珍稀濒危树种陆均松天然种群结构与空间分布格局
2020-06-11刘浩栋徐志扬吴春燕陈永富
刘浩栋,陈 巧,徐志扬,吴春燕,陈永富,*
1 中国林业科学研究院资源信息研究所,北京 100091 2 国家林业和草原局华东调查规划设计院,杭州 321000
种群作为物种和植物群落的基本单位,其结构和空间分布格局一直是生态学研究的核心问题[1]。种群结构反映不同年龄的种群个体在群落中的组配状况,其既能揭示种群历史动态特性与受干扰状况,也能预测未来动态及演变趋势[2- 4]。种群空间格局则体现种群个体在水平空间中的分布状况,是揭示种内种间关系、环境适应机制及空间异质性的重要属性[5- 6]。由于环境因素以及物种自身的特性,同一物种在不同发育阶段和不同分布区域的特征会有一定的差异[7]。因此,分析种群的空间异质性特点对于种群尤其是濒危种群的更新复壮及其所在群落和生态系统的生物多样性保护具有重要意义[8- 10]。目前,种群空间格局研究最为广泛的方法是大样地监测与点格局分析技术,该方法以大样地内连续的空间调查数据为基础,通过植物种个体的空间坐标分析各种尺度下的种群格局,克服了传统方法只能分析单一尺度的缺点[11- 13]。但珍惜濒危种群多为小密度或极小密度种群,数量稀少,分布零星,传统样地调查方法更为适用[14]。
陆均松(Dacrydiumpectinatum)是罗汉松科(Podocarpaceae)陆均松属(Dacrydium)植物,其现代分布中心在新西兰及其附近岛屿(该属计有23种);在北半球的现代分布约限制在20°N以南,我国仅有Dacrydiumpectinatum一种,仅分布在海南岛的霸王岭、尖峰岭、吊罗山、五指山等700—1300 m的山地雨林中[15-16]。海南岛的山地雨林主林层高度一般在18 m以上,而高度大于27 m的树种在形成林隙尤其是较大面积的林隙中具有重要作用,从而对森林的更新、结构形成和多样性具有重要意义[17]。据调查,含陆均松的植物群落通常高达20—30 m,构成了海南岛热带雨林的顶层群落;因此,陆均松在本地的山地雨林中占据重要地位,同时也是优势种和建群种之一[18]。探明天然陆均松种群结构动态及空间分布格局对研究海南岛热带山地雨林的区系特征及生物多样性保护具有重要的科学意义。但由于对环境变化的反应敏感加之自身的生物学特性、自然灾害和人为破坏等原因,导致其日趋衰亡而成为渐危种。尤其自20世纪60年代后,过度砍伐加剧,种群资源日益减少,林下幼苗更新困难,林分品质也越来越差,已严重影响其正常演变、生存和发展,现已被列入濒危保护植物名录[19]。目前有关其种群生态学的研究多涉及群落特征、种群遗传多样性、孢粉分析与起源演化、天然更新和种子雨、土壤种子库等方面[20- 22],近年来,陆均松种群结构和空间格局的研究也多有报道,如李肇晨等[18]对海南霸王岭陆均松种群进行了年龄结构及点格局特征分析,练琚蕍和余世孝[23]研究了海南岛霸王岭陆均松及线枝蒲桃区系的特点。但前人多涉及单一区域,少有从空间异质性角度分析不同分布区域的种群格局差异。基于此,本文选择海南省霸王岭、吊罗山、尖峰岭3种不同区域的陆均松种群作为研究对象,通过传统样地调查方法试图研究探讨以下问题:(1)陆均松天然群落乔木层物种多样性及优势种组成;(2)陆均松种群的年龄结构特征;(3)陆均松种群分布格局的空间异质性差异。旨在深入分析探讨陆均松种群的生存现状及其种群格局,以期为进一步研究陆均松种群濒危原因、并保护其天然群落生物多样性提供科学依据。
