桂子山鸟类群落种类组成、结构及多样性分析
2020-06-18熊广泽蒙建忠张洪茂
熊广泽,周 权,蒙建忠,张洪茂
(华中师范大学生命科学学院进化与生态学研究所,武汉 430079)
鸟类因具有分布广泛,种类多样以及易于识别等特点,是研究最多、调查最频繁的动物类群之一[1].作为重要的生态系统组成部分,因其对栖息地环境改变极为敏感,故可以在一定程度上反映相应生态环境的变化.鸟类的分布、数量、物种多样性等特征可作为生物多样性和环境监测的重要指标[1-2].城市扩建引起的环境变迁和森林高度斑块化,对动物多样性及其生态功能具有重要的负面影响,但相关研究缺乏长期监测的数据支持.近十年来,武汉市城市建设持续发展,连年的建设施工使城市环境发生了很大变化,诸多原本适宜鸟类栖息的环境被破坏.同时,近年来武汉市也开始大规模地进行环境整治和生态环境修复.但城市扩建和环境整治对城市生态系统生物多样性的影响,尚无较详细的监测案例.因此,监测某一环境条件相对稳定的区域(如高校、森林公园等)的鸟类群落,对了解生态环境变化对生物多样性的影响具有较为重要的意义.
桂子山是华中师范大学主校区,属于受城市建设干扰相对较小的区域,校内环境一直比较适合伴人居鸟类栖息,适合研究城市迅速扩建条件下,鸟类群落种类组成及变化.此外,距最近一次发表对桂子山全年的鸟类调查已过去24年[3],距最近一次发表对桂子山冬季的鸟类调查也已过去12年[4].在城市快速扩建背景下,近十年来桂子山区域鸟类发生了何种变化尚无系统的报道.
本文旨在对桂子山的鸟类群落种类组成、空间分布、居留类型等进行报道,通过比对过去的数据,初步反映城市快速扩建背景下,生态环境变化对鸟类的影响,以为城市森林斑块功能评估、生态文明城市建设以及校园鸟类保护提供一定的参考依据.
1 研究方法
1.1 调查地区概况
武汉地处江汉平原东部,长江与汉水交汇处,河网密集,地形为残丘性冲积平原,属亚热带湿润季风气候,雨量充沛,日照充足,四季分明.武汉市全域海拔在19.2~873.7 m[5].武汉地处北亚热带和中亚热带分界区,在生物地理分布区划上,靠近东洋界北缘[6],主城区植被类型以中亚热带常绿阔叶林为主[7],鸟类资源丰富,留鸟众多,同时也是我国很多冬候鸟的中转站和越冬地.
桂子山位于武汉市主城区东部,东湖以南,南湖以北,是华中师范大学及周边居民区、商业区的集中分布区.地理位置为30°31′~30°32′ N,114°21′~114°22′ E,面积102.41 hm2.海拔32~60 m.桂子山区域植被类型为常绿和落叶阔叶林,绿化覆盖率74%.其中,乔木以樟(Cinnamomumcamphora)、二球悬铃木(Platanusacerifolia)、马尾松(Pinusmassoniana)、木犀(Osmanthusfragrans)、雪松(Cedrusdeodara)等较为常见;灌木多以杜鹃(Rhododendronsimsiis)、海桐(Pittosporumtobiras)、八角金盘(Fatsiajaponica)、棕竹(Rhapisexcelsa)、花叶青木(Aucubajaponicavar.variegata)、红花檵木(Loropetalumchinensevar.rubrum)等较常见;草本多以吉祥草(Reineckiacarnea)、白车轴草(Trifoliumrepens)、红花酢浆草(Oxaliscorymbosa)、车前草(Plantagoasiatica)、一年蓬(Erigeronannuus)、喜旱莲子草(Alternantheraphiloxeroides)、阿拉伯婆婆纳(Veronicapersica)等较常见.植被多样性相对较高,生境多样,为鸟类的栖息提供了较良好自然条件.
