水稻成花素基因RFT1对抽穗期的促进作用
2020-06-12王世林张振华朱玉君樊叶杨庄杰云
王世林 张振华 朱玉君 樊叶杨 庄杰云
(中国水稻研究所/水稻生物学国家重点实验室/国家水稻改良中心,杭州310006;第一作者:15621566500@163.com;*通讯作者:zhuangjieyun@caas.cn)
南方双季稻区是我国水稻的主要产区之一。双季稻种植模式有利于充分利用光温资源,增加水稻复种指数,从而提高水稻产出能力,保障农民的经济收入[1];同时,双季稻种植模式能够降低由自然灾害所引起的减产风险[2]。自2014 年以来,我国水稻种植面积已连续5 年减少[3-4];同时,我国水稻生产进入效益优先阶段,单位面积的高效率产出将是今后的重要生产目标[5]。进一步缩短早稻品种的生育期,能够为后茬晚稻提供更充足的生育时间,优化晚稻产出,从而提高双季稻的生产潜能。因此,挖掘能够促进早稻品种抽穗,同时不会导致稻谷产量大幅度下降的基因,对我国南方双季稻区的粮食安全生产具有重要意义。
近20 年来,水稻抽穗期的遗传和分子机理研究取得巨大进步,其主要调控网络已基本明确[6]。在获得克隆的水稻抽穗期基因中,有7 个在调控网络中居关键地位,分别是 Hd3a、RFT1、Hd1、Hd2、Ghd7、DTH8 和Ehd1。Hd3a 和RFT1 是水稻的2 个成花素基因,前者主要在短日照条件下发挥作用,后者主要在长日照条件下发挥作用,其他抽穗期基因通过调控Hd3a 和RFT1 影响抽穗[7-8];Hd1 和 Ehd1 分别是水稻抽穗期 2条主要调控通路的核心基因,前者在长、短日照条件下作用差异很大,后者的功能型等位基因在长、短日照条件下均促进抽穗[9-10];Hd2、Ghd7 和 DTH8 通过与 Hd1和Ehd1 的相互作用影响抽穗[11-14]。在我国长江中下游稻区和华南稻区中,早籼品种生长于长日照条件下均携带Ehd1 的早熟功能型等位基因;在Hd1 和Ghd7 座位上,该类品种均拥有早熟型组合,不携带功能型等位基因或仅在1 个座位携带功能型等位基因;在Hd2 和DTH8 座位上,绝大部分早籼品种携带不延迟抽穗的功能缺失型等位基因[14-15]。
前期研究表明,在其他抽穗期基因不分离的背景下,RFT1 本身的等位变异对水稻抽穗期和产量性状具有重要影响。与来源于早籼品种珍汕97 的等位基因相比,来源于中籼品种密阳46 的等位基因具有促进抽穗、降低产量的作用。当遗传背景中抽穗期基因呈珍汕97 纯合型和密阳46 纯合型自然混合时,密阳46 型RFT1 等位基因在长日照条件下促进抽穗18.4~31.2 d,同时对产量具多效性[16];当遗传背景中Hd1 座位固定为珍汕97 纯合型时,RFT1 的作用大幅度减弱[17]。本研究以珍汕97 和密阳46 为双亲构建新群体,进一步固定RFT1 之外6 个关键抽穗期基因的作用,在早籼遗传背景和自然长日照种植条件下,分析RFT1 对抽穗期和产量性状的遗传作用,探讨利用RFT1 进一步缩短早稻抽穗期的途径。
1 材料与方法
1.1 水稻材料
本研究所用水稻材料为衍生于珍汕97//珍汕97/密阳46 组合的2 个分离群体。在这2 个群体中,RFT1均呈分离,而其他6 个关键抽穗期基因(Hd3a、Hd1、Hd2、Ghd7、DTH8 和 Ehd1)均无功能性变化。其构建过程如图1:从珍汕97/密阳46 的F9群体挑选出1 个单株,与珍汕97 回交,连续自交加代至BC1F4,回交、自交过程中所挑选单株均为倾向于早熟的材料;经标记鉴定,筛选到在RFT1 位点座位上呈杂合的BC1F4单株2个,自交获得世代为BC1F5的近等基因系-F2(NIL-F2)群体,共含543 个单株,称之为Z3;经标记鉴定,从该群体挑选RFT1 纯合型单株,自交构建了1 个含有46个珍汕97 纯合型株系和55 个密阳46 纯合型株系的NIL 群体,称之为 R3。

表1 本研究所用群体及其亲本在7 个关键抽穗期基因上的功能型

图1 材料构建过程
1.2 田间试验与性状考查
