H,LIANG D,et al.Delayed senescence of apple leaves by exogenous melatonin treatment:toward regulating the ascorbate-glutathione cycle[J].Journal of Pineal Research,2012,53(1):11-20.
[10] 宋雪飞,甘淳丹,赵海燕,等.叶面喷施褪黑素调控水稻幼苗耐盐性的浓度效应研究[J].土壤学报,2018,55(2):455-466.
[11] 李爱,孙汪亮,李林,等.外源褪黑素对干旱胁迫下紫苏幼苗生长的影响[J].江苏农业科学,2018,46(2):70-73.
[12] 高帆,谢玥,沈妍秋,等.外源褪黑素对氯化钠胁迫下美味猕猴桃实生苗抗氧化物酶和渗透调节物质的影响[J].浙江农林大学学报,2018,35(2):291-297.
[13] 倪知游,夏惠,高帆,等.外源褪黑素对猕猴桃幼苗盐胁迫的缓解作用[J].四川农业大学学报,2017,35(4):535-539.
[14] 苏正淑,张宪政.几种测定植物叶绿素含量的方法比较[J].植物生理学通讯,1989(5):77-78.
[15] 张海波.香椿种子特定贮藏条件下活力变化的研究[D].北京:中国林业科学研究院,2018.
[16] ZHANG H J, ZHANG N, YANG R C,et al.Melatonin promotes seed germination under high salinity by regulating antioxidant systems,ABA and GA4 interaction in cucumber (Cucumis sativus L.)[J].Journal of Pineal Research,2014,57(3):269-279.
[17] 徐暑晖.外源褪黑素促进盐胁迫下玉米种子萌发的分子机理[D].江苏扬州:扬州大学,2018.
[18] 袁志刚.外源褪黑素在维持盐胁迫下棉花种子萌发和幼苗生长中的应用[D].河北秦皇岛:河北科技师范学院,2018.
[19] MITTLER R.Oxidative stress,antioxidants and stress tolerance[J].Trends in Plant Science,2002,7(9):405-410.
[20] 杜天浩.外源褪黑素对NaCl胁迫下番茄幼苗生理代谢和产量及品质的影响[D].陕西杨凌:西北农林科技大学,2016.
[21] 李猛,陈栋,李秀妮,等.盐胁迫下外源褪黑素对烟草幼苗抗氧化特性和光合特性的影响[J].中国农业科技导报,2019,21(2):141-147.
[22] LI Y X,WILLS R B H,GOLDING J B.Interaction of ethylene concentration and storage temperature on postharvest life of the green vegetables pakchoi,broccoli,mint,and green bean[J].The Journal of Pomology & Horticultural Science and Biotechnology,2017,92 (3):288–293.
[23] 蔡沖,龚明金,李鲜,等.枇杷果实采后质地的变化与调控[J].园艺学报,2006,33(4):731-736.