1.2 调查方法
采用样线法(line transect)和样点法(point count)对鸟类进行调查.根据调查区域的植被分布与生境特征,调查范围涉及4种主要生境.1) 建筑及周边:指教学楼、学生宿舍、教职工居住区、食堂等人流量较大的人工建筑及其周边的空地、绿化带组成的生境,所占面积大,受人类干扰较大,植被较稀疏;2) 乔木林:指山体上发育良好的次生林和部分高大的成片的行道树所形成的生境,主要分布在桂子山北坡部分杉树、松树林、桂子山东坡环山东路的樟树林、元宝山北麓的马尾松林、桂中路及周边道路两边高大茂盛的悬铃木林等处,所占面积较小,受人类干扰影响较小,植被高度和盖度大,地表少灌木草本;3) 乔灌混合林:包括校园内大部分保存较完好的次生林和绿地,主要由乔木层、林下及林间灌草丛组成,往往是近年翻新的绿地等,主要分布在桂子山南坡、玉兰园、图书馆北面、杜鹃广场等处,所占面积较大,受人类干扰影响相对较小,植被多样性较高、灌丛密集;4) 草地:包括两个草坪广场和各种成片的观赏草坪,主要由低矮密集茂盛的草本植物组成,有时伴有稀疏的几株乔木,主要分布在博雅广场和喷泉广场等处,所占面积较小,受人类干扰影响较大,植被多样性低.除此之外,桂子山区域还有总面积约1 100 m2的3个人工水池,因面积较小,故归入周围生境.调查采用固定样线法和样点法,其中,样线一共设置3条(图1),每条样线长1.0~2.0 km,单侧宽50 m,样线总长度4.3 km,相邻样线间隔约500 m,观测时的步行速度1~1.5 km·h-1[1-2,4].一共设置4个固定观察样点 (图1),每个半径约25~50 m[4],样点间间距约500 m,每次观察时,等周围鸟类适应观察者后开始计数,每次每个样点的观测时间为10~15 min[8].
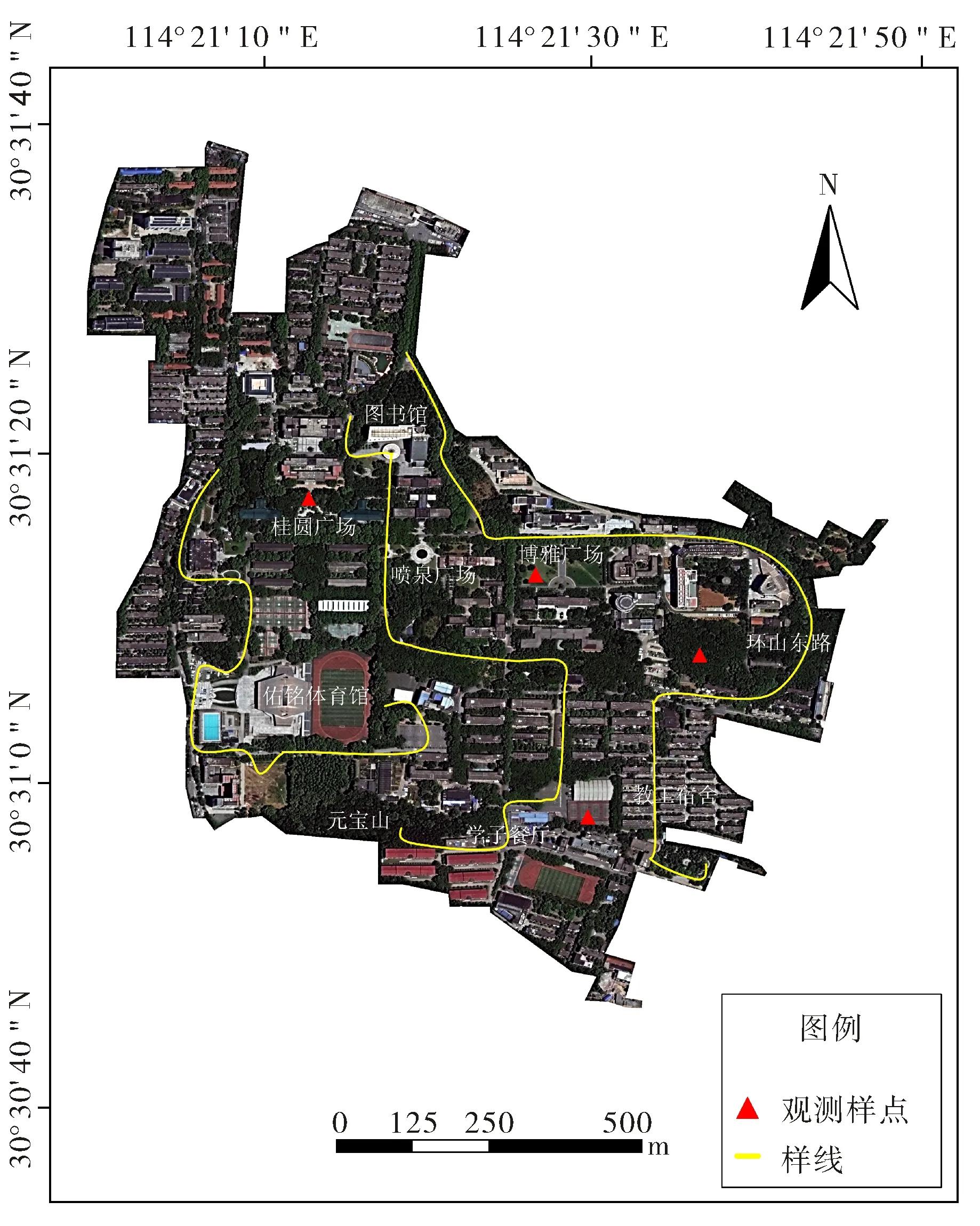
图1 桂子山鸟类调查样线及样点分布图Fig.1 Locations of line-transect and point-account of bird survey in Guizi Hill
自2017年9月至2018年8月,每月调查1次(11月和3月冬候鸟和夏候鸟陆续来武汉,每月各调查2次).调查选取晴朗无大风日期6∶30~11∶00及15∶30~18∶00鸟类活动高峰时段进行,用双筒望远镜(SAKURA,10×42)观察,通过直接观察、鸣声及活动痕迹等辨别种类,记录观察日期、时间、天气以及鸟类种类、数量、栖息生境及行为习性等信息.种类鉴别参考《中国鸟类野外手册》[9]和《BIRDS OF CHINA》[10];分类、学名和居留类型的确定依据《中国鸟类分类与分布名录(第三版)》[11];区系类型的确定则依据《中国动物地理》[6],濒危状况依据《中国脊椎动物红色名录》等[12].
