中国缟蝇科(双翅目:缟蝇总科)高级阶元的几何形态学分析
2020-06-11戚凌何向怡李文亮
戚凌, 何向怡, 李文亮
(河南科技大学林学院,河南洛阳471023)
双翅目Diptera的分类较为复杂,迄今未形成统一的分类体系。目前一般按照Delfinado和Hardy(1977)、Richards和Daries(1977)的分类标准,将缟蝇科Lauxaniidae划属于双翅目环裂亚目Cyclorrhapha无瓣类Acalyptrata有缝组Schizophora缟蝇总科Lauxanioidea(高彩霞,2003);目前世界已知 3亚科172属约2 150种,我国已知32属343种(李文亮,2014)。缟蝇总科是无瓣蝇类各总科系统进化过程中较为原始的类群之一,对缟蝇科昆虫的研究是揭示无瓣蝇类各总科间系统发育关系的重要环节(李文亮,2014)。在缟蝇科分类中,往往使用传统的形态学方法鉴定,即观察记述标本的外部分类特征,然后进一步解剖、鉴定、绘图等,需要花费大量的时间和精力。
近年来几何形态分析方法越来越成熟,并在分类学、系统发育学、生物进化学等方向有所应用。20世纪早期,生物学从定性描述向定量描述过渡;20世纪中期,传统的形态测量学产生;20世纪80年代,形态结构数值化以及数据分析技术得到重要突破,生物体形态的几何信息能够被获得,并用于综合分析(Rohlfetal.,1998;Perrardaetal.,2016)。这种方法利用了变量之间的角度转换、物体表面的纹理甚至实物标本的物理性能等数据信息,能够将生物体形态信息中的大小和形状分开(白明等,2014)。目前在几何形态学分析中,最常见的方法是标点法,该方法能够找到研究样本之间的形态“同源信号”和“简约信息位点”,排除样本之间由于位置排列产生的差异(Yanetal.,2011)。
翅是昆虫获得飞翔能力的重要形态结构,翅的形状与昆虫的生存方式、扩散能力有着密切的关系,翅形的变化有可能对物种的分化形成起到关键的作用(白明,杨星科,2007)。李玲等(2017)曾基于几何形态学对3种实蝇翅脉形态进行了分析,并证明几何形态学方法可用于实蝇种间的分类。段琛等(2018)也通过几何形态学方法对3种库蠓的翅形变化进行了分析,并成功找出了翅的主要变异区域。
目前尚无用几何形态学方法对缟蝇科翅脉进行研究的报道,而本试验运用几何形态学分析中的标点法分析了翅脉在缟蝇科分类标准上的科学依据,并利用几何形态学的原理对缟蝇科高级阶元进行初步分析,探讨翅位点的变异度,为缟蝇科系统分类开拓新的途径,并为缟蝇科几何形态学系统发育研究打下初步基础。
1 材料与方法
1.1 标本材料
缟蝇科昆虫标本来源于河南科技大学昆虫标本馆和中国农业大学昆虫博物馆,共2亚科20属252种(表1)。
1.2 试验方法
1.2.1 获取图像用EOS450D/5D相机拍摄照片,再用Adobe Photoshop CS将图片设置为灰度模式,并调整翅脉的角度。
1.2.2 数据提取及标准化处理用tps Util将翅脉图片格式文件转化为tps格式文件,然后分别对样本进行同源标点,每个样本均按照同样的位置、顺序标注14个标点(图1):(1)前缘脉(C)与肩脉(h)交点,(2)C与亚前缘脉(Sc)交点,(3)Sc与第一径脉(R1)交点,(4)第二、三合径脉(R2+3)与翅缘交点,(5)第四、五合径脉(R4+5)与翅缘交点,(6)第一中脉(M1)与翅缘交点,(7)第一肘臀合脉(CuA1)与翅缘交点,(8)第二臀脉(A2)末端,(9)第二肘臀合脉(A1+CuA2)末端,(10)径中横脉(r-m)与R4+5交点,(11)r-m与M1交点,(12)中肘横脉(dm-cu)与M1交点,(13)dm-cu与CuA1交点,(14)翅外缘黑色短鬃的末端。