所有群体均种植于中国水稻研究所试验基地(杭州富阳),种植季节均属长日照条件,其中,NIL-F2群体Z3 种植于 2017 年,5 月 24 日播种,6 月 16 日移栽;NIL 群体 R3 种植 2 年,2018 年于 4 月 18 日播种,5 月15 日移栽,2019 年于 4 月 18 日播种,5 月 16 日移栽。株行距为16.7 cm×26.7 cm,正常大田管理。对于NILF2群体Z3,记载各单株抽穗期,不考种。对于NIL 群体R3,采用完全随机区组设计,2 次重复,每重复每株系种植1 行10 株,记载各单株抽穗期,取平均值进行数据分析;成熟期每株系混收中间5 株,考查单株穗数、每穗总粒数、每穗实粒数、千粒重和单株产量。千粒重考查遵循ZHANG 等[18]方法。
1.3 抽穗期基因分析与标记检测
采用实验室保存的珍汕97 和密阳46 序列资料,分析双亲在7 个抽穗期基因上的功能型及双亲差异;根据双亲差异设计引物,检测群体。水稻材料移栽10 d后,每个单株取约2 cm 幼叶,采用ZHENG 等[19]方法提取DNA。PCR 扩增按照CHEN 等[20]方法进行,扩增产物使用6%非变性聚丙烯酰胺凝胶或2%琼脂糖凝胶电泳进行分离,分别使用银染和GelRed 检测。
1.4 数据分析
对于NIL-F2群体Z3,应用Windows QTL Cartographer 2.5 软件[21]中单标记作图法进行QTL 分析,以LOD=3.0 为阈值。对于NIL 群体R3,应用SAS 软件一般线性模型(Proc GLM)[22],采用双因素方差分析法检验2 种基因型之间的表型差异,具体方法同DAI 等[23]所述;若差异达到显著水平(P<0.05),则计算加性效应和贡献率。
2 结果与分析
2.1 群体遗传背景
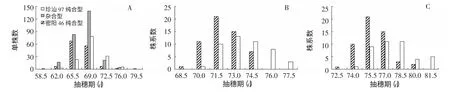
图2 NIL-F2 和NIL 群体的抽穗期表型分布

图3 NIL 群体的产量性状表型分布
根据珍汕97 和密阳46 的基因组序列,确定了它们在7 个目标基因座位上的功能型及差异情况(表1)。有3 个基因在2 个亲本之间无功能差异,其中,Hd3a 和Ehd1 为功能型等位基因,DTH8 为功能缺失型等位基因;根据这3 个基因对水稻抽穗期的作用,双亲所携带的等位基因均属早熟型。在其余4 个基因上,双亲之间存在功能差异:在RFT1 上,珍汕97 携带迟熟型等位基因,密阳46 携带早熟型等位基因;在Hd1 上,珍汕97 为强感光型,密阳46 为无感光型;在Hd2 上,珍汕97 为无感光型,密阳46 为强感光型;在Ghd7 上,珍汕97 为无功能型,密阳46 为强功能型。经检测构建分离群体的2 个BC1F4单株,发现这2 份材料除了RFT1 为杂合型外,其余3 个基因均为珍汕97 纯合型,即强感光型Hd1、无感光型Hd2 和无功能型Ghd7。无感光型Hd2 和无功能型Ghd7 丧失了在长日照条件下延长抽穗的作用,而强感光型Hd1 在无功能型Ghd7背景下具有促进抽穗的作用[12-13,24-25]。因此,Z3 和 R3 群体中Hd1、Hd2 和Ghd7 的基因型组合亦为早熟型。综上所述,本研究所用群体RFT1 呈分离,其余6 个关键抽穗期基因均与珍汕97 功能型一致,为早熟型遗传背景。
2.2 表型变异
在Z3 群体1 年试验和R3 群体2 年试验中,抽穗期均呈连续分布。以RFT1 基因型对群体内材料分类,可以发现,与珍汕97 纯合型材料相比,密阳46 纯合型材料趋向于早熟(图2)。该结果表明,在Z3 和R3 群体中,成花素基因RFT1 可能对抽穗期的变异具有显著作用。在考查了产量性状的R3 群体2 年试验中,各性状表型亦呈连续分布,且密阳型株系趋向分布于低值区(图3),表明RFT1 在这些试验中表现出对产量性状的多效作用。
2.3 QTL分析
NIL-F2群体中,Windows QTL Cartographer 2.5 结果显示,RFT1 对抽穗期呈显著作用(LOD=13.05),来自密阳46 的等位基因促进提早抽穗1.21 d,显性效应为0.62 d,贡献率为10.46﹪。