1.3 统计方法
根据记录到的信息,通过下述公式进行分析[2,4].
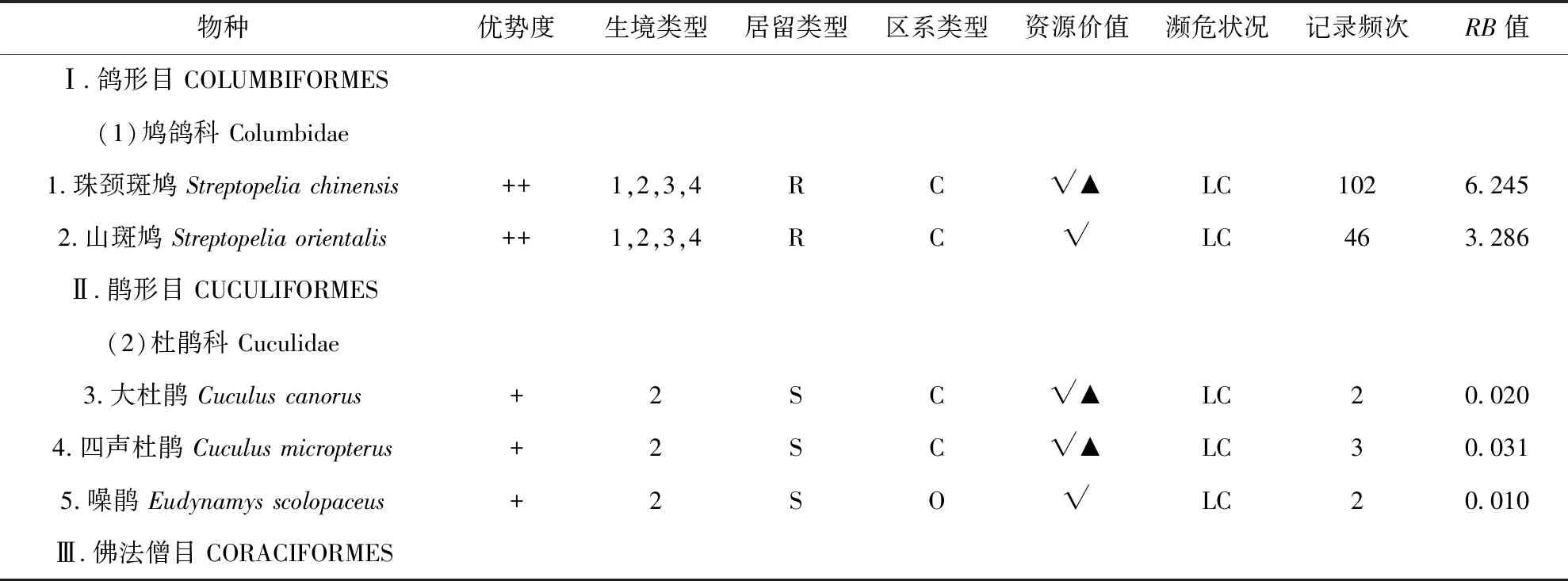
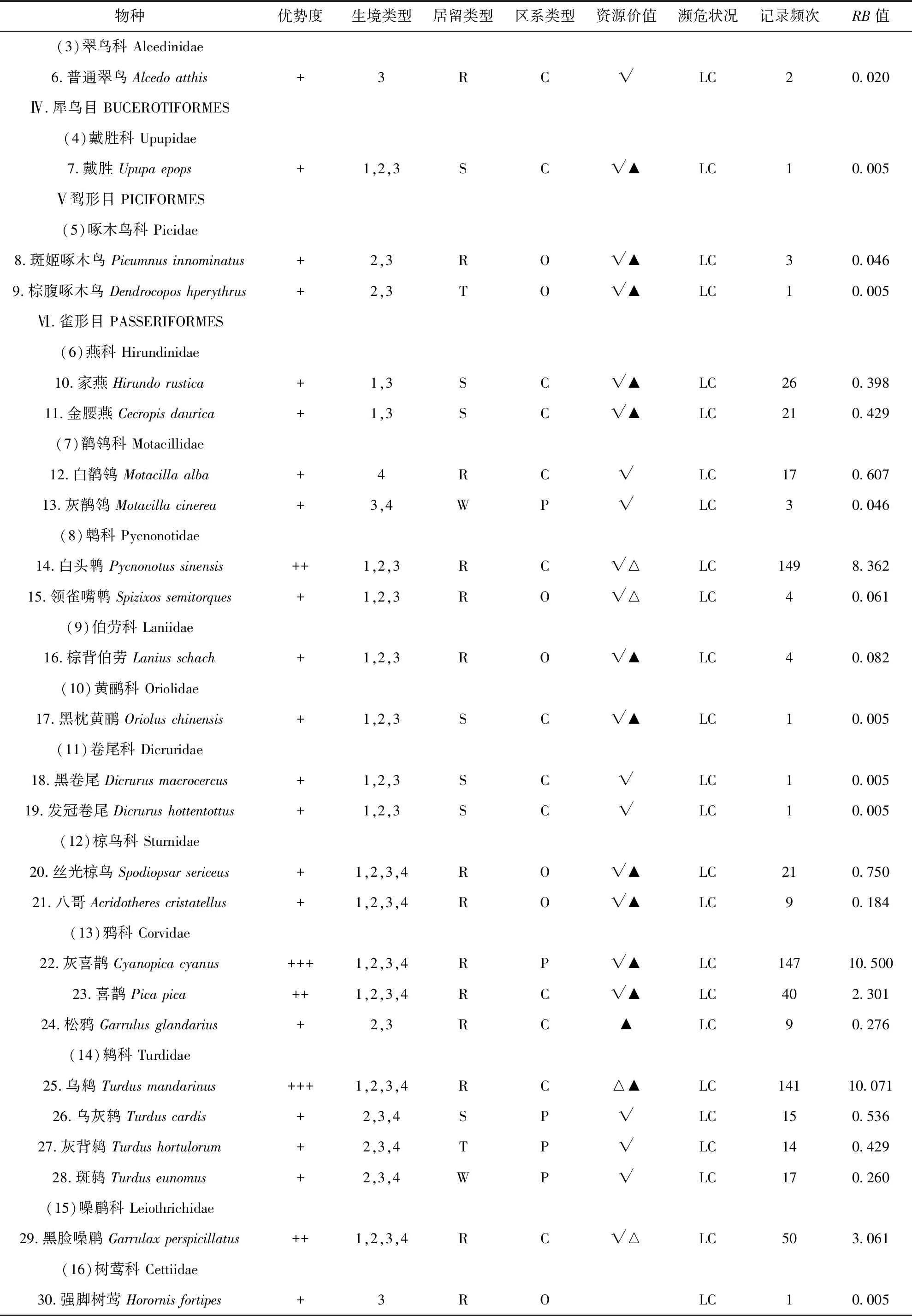
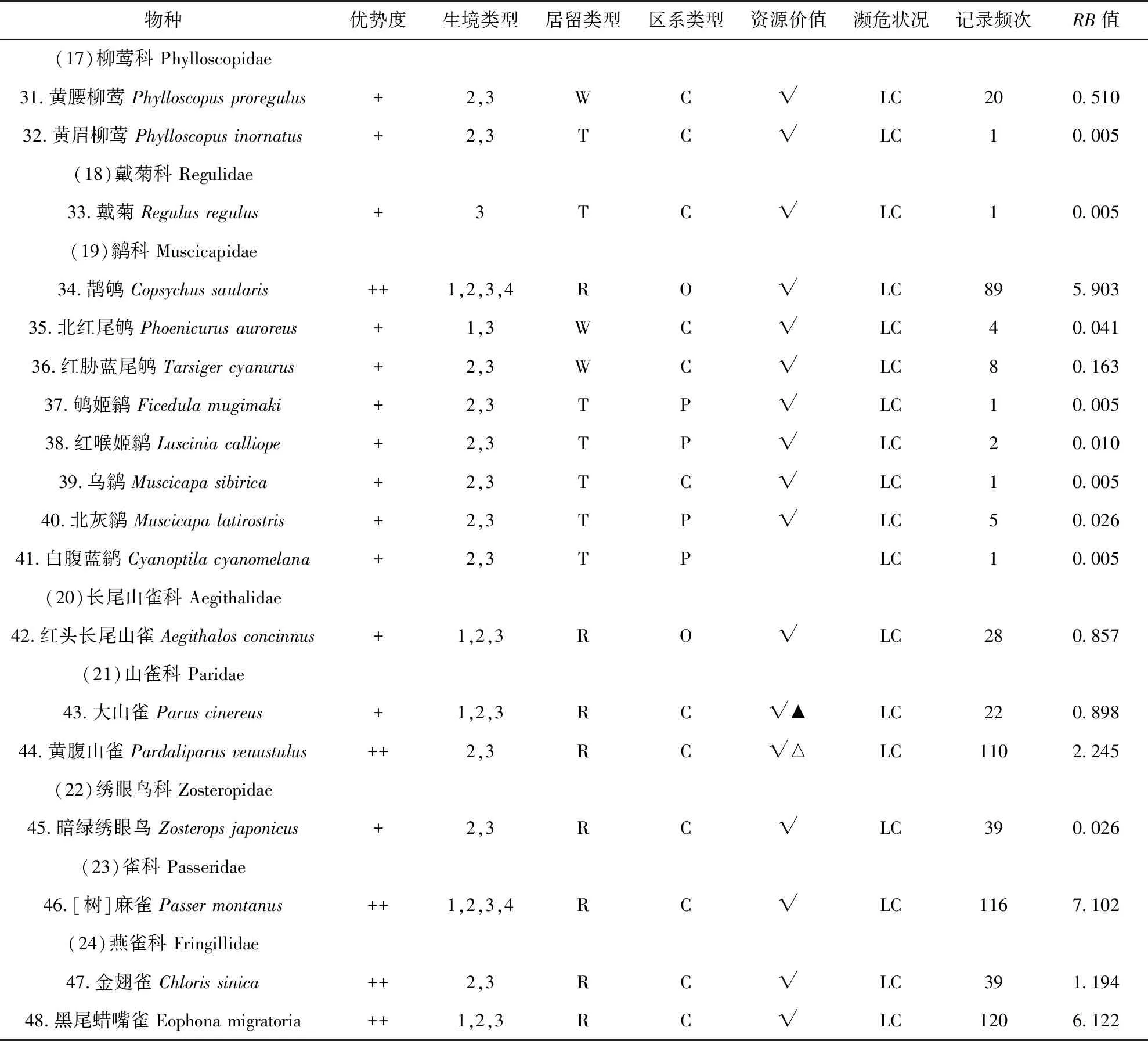
1) 频率指数公式:P=RB,其中P为频率指数;R为记录到该鸟的天数/调查的总天数;B为记录到该鸟的总数/调查的总天数.当RB>10时是优势种;1 3)Pielou物种均匀度指数 (E),公式:E=H′/Hmax=H′/lnS; 本次调查一共有249次有效记录,记录到鸟类48种,1 427只,分别隶属于6目24科 (表1),其中雀形目鸟类有19科39种,非雀形目鸟类有5科9种.种类最多的科是鹟科,有8种.优势种为乌鸫(RB=10.071)、灰喜鹊(RB=10.500)2种;常见种为白头鹎(Pycnonotussinensis) (RB=8.362)、[树]麻雀(RB=7.102)、珠颈斑鸠(Streptopeliachinensis) (RB=6.245)、黑尾蜡嘴雀(Eophonamigratoria) (RB=6.122)、鹊鸲(Copsychussaularis) (RB=5.903)、山斑鸠(Streptopeliaorientalis) (RB=3.286)、黑脸噪鹛(Garrulaxperspicillatus) (RB=3.061)、喜鹊(Picapica) (RB=2.301)、黄腹山雀(Pardaliparusvenustulus) (RB=2.245)、金翅雀(Chlorissinica) (RB=1.194)等10种;其余36种为少见种.在记录的48种鸟类中,中国特有种4种,即白头鹎、领雀嘴鹎(Spizixossemitorques)、黑脸噪鹛、黄腹山雀、乌鸫.有《国家保护的有益的或者有重要经济、科学研究价值的陆生野生动物名录》[13](简称《三有名录》)鸟类44种,占91.67%.有《湖北省重点保护陆生野生动物名录》[14]鸟类17种,占35.42%. 