表1 缟蝇科标本种类统计Table 1 Statistics of specimens in Lauxaniidae family
1.2.3 数据分析利用MorphoJ基于主成分分析(principal component analysis,PCA)和典型变量分析(canonical variate analysis,CVA)对翅脉的几何形态学数据进行分析。先进行原始数据的主成分分析,再从不同方向分组进行不同阶级的主成分分析和典型变量分析,并通过PCA和CVA的薄板样条图,详细解析每个标点所代表的翅脉的差异性。
2 结果
2.1 缟蝇科总体分析
全部样本的PCA结果(图2)显示:在提取的24个主成分中,第一主成分(PC1)占总变异量的30.010%,第二主成分(PC2)占总变异量的18.698%,第三主成分(PC3)占总变异量的15.377%,前三个主成分的特征值占总变异量的64.085%,能够说明各样本之间的主要差异。
薄板样条分析结果(图3)显示:在PC1中,扭曲度最大的是标点4和14;在PC2中,标点10和11的扭曲度相对较大。说明在缟蝇科的进化与变异中,R2+3、黑色短鬃和r-m是变异度较高的翅脉,在进化中相对较不稳定。

图1 缟蝇科昆虫翅的标点Fig. 1 14 landmarks on the wing of Lauxaniidae
C. 前缘脉, h. 肩脉, Sc. 亚前缘脉, R1. 第一径脉, R2+3. 第二、三合径脉, R4+5. 第四、五合径脉, M1. 第一中脉, CuA1. 肘脉(Cu)与第一臀脉(A1)的部分段合并而成的第一肘臀合脉, A2. 第二臀脉, r-m. 径中横脉, dm-cu. 中肘横脉, A1+CuA2. Cu与第一、二臀脉(A1、A2)合并而成的第二肘臀合脉
C. costa, h. humeral vein, Sc. subcosta, R1. first radial vein, R2+3. combined vein of second and third radial vein, R4+5. combined vein of fourth and fifth radial vein, M1. first medial vein, CuA1. combined vein of cubital and first anal vein, A2. second anal, r-m. radial-median crossvein, dm-cu. median-cubital crossvein, A1+CuA2. combined vein of cubital, first and second anal vein

图2 缟蝇科昆虫翅脉主成分分析Fig. 2 Principal component analysis for the wing veins of Lauxaniidae
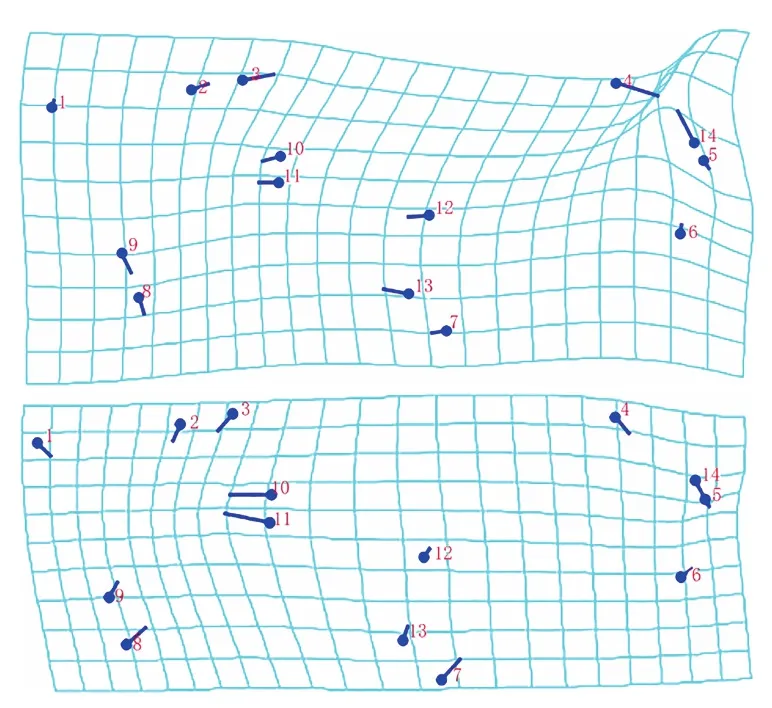
