水稻土中铁氧化物对产甲烷古菌群落结构的影响
2020-06-11徐健鑫扆幸运李晓明丁龙君朱永官
徐健鑫,扆幸运,李晓明,丁龙君,朱永官,4,*
1 中国科学院生态环境研究中心土壤环境研究室, 北京 100085 2 中国科学院大学中丹学院, 北京 100049 3 中国科学院大学, 北京 100049 4 中国科学院城市环境研究所, 厦门 361021
甲烷(CH4)是继二氧化碳(CO2)之后第二大温室气体,其较强的红外线吸收能力使其单分子增温的潜力是CO2的20—30倍[1- 2]。目前全球每年CH4排放量约5×108—6×108t,其中约70%来源于微生物作用[3]。产甲烷菌是其中最大的贡献者。产甲烷菌是地球上最古老的生命形式之一[4],广泛分布于稻田、海洋沉积物、厌氧消化器等厌氧环境中[5- 7],是复杂有机质厌氧降解的重要推动者[8]。根据碳源差异,产甲烷菌的能量代谢方式可分为:以H2或甲酸为电子供体还原CO2、乙酸发酵及甲基营养型等三种[9]。
稻田是我国最重要的人工湿地生态系统,也是CH4排放的人为源之一[10]。每年来源于稻田的CH4排放量约2×107—1.2×108t,中国位于世界首位[11]。氧化铁是稻田土中最丰富的氧化物。多形态、高化学活性的氧化铁在周期性干湿交替的稻田土壤中表现出活跃的价态变化,影响着其他营养元素的生物地球化学循环[12-13]。有研究报道,由微生物介导的铁还原直接影响着甲烷合成等其他代谢途径[14]。Jackel等研究发现向水稻土中添加15或30 g/kg水铁矿时,甲烷释放的抑制效率分别可达43%和84%[15];Peng等向不同植区水稻土中添加葡萄糖后发现土壤中Fe(II)的产生量与CH4的释放量显著相关,并提出以Fe(II)/[Fe(II)+Fe(III)]预估CH4排放量的模型[16]。然而,目前关于铁氧化物对产甲烷菌群落结构的影响报道较少。
本研究通过泥浆厌氧培养,以甲酸盐为产甲烷底物,水铁矿为唯一电子受体,探究添加铁氧化物(铁还原)对产甲烷菌的群落组成的影响。本实验将为探究水稻土中铁还原与碳代谢间耦合的分子机制奠定基础。
1 材料与方法
1.1 水铁矿的制备
水铁矿(Fe5HO8·H2O[12])的制备方法参照Schwertmann和Cornell[17]进行了适当修改。用去离子水配制0.4 mol/L FeCl3溶液,并用1.0 mol/L NaOH溶液将pH调至7.0。然后用10倍体积的去离子水洗涤、离心沉淀(5000 r/min,10 min),直到上清液中Cl-浓度小于1 mmol/L。将洗好的沉淀冷冻干燥,研磨过筛(0.07 mm),室温保存。
1.2 无菌缺氧去离子水的制备
向去离子水中添加微量刃天青作为氧化还原指示剂,将无菌针头插入液面以下并通入高纯N2,加热并维持沸腾状态30 min,以去除去离子水中的溶解氧,此时液体成粉紫色;稍冷,加入0.3 g/L L-半胱氨酸,持续微热并通入高纯N2至液体变为无色;迅速封装,高压灭菌(121℃,20 min),冷却待用。
1.3 泥浆厌氧培养
土壤样品采自湖南省桃源县长期施肥实验站(28°21′N,116°92′E)。采集土壤表层0—20 cm,并去除样品中细小的沙石和根系,放入无菌自封袋中密封,冷藏运回实验室。将样品风干过筛(1 mm),充分混匀。