本次调查记录的鸟类含留鸟24种,占50.00%;夏候鸟10种占20.83%;冬候鸟5种占10.42%;旅鸟9种,占18.75%.留鸟种类、数量占比均最大;夏候鸟次之,以杜鹃科、燕科的鸟类为主;因地处地理区系的华中过渡地区,冬候鸟种群多样性不高,并伴随有数种旅鸟在该地区停留歇息. 本次调查记录的鸟类有广布种29种,占60.42%;东洋界鸟类10种,占20.83%;古北界鸟类9种,占18.75%.武汉在动物地理区划上,属于东洋界中印亚界华中区(Ⅵ)东部丘陵平原亚区(ⅥA)[6],地处古北界和东洋界分界线稍偏南,区系成分较复杂[4].既有北方渗入的一些古北种,如鹡鸰科、鸫科、鹟科的一些种类,也有华中、华南地区共有的一些东洋种,如鹎科、椋鸟科的一些种类,同时还有一些广布种的鸟类作为留鸟或夏候鸟在该地区栖息,如[树]麻雀、大杜鹃(Cuculuscanorus)等.华中区动物区系是华南区的贫乏化,由华南区向华中区,热带成分明显减少,典型类群在本地仅为华南区的一半.而与华北区的共有种大多为广泛分布于我国东部的北方成分[6].本次调查中,广布种最多,占60.42%,它们的繁殖区在华北和华中华南都有分布,如白头鹎.东洋界鸟类仅占20.83%,东洋种较少且不具有地区特有性.证实了鸟类在此处于古北界与东洋界的过渡状态. 表1 桂子山鸟类群落组成名录Tab.1 Birds in Guizi Hill 续表1 续表1 注:R=留鸟,S=夏候鸟,W=冬候鸟,T=旅鸟;P=古北种,O=东洋种,C=广布种;1=建筑及周边生境,2=乔木林生境,3=乔灌混合林生境,4:草坪生境;√=在《三有名录》中,▲=湖北省重点保护鸟类,△=中国特有种;LC=无危;RB:频度指数 Notes:R=resident,S=summer migrant,W=winter migrant,T=transient migrant,P=palearctic region,O=oriental region,C=cosmopolitan;1=building area,2=arbor forest,3= mixed forest,4=grassland;√=List of terrestrial wildlife that are beneficial to the state or have important economic and scientific value,▲=key protected terrestrial