1 研究区域与研究方法
1.1 研究区域概况

图1 研究区域地理位置Fig.1 Research area location
实验地点选择陆均松天然群落主要分布区,包括海南岛霸王岭、吊罗山和尖峰岭国家森林公园(以下用BWL、DLS、JFL表示)(具体地理位置见图1)。
BWL位于海南岛西南部,地理坐标介于109°03′—109°17′ E,18°57′—19°11′ N之间,总面积72000 hm2,其中1980 年建立的长臂猿(Hylobatesconcorloris)保护区位于800 m 以上的山地,覆盖面积约2500 hm2。其地处热带季风气候区,年均温度23.6℃,年均降雨量1500—2000 mm,平均相对湿度65%—90%[24],地形以山地为主,植被类型以低山雨林、沟谷雨林和山地雨林为主,森林覆盖率97.3%。内有维管束植物220科967属2213种,属于国家一级保护的有海南苏铁(Cycashainanensis)、坡垒(Hopeahainanensis)2种,国家二级保护的有油丹(Alseodaphnehainanensis)、海南风吹楠(Horsfieldiahainanensis)、海南梧桐(Firmianahainanensis)、蝴蝶树(Heritieraparvifolia)、海南紫荆木(Madhucahainanensis)等17种。
DLS位于海南岛东南部,地理坐标介于109°41′38″—110°4′46″E、18°38′42″—18°50′22″N之间,总面积37900 hm2,属东亚海洋性热带季风气候,年均温度24.4℃,降雨量1870—2760 mm[25],总地势西北高东南低,森林植被类型为热带次生林、季雨林、常绿阔叶林,森林覆盖率达96.26%。截至2016年,区域内有维管植物239科959属2116种,大型真菌16目35科112属281种,优势种为海南杨桐(Adinandrahainanensis)、陆均松(Dacrydiumpectinatum)、海南鹅掌柴(Schefflerahainanensis)、钝齿木荷(Schimacrenata)、狗骨柴(Diplosporadubia)、华南毛柃(Euryaciliata)、五列木(Pentaphylaxeuryoides)、鸡毛松(Podocarpusimbricatus)等[26]。
JFL位于海南岛西南部,地理坐标介于108°46′—109°45′E、18°23′—18°50′N之间,总面积64000 hm2,属热带雨林气候区,年降水量1300—3700 mm,年平均气温24.5℃[27],植被类型包括热带半落叶季雨林、热带常绿季雨林、滨海有刺灌丛、稀树草原、山顶苔藓矮林6种景观,森林覆盖率高达98%。此地为中国生物多样性最高的地区之一,与全海南岛总物种数目相比,目前已知野生蕨类植物39科79属150种,占30.9%;裸子植物5科7属13种,占52.0%;被子植物189科902属2124种,62.0%(表1)。
