世界大麦种质资源遗传多样性分析
2020-06-10张蓝天楚单单方云霞吴玉环陈建明薛大伟张晓勤
张蓝天,楚单单,方云霞,吴玉环,陈建明,薛大伟,张晓勤
(1. 杭州师范大学生命与环境科学学院,浙江 杭州 311121; 2. 浙江省严州中学新安江校区,浙江 杭州 311607)
作物种质资源是经过长期的自然进化和人工选择后形成的,积累了极其丰富的遗传变异,是作物育种的物质基础.作物品种的遗传改良进程,很大一部分取决于不同类型优质品种资源的利用.对作物种质资源进行正确合理的鉴定和评价是进行有效改良的基础.大麦(HordeumvulgareL.)是世界上古老的栽培作物之一,对环境适应性很强[1],广泛分布于世界范围内,栽培面积在谷类作物中仅次于小麦、水稻及玉米,位居全球第四[2],可作为粮食、饲料,也可用于啤酒酿造、保健品研发等[3].
不同国家和地区种植的大麦存在很大的差异.目前,大麦品种资源由于长期的驯化栽培、自然进化和遗传定向改良,导致其遗传基础比较单一,遗传背景日趋一致,在品质和抗逆性等方面一直没有大的突破.因而,研究不同地区大麦种质资源的遗传多样性,是保护及开发利用优质大麦新种质的基础[4].关于大麦遗传多样性的研究已有较多的报道[5-11],但大多局限于某一国家或地区,对世界大麦种质资源遗传多样性分析的报道较少.
分子标记由于具有不受季节与环境的影响、无表型效应、重现性好、简单快捷等优点,已经越来越多地被用于植物遗传多样性研究,常用的分子标记技术有RFLP、AFLP、RAPD、ISSR、SSR、SRAP等.SRAP (sequence-related amplified polymorphism)——相关序列扩增多态性标记技术,是2001年由Li等[12]在PCR基础上发明的一项DNA分子标记技术.由于该标记简便、快速、不需预知物种的序列信息,目前已应用于植物遗传多样性分析、种质鉴定、遗传连锁图谱的构建、基因连锁标记、基因定位、比较基因组学研究及杂种优势预测等方面的相关研究[13].其中尤以遗传多样性的检测应用最为突出,在西瓜、辣椒、油菜和马铃薯等多种植物上成功地进行了遗传多样性分析[9,14-15].
本实验以来源于不同国家和地区的115份大麦微核心种质资源为材料,采用SRAP分子标记技术分析其遗传多样性,以期为大麦的遗传基础研究和育种应用提供参考.
1 材料与方法
1.1 大麦种质
大麦微核心种质资源由浙江省农业科学院提供,115份大麦种质共来源于4个大洲的33个国家和地区,其中亚洲44份,北美洲5份,欧洲49份,非洲17份.
1.2 实验方法
1.2.1 基因组DNA的提取
大麦种子萌发在26 ℃暗室中进行,空气相对湿度约为70%.待大麦幼苗长至两叶期,取3 cm左右幼嫩叶片用于基因组DNA提取,参照Xue等[16]提供的方法并进行少许改动.
1.2.2 SRAP分析
根据多态性高的原则,从Li等[12]公布的引物序列中选取17对SRAP引物(表1),并交由上海英维捷基生物技术有限公司合成.SRAP反应体系参照Xue等[16]提供的方法并进行少许改动.

表1 17对SRAP引物序列Tab.1 17 primer sequences of SRAP-PCR
1.3 数据分析
对凝胶成像系统收集到的图片进行分析,并通过人工分析统计得出各个引物对115份大麦种质资源品种扩增的SRAP标记条带.谱带按照0/1矩阵记录,在相同迁移位置上有SRAP谱带记为“1”,无谱带记为“0”,建立矩阵后进行数据统计.利用GenAlEX 6.5软件[17]、Arlequin 3.5软件[18]和STRUCTURE (http://pritch.bsd.uchicago.edu/structure.html)对所获得的数据进行相关分析.
2 结果与分析
2.1 SRAP引物的筛选及扩增
为了获得稳定可靠、重复性好的SRAP引物,本实验前期共计筛选标记72个,从中挑选出17对扩增条带清晰、重复性好的SRAP引物组合用于进一步的实验(图1).利用这17对引物对不同地域来源的大麦种质进行分析.结果表明,所有引物在不同品种中均能扩增出3条以上的不同条带,共计获得114个扩增条带,其中组合em4-me8、em7-me8和em1-me9获得扩增条带最多,为10条.可见,本研究选用的引物检测效率较高,在各材料间具有较好的多态性.