wild animals in Hubei Province,△=endemic species in China;LC=Least Concern;RB=frequency index 建筑及周边区域一共记录到鸟类22种1 144只,多样性指数为2.576 9,物种均匀度指数为0.833 7,物种优势度指数为0.486 9(表2),无只存在于该生境的鸟类,常见种类有[树]麻雀、鹊鸲、灰喜鹊等. 表2 桂子山不同生境鸟类群落特征比较Tab.2 Characteristics of bird community in different habitats in Guizi Hill 乔木林内共记录到鸟类39种1 352只,多样性指数为2.872 8,均匀度指数为0.784 2,物种优势度指数为0.457 3,常见种类有珠颈斑鸠、山斑鸠、乌鸫、灰喜鹊等,常集群活动,在林间或林下飞行、觅食、鸣叫等;仅出现在乔木林中的鸟类有大杜鹃、四声杜鹃(Cuculusmicropterus)、噪鹃(Eudynamysscolopaceus)(表2). 乔灌混合林区域一共记录到鸟类43种1 403只,多样性指数为2.958 5,物种多样性在4种生境中最高,物种均匀度指数为0.786 6,物种优势度指数为0.446 8(表2).有3种鸟类仅在该生境被记录,即普通翠鸟(Alcedoatthis)、戴菊(Regulusregulus)和强脚树莺(Horornisfortipes).该生境的常见种类有乌鸫、灰喜鹊、白头鹎、黑脸噪鹛、黑尾蜡嘴雀、丝光椋鸟等. 草坪区域一共记录到鸟类15种828只,多样性指数为2.324 2,物种多样性在4种生境中最低,物种均匀度指数为0.858 3,物种优势度指数为0.495 8(表2).仅出现在草坪的鸟类白鹡鸰(Motacillaalba),常成对或3只一起活动,于清晨人少时在草坪上跳跃觅食.该生境的其它常见种类有灰喜鹊、乌鸫等. 1953年-1965年,在桂子山共记录到鸟类127种,1966年-1990年共记录到113种,除去游禽、涉禽,1953年-1965年记录到81种,1966年-1990年记录到85种[3],本次调查为48种.过去30年中,桂子山的鸟类种类明显减少,由85种减少为48种,减少了43.53%.其中,鸡形目鸟类不再分布于该地区,隼形目、鸮形目、雀形目中的百灵科、鹡鸰科鹨属、蝗莺科、太平鸟科、鸦科鸦属、鹀科鸟类本次调查均未见.