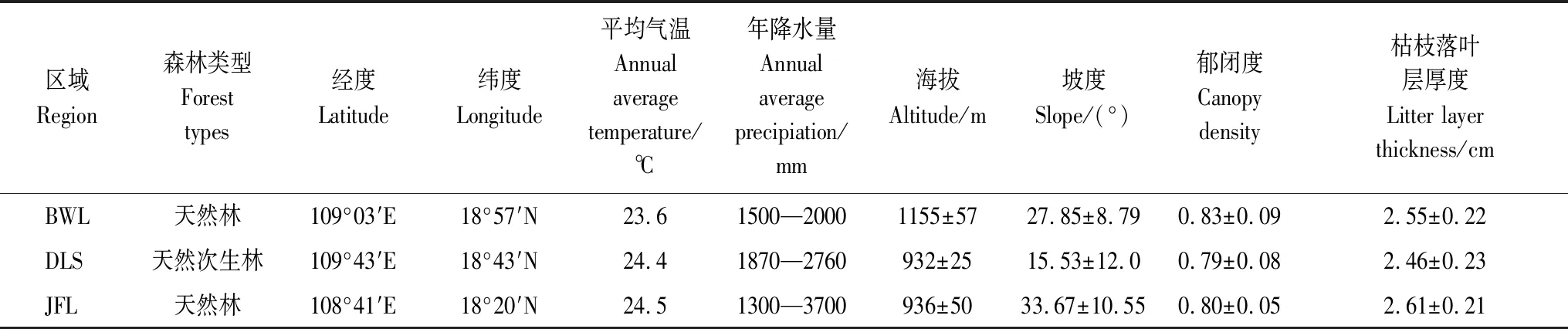
表1 陆均松天然群落分布区概况
BWL: 霸王岭国家森林公园Bawangling National Forest Park; DLS: 吊罗山国家森林公园 Diaoluoshan National Forest Park; JFL: 尖峰岭国家森林公园 Jianfengling National Forest Park; 表中所有数据以均值±标准差(Mean±SD)的方式表示
1.2 研究方法
1.2.1野外调查
沿用传统样地调查方法,布设面积为900 m2(30 m×30 m)的陆均松固定样地96个,BWL和JFL各30个,DLS为36个。因陆均松种群数量稀少,个体分散,故每个样地中至少包含1株及以上成年陆均松。使用相邻格子法把每个样地划分为36个5 m×5 m单元格,检尺单元格内所有DBH≥5 cm的乔木特征值(注:陆均松的调查标准为DBH≥1 cm),包括胸径、枝下高、树高、冠幅、相对位置等,并逐株挂牌;记录各样地海拔、经纬度、坡向、坡位、坡度、地形、林层数、郁闭度等指标。每个样地沿对角线于左下、中、右上设置3块2 m×2 m的灌木和草本样方,调查株丛数、种名、盖度、平均高度。乔木幼苗更新调查:每个样地内随机选择3块2 m×2 m乔木幼苗样方,记录DBH<5 cm的乔木种名、地径(H>1.3 m记录DBH)、高度、冠幅,并逐株挂牌。
1.2.2乔木层树种重要值
重要值(IV)=相对多度(%)+相对频度(%)+相对胸高断面积(%),范围为0—300%。
1.2.3乔木层物种多样性
(1)物种丰富度指数(S)= 样方内出现的物种数目
(2)Shannon-Wiener指数
(3)Simpson指数
(4)Pielou均匀度指数
Jsw=H′/lnS
式中,Pi=Ni/N,即某个物种的相对多度,Ni为种i的株数,N为种i所在样方所有物种的总株数[28- 29]。
1.2.4龄级结构划分标准
以立木个体径级代替年龄龄级分析种群年龄结构特点[18],根据陆均松生活史特征及实测数据,将其划分为12个龄级,DBH≤4 cm为第Ⅰ龄级,4
1.2.5空间分布格局
使用方差比均值(以下用V/M表示)的t检验法分析3处区域不同发育阶段的陆均松种群空间分布格局,并使用丛生指数(I)、平均拥挤度(M*)、Cassie指数(CA)、Lloyd聚块性指数(m*/m)、负二项式参数(K)共5种指标判断其聚集强度。
1.3 数据处理
所有数据均使用Excel 2013进行预处理,使用SPSS 20.0进行数理统计和分析,Origin 9.0软件作图。
2 结果与分析
2.1 陆均松天然群落乔木层物种多样性及优势种组成