图3 缟蝇科昆虫翅脉PC1与PC2的薄板样条Fig. 3 Thin plate spline of PC1 and PC2 for the wing veins of Lauxaniidae
1~14见图1
1-14 see Fig.1
2.2 同脉缟蝇亚科与缟蝇亚科的分析
在总体分析的基础上,已知前3个主成分的特征值占总变异量的64.085%。缟蝇科2亚科的PCA结果(图4)显示:同脉缟蝇亚科主要聚集在PC1轴的0点以左(-0.10~0),而缟蝇亚科主要聚集在PC1轴的0点以右(0~0.01);两者在PC2方向上并没有明显差异。缟蝇科2亚科的CVA结果(图5)显示:同脉缟蝇亚科与缟蝇亚科仅在横轴的0点附近有小部分重合,而绝大部分区间是非重合的。同脉缟蝇亚科与缟蝇亚科在PC1和CV1方向上具有明显的差异,说明这2个亚科能够通过几何形态学方法分析翅脉特征来进行分类鉴定。

图4 同脉缟蝇亚科(红色)与缟蝇亚科(蓝色)翅脉的主成分分析散点
Fig. 4 Principal component analysis for the wing veins of Homeoneuriae (red) and Lauxaniinae (blue)
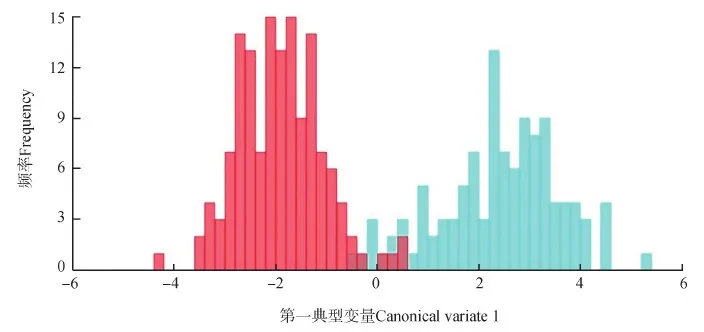
图5 同脉缟蝇亚科(红色)与缟蝇亚科(蓝色)翅脉的典型变量分析柱形Fig. 5 Canonical variate analysis for the wing veins of Homeoneuriae (red) and Lauxaniinae (blue)
CV1与PC1的薄板样条图(图6)显示:扭曲度最大的点均为标点4、14,扭曲度较小的均为标点1、5和6。说明R2+3和黑色短鬃的变异度最大,而C、R4+5和M1的变异度较大。
2.3 缟蝇科4属间的分析
为了找出缟蝇科在属级特征上的翅脉变化,本试验选择了4个样本数量相对较多的属(表2),即同脉缟蝇属Homoneura、黑缟蝇属Minettia、四带缟蝇属Tetroxyrhina和斑缟蝇属Trypetisoma进行CVA。在CVA中,前3个典型变量分别占总变异量的74.767%、18.956%和6.277%,共计100%。
CVA结果(图7)显示:不同属各自聚集在不同区域,说明这4属在翅脉上能作出明显区分。
薄板样条图(图8)显示:在CV1薄板样条中,标点4和14相对扭曲度较大,且它们的变异趋势相反,说明R2+3与翅缘的交点向上变异,黑色短鬃则越来越靠近标点5,R2+3与短鬃间的距离是逐渐增大的。在CV2薄板样条中,扭曲度较大的是标点10和11,说明r-m有相对右移且增长的变异趋势;标点1、2、6和7的变异度极小,说明这些点所对应的C、Sc、M1和CuA1是相对稳定、不易变异的。
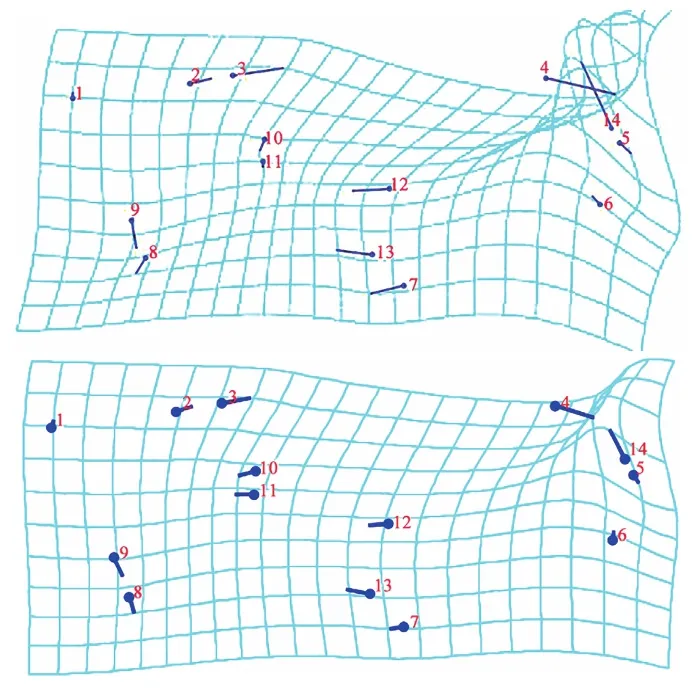
图6 同脉缟蝇亚科与缟蝇亚科翅脉CV1和PC1的薄板样条
Fig. 6 Thin plate spline of CV1 and PC1 for the wing veins of Homeoneuriae and Lauxaniinae

表2 缟蝇科4属间分析的样本数量Table 2 Number of samples for the 4 genera of Lauxaniidae
3 结论与讨论
3.1 结论