由于微生物对甲酸盐的大量消耗,在10 d培养期间每天向体系中按7 μmol/g的浓度补充甲酸盐,且培养结束时(记为第10天)体系中几乎没有甲酸盐剩余。
1.4 泥浆DNA提取及16S rRNA基因测序
取培养第0、10天的泥浆0.5 g(干土计),用FastDNA Spin Kit for Soil(MP bio,CA,USA)试剂盒提取泥浆 DNA。选择16S rRNA基因的V4高变区(ArBa515f_Arch806r)进行扩增,20 μL PCR体系包括:4 μL 5×FastPfu Buffer,2 μL 2.5 mmol/L dNTPs,前、后引物各0.8 μL,0.4 μL FastPfu Polymerase,0.2 μL BSA和2 μL DNA,剩余用无菌水补齐。PCR反应条件如表1。样品送至上海美吉生物医药科技有限公司进行测序。

表1 PCR反应引物和反应条件
1.5 数据分析
初始数据分析采用Excel 2016进行,采用SPSS(20.0,SPSS Inc.,Chicago,IL,USA)软件进行显著性分析(one-way ANOVA)。计算香农指数和辛普森指数描述古菌群落的α多样性,基于Weighted normalized Unifrac距离算法进行主坐标分析。将相对丰度发生显著差异的操作分类单元(operational taxonomic unit,OTU)序列与NCBI数据库中序列比对,利用MEGA的Neighbor-Joining法构建系统发育树,检验方法为bootstrap,重复1000次。
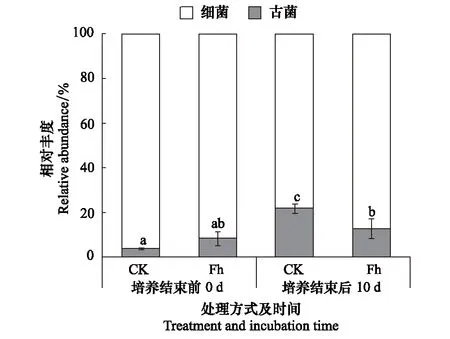
图1 培养结束前、后微生物群落组成 Fig.1 Barplot of microbial community structure in CK and Fh treatments at day 0 and 10 CK:对照处理 Control;Fh:水铁矿处理 Ferrihydrite.误差线为三组重复的标准偏差,上方的小写字母为不同处理培养前后的显著性差异(P<0.05)
2 结果
2.1 古菌群落多样性变化分析
基于16S rRNA基因测序结果,我们发现微生物群落大多由细菌构成,古菌占比不足30%,且培养结束后仅CK处理中古菌所占比例显著(P<0.05)升高,Fh处理中无显著变化(图1)。
进一步从OTU水平上分析CK和Fh两组处理中古菌群落组成的多样性,发现两组处理中香农指数均极显著下降(P<0.001),而辛普森指数极显著上升(P<0.001,图2)。这说明添加甲酸盐处理10天后,不仅降低了体系中古菌群落的物种多样性,同时降低了OTU水平上古菌群落的均一性。另外,发现与CK相比,添加水铁矿尽管可以降低微生物群落结构中古菌的占比,但对古菌群落组成的多样性没明显影响。基于Weighted normalized Unifrac算法的主坐标分析(图3)表明经10天厌氧培养,CK和Fh处理的古菌群落结构均与培养前明显不同,且两者培养后的古菌群落组成也具有显著差异;主坐标解释了总方差的82.84%。

图2 培养前、后泥浆中古菌群落结构的多样性指数(*** P<0.001)Fig.2 Barplot of Shannon index and Simpson index of archaea in CK and Fh treatments at day 0 and 10