物种多样性代表了群落稳定健康与否和种群资源丰富程度,本次调查(表2),在BWL、DLS、JFL记录了DBH≥5 cm的乔木层木本植物活个体数分别为5572、5506、4781株,平均种群密度表现为BWL(2063 stems/hm2)>JFL(1771 stems/hm2)>DLS(1699 stems/hm2);平均胸高断面积表现为JFL(65.85 m2/hm2)>BWL(61.53 m2/hm2)>DLS(37.58 m2/hm2)。物种丰富度(S)最高的是BWL,其次为JFL,DLS最低;Simpson指数(D)表现出相同的规律,即BWL>JFL>DLS;Shannon-Wiener(H′)最高的是JFL,具体表现为JFL>BWL>DLS;Pielou指数(jsw)DLS最高,呈DLS>BWL>JFL的趋势。3种陆均松群落乔木层结构及物种多样性指数均较高,之间差异较小,基本表现为BWL>JFL>DLS。表2列举了4种其他森林植被的多样性指标,通过比较,其S、H′等小于BWL、DLS、JFL;可见,陆均松天然群落物种多样性指数远高于中高纬度森林群落。
群落内物种组成是群落的主要特征,表3统计了3种陆均松天然群落乔木层重要值排前10的优势树种。除DLS的陆均松和鸡毛松的重要值相对较高之外,其他优势种呈现类似的规律,即重要值相对较小(1.44—8.68),少有占绝对优势地位的树种,符合海南热带山地雨林物种组成的一般规律。BWL优势种重要值排第1位的为陆均松;JFL则为白颜树,陆均松在JFL排第4位。海南岛热带山地雨林植物区系复杂,根据表2可知,BWL陆均松天然群落乔木层主要包括罗汉松科(Podocarpaceae)、壳斗科(Fagaceae)、桃金娘科(Myrtaceae)、樟科(Lauraceae)、木兰科(Magnoliaceae)、远志科(Polygalaceae)、冬青科(Aquifoliaceae)、山矾科(symplocaceae)等;DLS主要包括罗汉松科(Podocarpaceae)、茶科(Theaceae)、山矾科(symplocaceae)、五列木科(Pentaphylacaceae)、安息香科(Styracaceae)、五加科(Araliaceae)、大戟科(Euphorbiaceae)组成;而JFL则主要由榆科(Ulmaceae)、棕榈科(Palmae)、罗汉松科(Podocarpaceae)、樟科(Lauraceae)、无患子科(Sapindaceae)、茜草科(Rubiaceae)组成。

表2 陆均松天然群落乔木层结构及物种多样性与其他类型群落对比
BWL:霸王岭Bawangling;DLS:吊罗山Diaoluoshan;JFL:尖峰岭Jianfengling;DQS:大青山Daqingshan;THS:太行山Taihangshan;GDS:关帝山Guandishan;YLP:鹞落坪Yaoluoping;ALS:哀牢山Ailaoshan
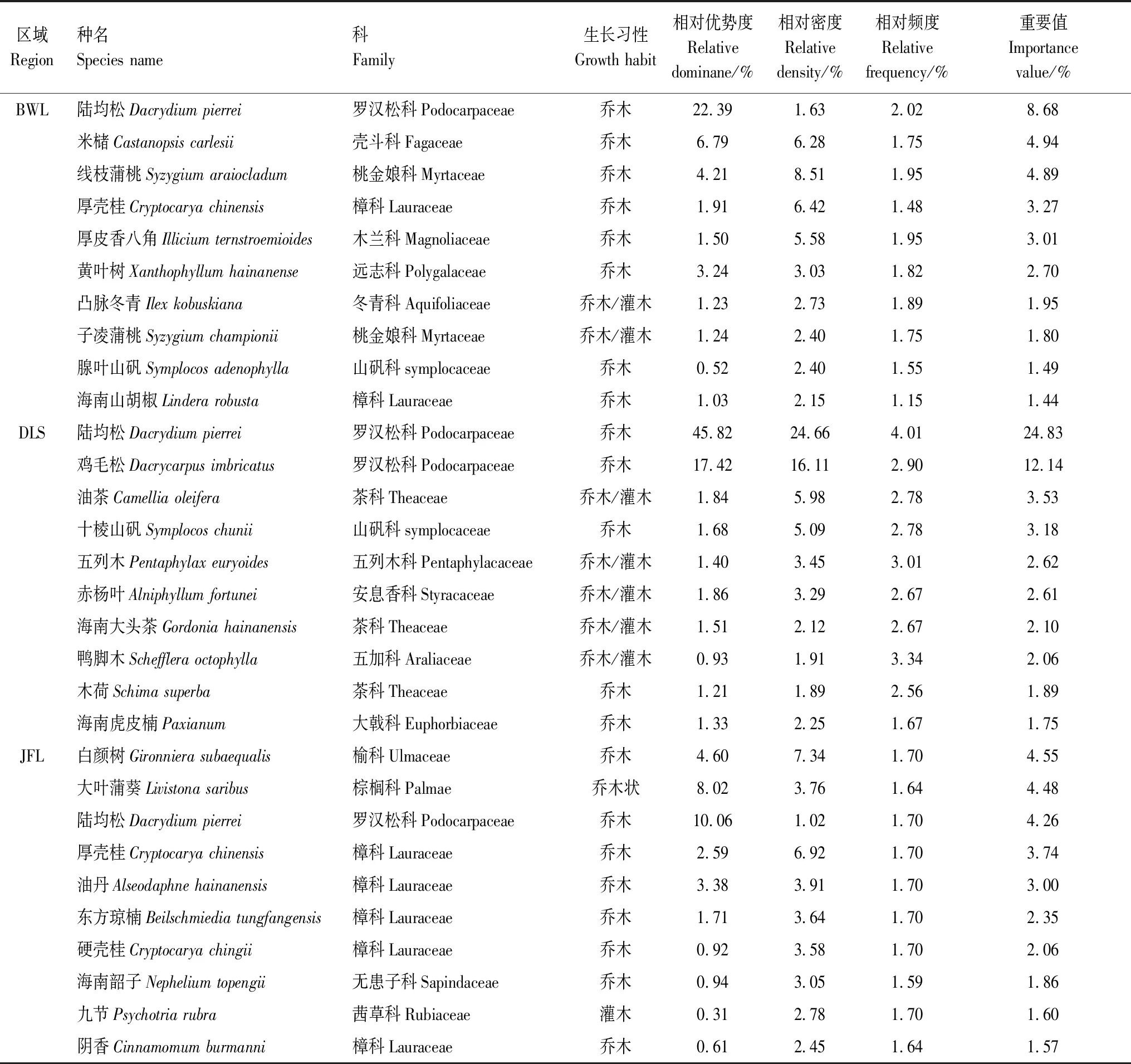
表3 陆均松天然群落乔木层重要值排前10位的优势种
2.2 陆均松种群龄级结构特征
3种陆均松种群的龄级结构特征如图2所示,BWL种群数量在幼年、中年和老年阶段均出现峰值,分别占种群个体总数的31.87%、34.07%、34.07%,3种年龄结构比例适中;但幼年个体中Ⅲ—Ⅴ龄级数量出现一定比例的缺失,从其整个生活史阶段分析,种群呈衰退趋势。DLS种群个体集中分布在幼年阶段,且在Ⅱ—Ⅴ龄级数量最多,占总数的98.45%;中年和老年阶段数量较少,2个阶段只占总数的1.45%,种群属于增长型。JFL种群幼年阶段数量较少,且在Ⅰ龄级出现缺失,只占总数的18.37%;中年阶段比例最高,占53.6%,数量多集中在Ⅶ—Ⅻ龄级;老年阶段占比28.57%,高于幼年阶段10.2%,由于幼龄个体的缺失,JFL种群属于衰退型。