双因子方差分析结果显示,在NIL 群体R3 中,RFT1 表现出对抽穗期、每穗总粒数、每穗实粒数、千粒重和单株产量的显著作用,而对单株穗数无显著作用(表2)。2018 年,密阳46 等位基因促进抽穗,加性效应为0.84 d,贡献率为18.62%;同时,密阳46 等位基因降低每穗总粒数、每穗实粒数、千粒重和单株产量,加性效应分别为 3.06 粒、2.98 粒、0.12 g 和 0.63 g,贡献率分别为1.66%、2.13%、2.53%和2.76%。2019 年,密阳46等位基因亦促进抽穗并降低每穗总粒数、每穗实粒数和单株产量,加性效应分别为0.89 d、2.02 粒、1.48 粒和 0.38 g,贡献率分别为 18.48%、1.91%、0.99%和1.48%;在千粒重上处于临界水平,P 值为0.0652,加性效应和贡献率分别为0.08 g 和1.28%。综上结果表明,在早熟遗传背景和自然长日照种植条件下,水稻成花素基因RFT1 不仅对抽穗期具有显著作用,同时对产量性状呈多效作用。

表2 近等基因系群体中RFT1 对抽穗期和产量性状的作用
3 讨论与结论
本研究应用早籼稻珍汕97 与中籼稻密阳46 为双亲材料,针对 Hd3a、RFT1、Hd1、Hd2、Ghd7、DTH8 和 E-hd1 这7 个控制水稻抽穗期的关键基因,构建了RFT1分离、其余6 个基因均为早熟基因型的群体,分析了RFT1 对抽穗期和产量性状的遗传作用。结果表明,在早熟型遗传背景和自然长日照种植条件下,与早籼稻珍汕97 相比,中籼稻密阳46 的RFT1 等位基因能够稳定促进水稻抽穗,同时对产量性状具有微效作用。
抽穗期属于数量性状,由主效和微效数量性状座位(QTL)共同控制。主效QTL 主要影响水稻品种的地区和季节适应性;而微效QTL 在相同或相似的生态适应区域内通过微调抽穗期,在充分利用自然资源或规避逆境胁迫中发挥关键作用[26]。近年来,水稻抽穗期微效QTL 的研究逐渐受到重视,迄今有4 个微效QTL 获得克隆、3 个QTL 得到精细定位[26-32]。此外,一些已克隆的主效QTL 在特定的遗传背景或环境条件下,也呈现出微效作用。例如,TAKEUCHI 等[33]利用分子标记辅助选择,将水稻品种Kasalath 的Hd4/Ghd7 等位基因导入另一品种Koshihikari 中,使其抽穗期延长约3 d,同时增加了穗长和单株产量;ZHAO 等[34]以 T65 和 GLA 为双亲构建的新材料中,在抽穗期基因组合为Hd3a、RFT1、Hd1 和 ghd7 的背景下,Ehd1-GLA 在长日照条件下促进水稻抽穗约5 d;FUJINO 等 [35]以早熟品种Hoshinoyume 为受体亲本,构建了 NIL(hd5/dth8),在长日照条件下较亲本延迟抽穗5.2 d。在本研究中,RFT1在早熟遗传背景和长日照种植条件下呈现出微效作用,3 年加性效应为0.84~1.21 d。我们可以将RFT1 的这一特性利用到早稻品种的抽穗期精准调控中,以最大化利用自然资源和规避自然灾害。综上表明,针对已克隆的主效QTL,构建特定的遗传背景材料来分析它们的微效作用,一方面可以获得新的微效QTL 资源,另一方面可为主效变微效分子机理研究提供基础。
进一步缩短早稻品种的生育期,为后茬晚稻提供更充裕的生长发育时间,是我国南方双季稻区一个重要的育种目标。本研究结果表明,密阳46 的RFT1 等位基因能够进一步缩短早稻的生育期,为晚稻提供更充裕的生长时间,进而提高双季稻的生产潜能。据浙江省品种审定条例,早籼稻参试品种全生育期比对照缩短1.0~3.0 d 的,产量要求每缩短1.0 d 可以比对照减产2.0%;缩短4.0 d 及以上的天数,每缩短1.0 d,可比对照减产3.0%。本研究2018 年和2019 年NIL 结果显示,中籼稻密阳46 型RFT1 分别促进水稻提早抽穗1.68 d 和 1.78 d,降低单株产量 1.26 g 和 0.76 g,符合早籼品种审定需求。由此可见,来源于其他品种类型的RFT1 等位基因,可为南方双季稻区特早熟水稻品种的培育提供新的资源。