图3 培养前、后泥浆中古菌OTU水平的主坐标分析图Fig.3 Principal co-ordinate analysis plot for archaea community structure in CK and Fh treatments at OTU level
2.2 OTUs水平上古菌群落结构变化分析
为进一步分析添加甲酸盐和水铁矿对古菌群落的影响,我们对比不同处理中培养前后每个OTU在体系中的相对丰度,最终发现OTU 2056,OTU 911,OTU 3870,OTU 2032,OTU 1970,OTU 2456,OTU 1931和OTU 895等8个培养后相对丰度发生显著(P<0.05)变化的OTU(图4)。其中,仅OTU 2056和OTU 911相对丰度升高,分别占CK和Fh处理古菌群落的80%和76%;根据系统发育分析,两者均与Methanobacterium具有较近的亲缘关系。OTU 2032和OTU 2456仅在CK处理中相对丰度下降,它们与Methanosarcina具有较高的相似性;而OTU 1931和OTU 895相对丰度变化仅发生于Fh处理中,与Methanocellales较为相似。

图4 厌氧培养10天后CK处理和Fh处理中相对丰度显著变化的OTUs及其系统发育分析(*P<0.05,*** P<0.01,*** P<0.001)Fig.4 Phylogenetic tree of OTUs whose relative abundance change significantly after 10-day anaerobic incubations in control and Fh treatment

图5 厌氧培养10天后Fh和CK处理中OTU差异的韦恩图 Fig.5 Venn diagram of OTU difference between CK and Fh after 10-day incubations
3 讨论
甲酸是生物大分子降解过程中除乙酸外又一重要的中间代谢产物[18],也是大多H2营养型产甲烷菌可以利用的一种底物[10]。从能量角度上,除直接还原CO2/CO途径以外,以甲酸盐为电子供体是微生物最容易产甲烷的途径[10]。本研究结果显示,以甲酸盐为底物进行泥浆厌氧培养后,体系的古菌群落结构从OTU水平上发生了明显的改变(图3),其物种多样性和群落结构的均一性均显著降低(P<0.05,图2)。造成这一结果的原因是添加甲酸盐刺激了OTU 2056和OTU 911的显著富集(P<0.001,图4),使其成为古菌中的主要优势种,从而降低了其他如Methanosarcina、Methanocellales和Crenarchaeote等类似古菌的相对丰度。系统发育分析结果显示这两个OTU均与Methanobacteriumbryantii有较高的相似性。M.bryantii在系统发育上与M.formicicum相似,两者都能利用甲酸盐产H2和CH4[19]。Bryant等最早发现M.bryantii可以利用S菌株氧化乙醇的产物H2合成甲烷,降低了H2累积对S菌株生长的抑制,从而实现了两者的互营共生[20]。之后,Guyot等也发现M.bryantii可以利用甲酸,与DesulfovibriovulgarisJJ共生产甲烷[21]。我们的体系中是否也存在着这种互营关系有待进一步确认。
水稻土中存在着丰富的铁氧化物,在周期性灌溉-排水的管理模式下,铁也发生了活跃的价态变化,并影响着其他生命活动[13,22]。结果发现,添加水铁矿培养后,古菌在全部微生物群落中的占比没有发生明显变化,显著低于CK处理(P<0.05)。这与早期的研究报道一致。微生物(细菌)介导的铁还原与产甲烷过程竞争电子和底物,因此与不发生铁还原相比,产甲烷菌的生长会受到抑制[5]。但我们进一步发现,CK和Fh处理培养前后古菌群落的α多样性没有差异,且培养后两个处理组中的优势种均为OTU 2056和OTU 911,占比也相似。这说明添加水铁矿对古菌群落的迁移没有主导作用。培养后83% OTUs同时存在于CK和Fh处理中(图5),也验证了这一结论。
4 结论
添加甲酸盐可以有效地富集土壤中Methanobacteriumbryantii的类似微生物;添加铁氧化物只能从总体上减少古菌占微生物总体的比重,但对古菌群落的多样性和均一度以及显著富集的菌株类型没有影响。因此,在有机物的厌氧降解过程中,铁氧化物可能只通过抑制产甲烷菌的生长减少甲烷的释放,而对主要发挥功能的菌株不具选择作用。
