外源脱落酸对荒漠植物红砂幼苗光合及叶绿素荧光特性的影响
2020-06-09曾继娟种培芳朱强
曾继娟 种培芳 朱强

摘要:以2年生红砂幼苗为试材,采用盆栽试验,通过喷施浓度为0.1、1、10、100 μmol/L的脱落酸(ABA)溶液,研究外源ABA对红砂幼苗光合和叶绿素荧光参数的影响,旨在为荒漠区植被恢复提供依据。结果表明,叶面喷施ABA后,气孔导度(Gs)、蒸腾速率(Tr)和净光合速率(Pn)都在短时间内得到较好的抑制,但是当ABA喷施浓度为100 μmol/L 时,Pn下降,Gs的变化趋势不明显,主要与气孔因素有关,说明适度的ABA浓度(<100 μmol/L)有助于红砂更好地适应逆境,超过此浓度,则不利于红砂进行光合作用。对照的PSⅡ原初光能转化效率(Fv/Fm)、PSⅡ的潜在光化学活性(Fv/Fo)、最大荧光/初始荧光(Fm/Fo)、可变荧光(Fv)均维持在较高水平,其中Fv/Fm>0.8,外施ABA后,Fv/Fm、Fv/Fo、Fm/Fo、Fv均不同程度地降低,随着ABA浓度的变化,各处理的变化趋势一致;随着ABA胁迫时间的延长,Fv/Fm、Fv/Fo、Fm/Fo、Fv均低于胁迫初期。外施ABA对红砂光合生理机能的影响表现为多个指标的相互作用,并受ABA胁迫浓度、胁迫时间共同影响。综合分析表明,喷施一定浓度的脱落酸可以维持红砂生长季叶片正常的生理代谢功能,从而有效控制叶片的衰老进程。
关键词:红砂;脱落酸;光合特性;叶绿素荧光
中图分类号: Q945.78文献标志码: A
文章编号:1002-1302(2020)07-0136-06
近年来,全球气候变化和过度的人为活动加剧了我国北方荒漠草原的沙漠化进程,其生态环境极为脆弱,成為制约我国“西部大开发”战略实施的重要因素[1]。植被恢复与重建可以有效抵御荒漠化威胁,而水分是影响植物生长与分布的关键因子。因此,研究荒漠植物抗旱的生理生态适应机制,采用有效的方法提高植物的抗旱性,对于荒漠区植被恢复与重建意义重大。
红砂(Reamuria soongarica)是柽柳科红砂属超旱生小灌木,广泛分布于我国干旱荒漠地区,其抗逆性强,生态可塑性大,具有较强的抗旱、耐盐和集沙能力[2],是西北干旱、半干旱地区的荒漠复苏植物和主要的牧草饲料[3-4]。近年来,对于红砂的研究主要集中于种子萌发[5]、幼苗建植[6]、遗传多样性[7]、耐盐[8]、抗旱以及气候因子[9-10]等生理生态学方面,这对于分析荒漠化进程中水分对红砂的影响机制具有重要作用。
脱落酸(abscisic acid,ABA)最初被认为是一种抑制型激素,随着研究的不断深入,发现ABA在植物整个生长发育过程中不但能诱导植物休眠、合成种子贮藏蛋白,还在气孔开闭、水分调节、光合作用、衰老及植物对逆境的适应等多方面起到重要的调控作用[11]。干旱胁迫在诸多自然逆境中占据首位,成为制约许多地区农牧业发展的瓶颈。植物根系最先感知到外界水分胁迫刺激,诱导ABA的生物合成,通过一系列代谢过程降低气孔开度来调节植株水分的散失和生长等。目前,干旱胁迫下脱落酸对小麦(Triticum aestivum L.)、玉米(Zea mays)、樟子松(Pinus sylvestris)、尖果沙枣(Elaeagnus oxycarpa Schlecht.)、生姜(Zingiber officinale Roscoe)、拟南芥(Arabidopsis thaliana)、梭梭(Haloxylon ammodendron)等植物影响的研究已有相关报道[12-18],这些研究结果均表明,干旱胁迫诱导植物根部合成大量脱落酸后转移到叶片中,进而调节其生长、生理特性、气孔开度及光合作用等,使植物不断适应干旱环境。当前,鲜有针对脱落酸对荒漠优势种植物红砂影响的研究报道。因此,本研究通过分析外施ABA对红砂光合作用和叶绿素荧光参数的影响,拟从光合生理的角度探讨外源ABA对红砂抗旱性的调控机制。