新纪录到的鸟类有10种,分属3目9科,包括鹃形目的噪鹃、形目的棕腹啄木鸟(Dendrocoposhperythrus)、斑姬啄木鸟 (Picumnusinnominatus)、雀形目的领雀嘴鹎、黑脸噪鹛、鹊鸲等.校园内的优势种由[树]麻雀、家燕逐渐变为现在的灰喜鹊和乌鸫. 2005年11月年-2006年3月,在桂子山记录到鸟类44种[4],除去游禽、涉禽,有35种.本次调查同时期记录到鸟类37种.与十年前相比,桂子山冬季的鸟类种数略有增加,增幅5.71%,主要表现在鸫科、鹟科的鸟类来此越冬或途径停留的种类增加了,如斑鸫(Turduseunomus)、鸲姬鹟(Ficedulamugimaki)等. 对本次调查的鸟类种群结构的分析,可以发现,鸟类多样性下降,同时,本次调查所观察到的鸟类中,有36种为少见种,占75.00%,说明桂子山鸟类群落优势种明显,多数种类数量较少.对校园生境的调查分析可以看出,各生境间异质性较低.各生境中,建筑及周边人工开发度高,人类活动频繁,因此多样性较低.不过,由于人类产生的生活垃圾成为一些鸟类的重要食物来源,使得部分种类鸟类经常关顾食堂、垃圾站等地,并就近栖息,从而得以维持较大的种群数量,典型的有[树]麻雀、鹊鸲、灰喜鹊等.乔木林由于该生境植被类型多样性较低且面积狭小,因此制约了该生境内的物种数目和种群数量.乔灌混合林则因形成了乔木层和灌木层的梯度分布,植被茂密,食物多样,利于鸟类藏匿、栖息和繁殖且该生境面积较大,因此造成了在该生境中记录到的鸟类种数和总个数最多,多样性指数最大.而草坪因为植被低矮,食物较单一,面积较小,且受人类活动影响较大,使在该生境记录到的鸟类种数和总个数最少,多样性指数最低.优势种在2个生境中均有分布.依据生态位理论,生境异质性下降,必然导致物种减少和多样性丧失[15],本次调查也印证了这一点. 与之前报道相比,桂子山鸟类群落发生较大变化,鸟类物种数和数量减少.其主要表现和原因如下. 1) 栖息环境改变,生境异质性降低.1953年-1990年,由于华中师范大学的迁入和周边地区的城镇建设,桂子山地区人类活动突然加剧,较大程度地改变了原始环境,一些对环境变化比较敏感,对原环境比较依赖,对人类活动产生的噪音等容忍性较差的种类,诸如环颈雉华东亚种(P.c.torquatus),鹌鹑(Coturnixjaponica),棕扇尾莺(Cisticolajuncidis)等长期栖息于此的留鸟或候鸟,由于本地区食物、环境及植被的改变,人类干扰的激增,迁出了桂子山地区.