图1 引物em6-me8和em4-me8对部分大麦材料DNA的SRAP扩增结果Fig.1 Profiles produced in some barley varieties by the SRAP primer em6-me8 and em4-me8
2.2 SRAP引物多态性分析
利用GenAlEX 6.5软件对不同来源的115份大麦微核心种质的SRAP扩增结果进行了分析,其有效等位基因数(Ne)、等位基因杂合性(He)、期望杂合度(uHe)和Shannon’s信息指数(I)分析结果见表2.结果表明,em4-me2和em8-me6引物组合中所扩增出的3个条带在所有种质中均未表现出差异,占全部位点的4.39%.在114条扩增条带中,出现频率小于5%的稀有位点共6个,占全部扩增位点的5.26%.所有用于分析的引物在有效位点上的差异不大,Shannon指数分析表明不同引物存在非常显著的多态性差异,em7-me2引物Shannon指数最高,为0.581,em8-me6引物则最低,为0.23.这说明em7-me2引物的相关位点在不同品种中存在很大差异,可以为后期多态性标记的发展奠定基础.

表2 SRAP引物组合的遗传多样性Tab.2 Genetic diversity of SRAP primers
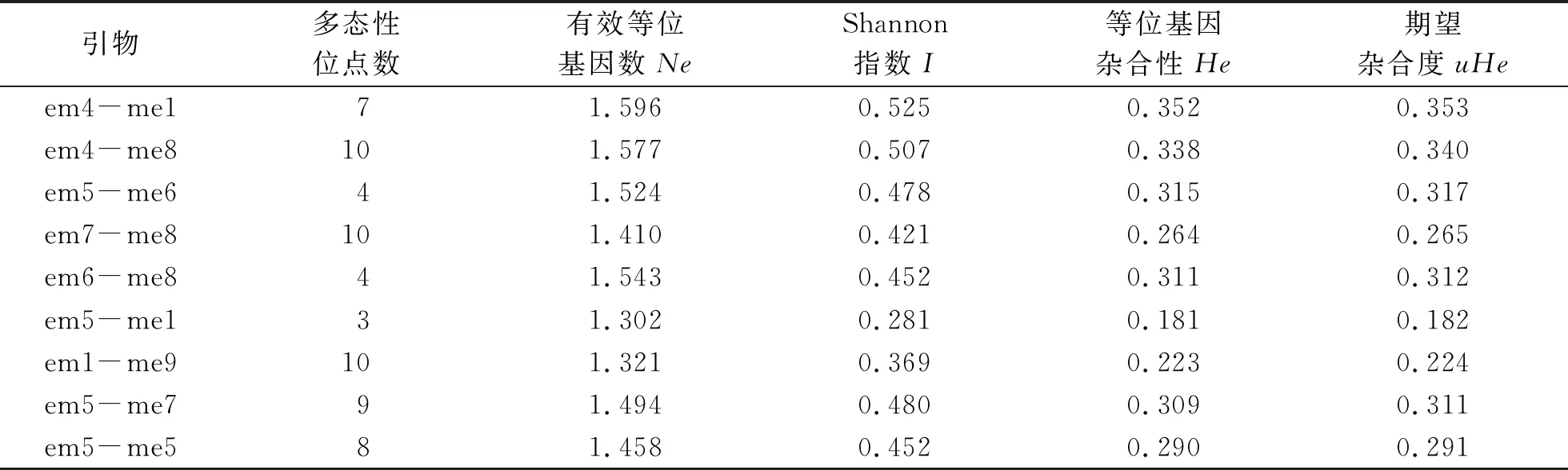
续表2
为进一步了解SRAP标记在这些不同来源大麦种质间的差异,利用种质的来源地点将这些大麦分为不同的群体,进一步展开多态性分析.结果发现,这些引物在亚洲来源的大麦中表现出最高的多态性,欧洲次之(表3).这表明在欧洲和亚洲,大麦品种呈现更为丰富的多样性.多态性片段的比例分析表明,亚洲(87.72%)和欧洲(84.21%)的大麦种质拥有最高的多态位点,非洲次之(78.95%),而美洲则最低(42.98%).

表3 SRAP引物组合在不同地理来源大麦种质中的多态性分析Tab.3 Polymorphism analysis of SRAP primer combinations in different geographical source of barley germplasm
2.3 遗传结构分析
利用Arlequin 3.5软件对不同来源的大麦群体进行AMOVA分析,以进一步了解不同大麦种质间遗传差异的主要来源.结果显示种群内的基因变化大大丰富了遗传多样性(表4).种群内的变异数占总变异的100%,说明这些种质间存在的差异全部来源于群体内.

表4 大麦种质群体差异的AMOVA分析Tab.4 AMOVA analysis of the population difference of barley germplasm
以上分析表明大麦种质呈现复杂的遗传多样性,而且遗传差异主要来源于群体内,因此为了更清楚地了解群体遗传结构,利用STRUCTURE软件进行了混合模型分析.结果表明,在K=4时,LnP(D)值维持一个高值,表明115份供试的品种可划分为4个类群,分别对应来源于4个洲的大麦品种(图2).