图2 陆均松种群龄级结构特征Fig.2 D. pectinatum population age structure characteristics Ⅰ—Ⅻ 表示陆均松的年龄结构划分为第1—12龄级, I—XII indicates that the age of D. pectinatum is classified as 1st to 12th grade
2.3 陆均松种群空间分布格局
根据表4可知,3种陆均松种群的分布格局因所处的环境不同表现出差异,BWL种群整体呈聚集分布,V/M值为1.38,但不显著;DLS种群表现为聚集分布,V/M值为19.07;JFL种群呈均匀分布V/M值为0.44。

表4 陆均松种群空间分布格局
*P<0.05;***P<0.01;V/M: 方差/均值 Variance /mean;t:t检验ttest;K: 负二项参数K值 Negative binomial distribution indexK;m*/m: 聚块指数 Index of patchiness;M*: 平均拥挤度 Mean crowding;I: 丛生指数 Clumping index;C: 集群分布 Clump distribution;U: 均匀分布 Uniform distribution
根据表5可知,BWL幼年树呈聚集分布,成年树和老年树,呈均匀分布;DLS幼年树呈明显的聚集分布状态(P<0.01),成年树呈均匀分布,老年树呈聚集分布,但不显著;JFL幼年、中年、老年树均呈均匀分布。从聚集强度结果分析,3种陆均松种群整体的K、M*、m*/m、CA、I共5种指标表现为DLS>BWL>JFL,不同发育阶段的种群个体表现为幼年树>成年树>老年树。
3 讨论和结论
3.1 陆均松天然群落乔木层物种多样性及优势种组成
本研究中,BWL、JFL、DLS 三种陆均松天然群落内种群密度、物种多样性等指数极高,三者之间差异较小,但远高于中高纬度温带及亚热带森林群落;从乔木层优势种多度、优势度、重要值等层面分析,林内优势种形成了各自的种群区系特点,大多数特有种为小密度种群或极小密度种群,未出现大密度种群聚集分布现象,少有占绝对优势的树种,符合海南岛热带山地雨林物种组成的一般规律。低密度小种群容易产生近亲繁殖及遗传变异的消失,最终结果是易受各种生境胁迫的影响而在群落中消失[36- 37],同时也会导致各种群种内种间关系竞争加剧,这对陆均松天然群落未来的演替发展提出了挑战。陆均松天然群落林分结构以及物种多样性特点对于林内优势种的龄级结构及分布格局具有重要影响,贯穿整个生活史的始终,进而直接影响种群的更新演替。种群天然更新及生长繁殖机制复杂,除与自身的生理生态状况有关之外,外部环境因子和人为干扰也是其重要因素[38]。此外,林窗参量、空间分布特征等也起到重要作用。目前有关陆均松天然群落内优势种种间关系及其与环境因子、人为干扰的共同作用的影响机制还需更加深入的研究,这对于探索其天然群落物种多样性、优势种龄级结构及分布格局具有重要意义。