另一些如小蝗莺(Locustellacerthiola),原是本地区可见的旅鸟,由于旅鸟的迁徙路线由周围环境所决定,若环境变化,则它们的路线会不可避免地受影响,转而选取更适宜的环境停歇.随着小蝗莺依赖的近水灌丛的逐步消失,它们也不再出现于本地区.相反地,由于同时期本地区进行了大量植树,因此吸引了许多不甚畏人的食谷鸟、食虫鸟、杂食鸟,诸如鸫科、鹟科等类群的增加.1990年-2018年,随着人口和城市建设的继续增加,“农田”这一生境类型消失,其他几种生境如乔木林、乔灌混合林、草地也有同质化的趋势.就桂子山及周边的土地利用情况而言,耕地和湿地由自1988年-2015年减少了205.7 hm2,且主要转换为建设用地[16].在桂子山上,校舍建筑总面积由1991年约300 000 m2[17]增加为2014年的923 359 m2[18],增长3.08倍,且近年来还在不断增加,如2016年建成的文科教学科研综合楼(10 042 m2)等(建设前为低矮平房及绿地).桂子山地区由上世纪五六十年代的城郊慢慢被扩展进中心城区,大量原本适合鸟类栖息的地貌环境被改变,鸟类栖息地面积减少,栖息地碎片化和退化,生境多样性被较大程度地削弱.这就造成了依赖相应栖息地和食物的鸟类种群离开.以雀形目中鹡鸰科鹨属(Anthus)、鹀科(Emberizidae)为例,这两个类群在上世纪种群数量波动不大,在1953年-1965年,前者被记录到4种,后者7种;到了1966年-1990年,前者被记录到3种,后者8种.然而到了2005年-2006年,鹨属未被记录到,鹀科仅记录到2种,本次调查(2017年-2018年)两类都未见.鹨属的鸟类的栖息环境为平原、灌木丘陵、河谷阶地,筑巢于林缘和林中空地[10].鹀科的鸟类,栖息觅食于干燥灌草丛及林缘裸岩[19],食性以食谷为主,喜食草籽、黍米、高粱等[10].而目前桂子山地区被居民区和商业区包围,内部生境同质化,人为干扰增多,已没有适宜的生境.因此不难看出,这两个类群在桂子山的消失,一个重要的原因是适宜生境、食物的消失.另一个受栖息地及食物变化影响较大的是燕科的类群,以家燕为例,作为桂子山地区1953年-1990年的优势种,当前种群数量大幅下降,变为少见种.其原因有多方面,一是九十年代后建筑材料改变,外墙立面常采用密封的石材、弹性涂料(树脂)、玻璃、瓷砖等材料取代了过去的泥砖、石灰墙,屋檐也变小或消失,导致家燕难以营巢.二是由于农田等生境的消失,燕子的捕食受到影响,它们难以在林中捕虫.