图2 115份大麦种质的群体遗传结构Fig.2 Genetic structure of 115 accessions of barley germplasm
2.4 33个国家和地区大麦种质的聚类分析
将115份大麦种质按国家和地区来源不同分为33个群体,在利用GenAlEX 6.5软件进行遗传距离运算的基础上,用MEGA软件聚类分析(图3),在截距0.75处可将33个群体划分为7个组,分别命名为组A—G.组A包括非洲和美洲的7个国家,组B、C和F只包括中国、日本和韩国各1个亚洲国家,组D包括5个亚洲和非洲国家,组E包括2个亚洲国家,组G则包括所有的15个欧洲国家和亚洲的尼泊尔.

图3 基于SRAP标记数据的不同国家和地区大麦种质资源遗传距离UPGMA聚类图Fig.3 UPGMA dendrogram using SRAP allele frequencies data, showing the relationship among the 33 countries and regions of barley
3 讨论
种质资源研究是展开育种改良工作的基础,遗传基础狭窄会导致难以培育出突破性品种.因此分析种质资源的遗传多样性,比较种质资源间亲缘关系的远近,对于育种工作具有重要的意义.SRAP标记具有简单性、可靠性、灵活性、可以检测多个基因位点以及成本低廉等优势,目前被广泛应用于植物遗传多样性分析.如郭蕾蕾[19]利用SRAP技术共标记了216对引物,并在亲本间进行多态性筛选,结果筛出38对SRAP标记,共产生77个SRAP多态性位点.刘仙俊[20]选用54对SRAP引物对亲本进行多态性筛选,其中21对SRAP引物在群体中扩增出52个清晰且有差异的位点.
本研究利用7个正向引物和9个反向引物组成的17对SRAP引物组合对115份世界大麦种质进行多态性分析,共扩增出114个多态条带,平均每对引物检测到6.3个等位变异,变异范围为3~10;Shannon’s多样性指数平均为0.430 6,属于具有较高多态性的位点.多样性指数和等位变异数低于前人利用SSR标记研究的结果[21],其可能原因在于SRAP标记与SSR标记扩增产物不同,SRAP扩增基因区域不能揭示非基因区域的遗传多样性,而SSR可扩增大麦基因区和非基因区,所揭示的是大麦基因组整体的遗传多样性水平.以上结果表明,33个国家的115份大麦材料具有较高的遗传多样性,能够很好地反映这些大麦品种间的遗传特性,是进行大麦遗传改良的丰富基因库.
从大麦种质不同洲际间的遗传多样性分析可以看出,亚洲种质资源的遗传多样性Shannon指数最高,其次是欧洲,而北美洲种质资源遗传多样性最低,这可能与样本的遗传基础和所处地理位置有关.国际大麦遗传学界普遍认同大麦的起源中心为中东-肥月亮湾地区,最近的研究发现中国西藏野生大麦和中东野生大麦是两个独立起源进化的群体,而中国栽培大麦的起源分支源于西藏野生大麦,从而证实了栽培大麦西藏起源中心假说[22].本研究也发现,欧洲和亚洲大麦品种呈现更为丰富的多样性(表3),这与前人的结果相类似,但由于各个国家的大麦样本量不同,而且之间差异太大,验证该结论尚需进一步的研究.
聚类分析是多种种质亲缘关系研究中应用较多的方法之一,能量化品种之间的差异.本研究通过聚类分析量化了中国和韩国及日本大麦种质资源之间的关系.此外,根据17对SRAP引物扩增出的多态位点,将33个国家的大麦群体划分为7个组,各组在一定程度上反映出品种的地理来源特点.这一结果与STRUTURE的分析结果相一致,暗示地理因素是形成物种分化的最直接原因,也是限制物种基因交流的最有效条件.大麦作为自交植物,在传播、利用和推广过程中,地理隔离就成为保持品种特性的重要保障.从图3的系统发育树可以看到,地理位置邻近地区的品种总是聚集在同一类群,例如组G中的欧洲大麦种质和组A中非洲大麦种质.来自北美的5份大麦种质和非洲的大麦种质聚在一组,说明美洲和非洲的大麦种质具有较近的亲缘关系.亚洲国家尼泊尔的大麦种质与欧洲15国的大麦种质聚为一类,这为研究欧洲大麦种质的起源和传播具有一定的借鉴意义.另外,虽然中国、韩国、日本3国之间的距离很近,但各自聚为一类.此外,Mantel测验表明供试种质遗传差异并不与物理分割呈现相关性,这暗示着引种等人类活动会对大麦种质多样性产生巨大影响.然而,大麦种质呈现的复杂性及各个地区的巨大差异性还有待深入的研究.
本研究利用SRAP分子标记技术对来自33个国家和地区的115份大麦种质资源进行遗传多样性研究,结果表明来自亚洲和欧洲的大麦具有更多的多态性位点,遗传多样性最为丰富且115份样本的遗传差异均来自群体内部.聚类分析结果显示来源地相同的大麦品种间亲缘关系更近,这与实际情况一致.所得结果揭示了不同来源的大麦品种间的亲缘关系和遗传多样性差异及其来源,有利于初步了解大麦的遗传背景,并为大麦育种的改良工作提供了理论依据.