表5 不同发育阶段的陆均松种群空间分布格局
*P<0.05; ***P<0.01;V/M: 方差/均值 Variance /mean;t:t检验ttest;K: 负二项参数K值 Negative binomial distribution index K;m*/m: 聚块指数 Index of patchiness;M*: 平均拥挤度 Mean crowding;I: 丛生指数 Clumping index;C: 集群分布 Clump distribution;U: 均匀分布 Uniform distribution
3.2 陆均松种群龄级结构特征
种群龄级结构反应种群的动态变化及其发展趋势,缺乏可更新的幼苗幼树即可认定该种群趋向衰退[39]。目前有关陆均松种群结构的定论大多表明其天然更新困难,更新方式以种子繁殖为主,幼苗幼树数量极少[22,40- 42]。本研究中,3种陆均松种群龄级结构特征因所处环境不同表现出差异性;BWL种群幼年树数量占比小于中年和老年树,呈衰退趋势;DLS种群幼年树数量较多且远多于成年树和老年树,属于增长型;JFL种群中年树和老年树占有较高比例,幼年树数量较少,属于衰退型。分析认为,本研究与以往结论存在一定差异的原因可能有两方面,一是研究区域、取样方法、龄级划分标准的不同[30],二是陆均松种群对生长环境的要求。本研究调查的区域包括BWL、DLS、JFL三种不同环境的热带山地雨林,海拔均在800 m 以上,BWL和JFL极少遭到人为干扰和破坏,陆均松群落保存最为完整;DLS实验区的陆均松群落 20 世纪 60 年代经历过不同程度的采伐,间接为其创造了适度的小生境促进其生长繁殖,这可能也是造成其中年、老年树数量较少,幼年树数量较多且属于增长型种群的主要原因;在取样过程中,样地设置以中老年陆均松为主,可能会造成幼苗幼树数量的缺失;本研究中将DBH≤40 cm 的个体划分为幼年树,这一划分标准将更多的陆均松个体划分为幼年树。另一方面,陆均松生长条件苛刻,对环境因子的依赖性强,而热带山地雨林林分郁闭度高,光照不足,土壤潮湿,林下枯枝落叶层厚度可达 2 cm以上[43- 44],不利于种子保存和繁殖,且生长和竞争能力极弱的幼苗幼树,也极易受到环境筛选被淘汰。调查发现BWL和JFL的陆均松天然群落平均林分郁闭度大于0.80,平均枯枝落叶层厚度大于2.55 cm,林内极少有陆均松幼苗,因此推测,外部环境因素是造成BWL、JFL种群幼苗和幼树缺少而逐渐演变成为衰退型种群的另一个重要原因。Milios等[45]对希腊东北部罗多彼山脉冷杉幼苗研究发现林内过分阴蔽是导致幼苗幼树高死亡率的重要因素,Narukawa等[46]和Lusk等[47]也认为林内过度荫蔽、土壤潮湿不利于植被种子繁殖进而导致幼苗幼树缺乏进而演变为衰退型种群。
3.3 陆均松种群空间分布格局