相反的,由于上世纪中叶及后来种植的乔木长势良好,树高及盖度都大幅增加,因此也吸引了鹟科等喜密林的鸟类,同时也造成了杜鹃科、鸫科、鹟科等类群的种群稳定,一些种类如乌鸫则成为该地区的优势种.而由于部分种植树木的非本土化和低多样性,也一定程度上造成了灰喜鹊种群的繁盛. 2) 人为噪声及活动的干扰.人为干扰也是影响鸟类群落结构的重要因子[23].华中师范大学在校生人数由1991年的12 000人[17]逐年增加为2014年的39 371人[18],增长3.28倍.随着城市的发展和人口数量的增多,桂子山内部及周边的人类活动和城建产生了较大的噪音,环境噪音对鸟类鸣声的频率和振幅的影响十分显著,可直接干扰部分鸟类的栖息[20].城市化似乎为鸟类设置了许多障碍,诸如噪音、污染、人类活动干扰等,只有那些在生物学上具有相应适应特性的种类才能忍受[21],如乌鸫、鹊鸲、灰喜鹊、喜鹊等.而一些林鸟则难以适应从而不再分布于本地区,如鹨属,它们普遍性机警,繁殖期受惊吓易弃巢. 栖息环境的破坏和人类活动等原因造成了鸟类多样性降低,而鸟类多样性是否丰富是反映良好生态环境的重要指标.研究表明,鸟类对栖息地的选择主要体现在栖息地结构间分布的异质性以及鸟类物种与栖息地结构的密切关系,而不是栖息地的面积[22].而在城市绿地植被结构中,乔木层结构复杂度与鸟类丰富度和多样性呈显著正相关 ;灌木层盖度和多样性也会显著提高鸟类,特别是林鸟的多样性水平[23];地被层的作用同样重要,其中的草本植物及小动物是鸟类食物的重要来源之一.因此根据现状,对桂子山地区的鸟类资源保护提出如下建议:①园林建设时树种应多样,避免单一的植被类型,且尽量使用本土植物;②在景观设计时应注意乔木灌木相结合,并通过合理规划植被丰富边缘效应,在地被上,建议减少地面硬化,多样化种植草本植物;③尽量保护现存的次生林,不搞大开发,保障现有鸟类的栖息条件;④有研究表明与没有植被的屋顶和墙壁相比,大型建筑上的绿色植物(屋顶、花园和绿色墙)支持了更丰富的鸟类种群[24].因此在学校建设过程中,可适量增加屋顶花园和绿色墙来作为生态走廊;⑤加强监管和教育,树立生态保护意识. 致谢感谢李菲凡、华中师范大学生命科学学院春野协会观鸟组对本次调查的帮助和部分数据的支持.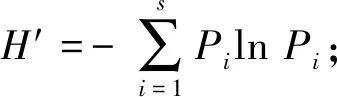

2 调查结果及分析
2.1 种类组成及区系分析
2.2 水平空间结构

2.3 群落动态
3 讨论