总体分析、两大亚科分析和4个常见属分析结果显示,R2+3与翅缘交点、黑色短鬃端点和r-m端点是变异度较大的位点,R4+5与翅缘交点、M1与翅缘交点、CuA1与翅缘交点为稳定的位点;翅缘黑色短鬃端点可作为亚科的重要鉴别特征,这与传统的缟蝇科分类标准相吻合,证明了本次试验的可行性与科学性;4属分析中属间离散度较高,说明标点法能较好地进行属级阶元的鉴别。
3.2 讨论
本试验得到的几何形态学分析结果与传统形态学鉴定结果相一致,即二者的相互印证度较好,证明了几何形态学方法在缟蝇分类中应用的可行性。但本试验采用的部分缟蝇属的样本数量较少,且每种仅选取了1个样本,可能会对试验结果有一定影响。在属间分析中,4个常见属不足以代表整个缟蝇科,下一步可以增加各个种的样本数量,扩充样本后进一步在低级阶元如属间或种间进行分析研究。此外,本试验选取的标点数量较少,且未结合翅轮廓的变化进行比较,研究结果只能确定一些差异性较大的翅脉。而翅的轮廓特征也是相对较重要的翅特征之一,曾有用半标点法结合翅轮廓对蚁蛉科Myrmeleontidae昆虫进行几何形态学分析的研究(詹庆斌,2014),所以可以更进一步结合缟蝇翅轮廓的研究来补充证明翅在分类依据上的重要性。

图7 缟蝇科4属翅脉的典型变量分析散点
Fig. 7 Canonical variate analysis for the wing veins of the 4 genera of Lauxaniidae
H. 同脉缟蝇属Homoneura, M. 黑缟蝇属Minettia, TET. 四带缟蝇属Tetroxyrhina, TRY. 斑缟蝇属Trypetisoma
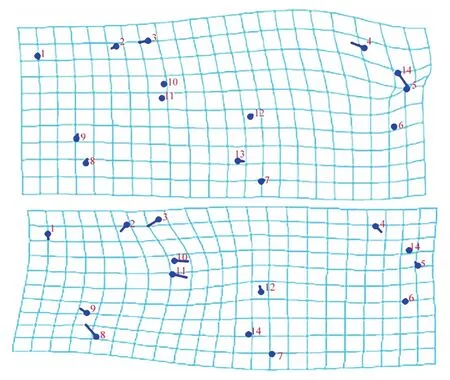
图8 缟蝇科4属翅脉的CV1与CV2薄板样条Fig. 8 Thin plate spline of CV1 and CV2 for the wing veins of 4 genera of Lauxaniidae
1~14见图1
1-14 see Fig.1
除了将翅作为分析对象,几何形态学方法还能够通过昆虫其他部位的特征来进行分类鉴定等研究。在关于昆虫分类地位的研究中,李荣荣等(2019)通过几何形态学方法对二星蝽属Eysarcoris及其近缘属昆虫的小盾片形状进行了分析,并根据试验结果将二星蝽属划归于二星蝽族。在有关昆虫进化发育的研究中,曾有利用几何形态学方法对锹甲的前胸背板与鞘翅进行分析从而探究其演变进化的试验,该研究使用半标点法标注了锹甲前胸背板与鞘翅的外轮廓,并结合分子数据建立了系统发育树,对锹甲的外部形态演变作出了推断(Zhangetal.,2019)。在昆虫形态与功能关系的研究中,佟一杰等(2016)对蜣螂后胸叉骨的形态多样性进行了几何形态学定量评估,证明滚粪球或直接掘洞的行为会对蜣螂后胸叉骨的形态有显著性影响。基于上述成功的研究案例,本试验下一步考虑将除了翅以外的其他部位特征作为几何形态学分析对象。在缟蝇科昆虫中,背中鬃着生位点是一个显著的形态分类特征,所以本试验将尝试用几何形态学方法分析中鬃着生位点,以期能够与翅的分析结果相结合,对缟蝇科昆虫进行更准确的分类。
近年来,昆虫学依据其独有的特性及后发优势,将几何形态学的应用推向一个新的高度,未来将不断开拓新的研究模式与方向。昆虫占领了众多生态位,并产生了极为多样的形态适应,通过几何学与发育生物学、支序系统学、古生物学、生态学等方面证据的深度结合与相互印证,可从独特的角度对相关科学问题进行解读(Baylacetal.,2003)。