种群空间分布格局的形成与种群自身的特性(种子扩散机制、植物繁殖特性、种内种间竞争等)、外部生境条件过滤(地形、土壤、水分、光照等)有关,是不同尺度上各种生物因子和非生物因子长期综合作用的结果[48- 49]。本研究发现,BWL和DLS陆均松种群整体及在幼年树阶段呈聚集分布状态,成年树及老年树阶段基本呈均匀分布;而JFL 整体及各发育阶段均呈均匀分布(可能与取样位置有关,样地内极少见到陆均松幼苗);3种陆均松种群整体聚集强度表现为DLS>BWL>JFL,不同发育阶段的种群个体表现为幼年树>成年树>老年树。这可能与种群自身的生长繁殖机制、种间竞争、生境异质性、人为干扰等方面有关。其一,幼年陆均松在生活史初期受种子扩散机制影响,散播于母树周围,数量集中,受母树庇护,汲取林内多余的水分、光照及土壤养分,在生境适宜地段发育成苗,形成生活史初期的聚集性分布格局[1,18]。种群进行到生活史的中后期,植株个体为获得足够的生长资源对光照、养分、水分需求增大,种内、种间竞争加剧导致个体数量锐减,发生自疏与它疏[50],由聚集分布转变为随机分布或者均匀分布,聚集强度也逐渐下降。这与Jiang等[51]和 Wilcox和Elderd[52]认为的随着种群年龄的增加,种群格局由聚集分布向随机分布或者均匀分布转变或聚集强度逐渐降低的研究结果一致[51- 52]。其二,陆均松天然群落结构复杂,群落内物种多样性极高,必然导致种间竞争激烈,此外,相邻植株往往不是同种个体,异种个体的影响也是影响其分布格局的重要因素。其三,3种陆均松种群所处的研究区域,其年均温、年均降水量、海拔和坡度等地形因子、林分因子等环境因素(表1)存在一定的差异;因此可推测,陆均松种群格局特征与生境异质性密切相关,这与郭垚鑫等[53]和Hao等[54]的研究结果一致,在较小尺度上,物种的分布格局主要由种内种间竞争、种子的扩散机制决定;而较大尺度上的空间分布格局则更多受到生境异质性的影响;黄运峰等也表明,扩散作用是形成热带低地雨林树木空间分布格局最重要的生态过程,其次是生境异质性和扩散作用的联合作用[55];本研究未对环境因子与陆均松种群格局进行更加深入的研究,这一推测有待进一步验证。其四,研究发现,未受到人为干扰的BWL和JFL种群幼年树数量较少,且多为均匀分布,而受到不同程度采伐的DLS种群幼年、成年树数量很多,且呈高密度聚集分布,因此认为,适度的人为干扰也是影响陆均松种群格局的重要原因,但合理的采伐强度有待进一步研究和验证。
本文从陆均松天然群落乔木层物种多样性及优势种组成、陆均松种群年龄结构、种群分布格局3个方面分析了 BWL、DLS 和 JFL 陆均松种群的生存现状及种群格局的空间异质性。研究发现,陆均松天然群落结构复杂,物种多样性极高,林内多为小密度或极小密度种群,少有占绝对优势地位的树种;BWL和JFL陆均松种群幼苗幼树数量较少甚至缺失,呈衰退趋势,经历过不同强度采伐的DLS种群则属于生长型。3种陆均松种群空间分布格局与其自身的生长繁殖机制、种间竞争、生境异质性、人为干扰等因素密切相关。陆均松是海南热带山地雨林的特有种和关键种,具有重要的科研价值,鉴于其目前依靠自身进行天然更新存在困难的处境,可根据其实际情况,利用人工辅助措施如开辟林窗、扰动土壤以促进种子落入土壤、伐除母树和幼苗周围的乔灌木、增加土壤养分、合理疏伐等方式[35,56]为其天然更新和生长繁殖创造有利的生存环境;此外,加强对陆均松遗传基因多样性的研究也是保护其更新繁衍的重要方式。