1 材料与方法
1.1 研究区概况
研究区位于甘肃省武威市林业综合服务中心良种繁育基地(103°51′E,38°38′N),地处甘肃省河西走廊东北部。该区属典型的温带大陆性荒漠气候,土壤为碱性沙土,年均降水量为113.2 mm,年均蒸发量为2 604.3 mm,年均气温为7.6 ℃,海拔为1 378 m。
1.2 试验材料与处理
2015年4月17日在甘肃省武威市林业综合服务中心良种繁育基地选择大小、长势较为一致的2年生红砂裸根苗木,株高18 cm,移栽在外口径28 cm、内口径25 cm、高21 cm的塑料花盆中,土壤为沙壤土,每盆1株,置于遮阴棚内缓苗,缓苗期间正常浇水。缓苗结束后,即5月17日进行干旱胁迫,胁迫前将所有苗木浇透水,采用便携式土壤水分速测仪(GMK-770S)和称质量法相结合的方式进行水分控制。计算土壤含水量并根据缺水量于每天17:30—18:00定量补充水分,使土壤含水量为田间持水量的45%~50%。6月2日(控水16 d后)开始人工喷施外源ABA,浓度分别设置为0.1、1、10、100 μmol/L,设3次重复,参照阮英慧等的方法[19]使用手持式喷雾器于晴朗无风的15:00—17:00 在植株叶片正反两面均匀喷施,用量以喷施部位湿润滴水为宜,每周喷施1次ABA,共8次,于ABA处理30、60 d后分别测定光合生理指标[蒸腾速率(Tr)、净光合速率(Pn)、气孔导度(Gs)、胞间CO2浓度(Ci)]并拟合得到光响应曲线,同时测定叶绿素荧光参数等指标。
1.3 测定项目与方法
采用LI-6400光合测定仪,参照周朝彬等的方法[20]测定光合生理指标并拟合得到光响应曲线,测定时控制空气CO2浓度为(375±5) μmol/mol,光合作用有效辐射(PAR)为(2 000±1) μmol/(m2·s),空气温度为(28±0.5) ℃。设置光照度为2 000、1 500、1 000、500、400、300、200、100、0 μmol/(m2·s),每个光照度下的测定时间均为120 s,每个处理选取标记枝条中部的3张成熟叶片进行测定,取平均值。
使用便携式脉冲调制叶绿素荧光仪(PAM-2100,英国)测定外源ABA处理下红砂叶片的叶绿素荧光动力学参数,与光响应曲线的测定同步进行。主要参数有可变荧光(Fv)、最大荧光(Fm)、初始荧光(Fo)、PSⅡ原初光能转化效率(Fv/Fm)、实际光化学反应量子效率ΦPSⅡ[ΦPSⅡ=(Fm′-Fo′)/Fm′]。按照Rohacek的公式[21]计算非光化学淬灭系数qN[qN=(Fm-Fm′)/Fv]、PSⅡ原初光能转化效率[Fv/Fm=(Fm-Fo)/Fm]。其中在测定Fo、Fm前必须使叶片经过30 min的完全暗适应。一般来说,在室内24 h或12 h的暗适应是最好的。由于在野外操作不方便,本试验采用叶绿素荧光分析仪的暗适应夹子遮光处理30 min,每个枝条测定3次,每个处理3次重复。
1.4 统计分析方法
试验数据通过Excel 2010和SPSS 16.0进行处理,试验结果采用Duncans新复极差法进行差异显著性多重比较。
2 结果与分析
2.1 外源ABA对红砂幼苗光合特性的影响
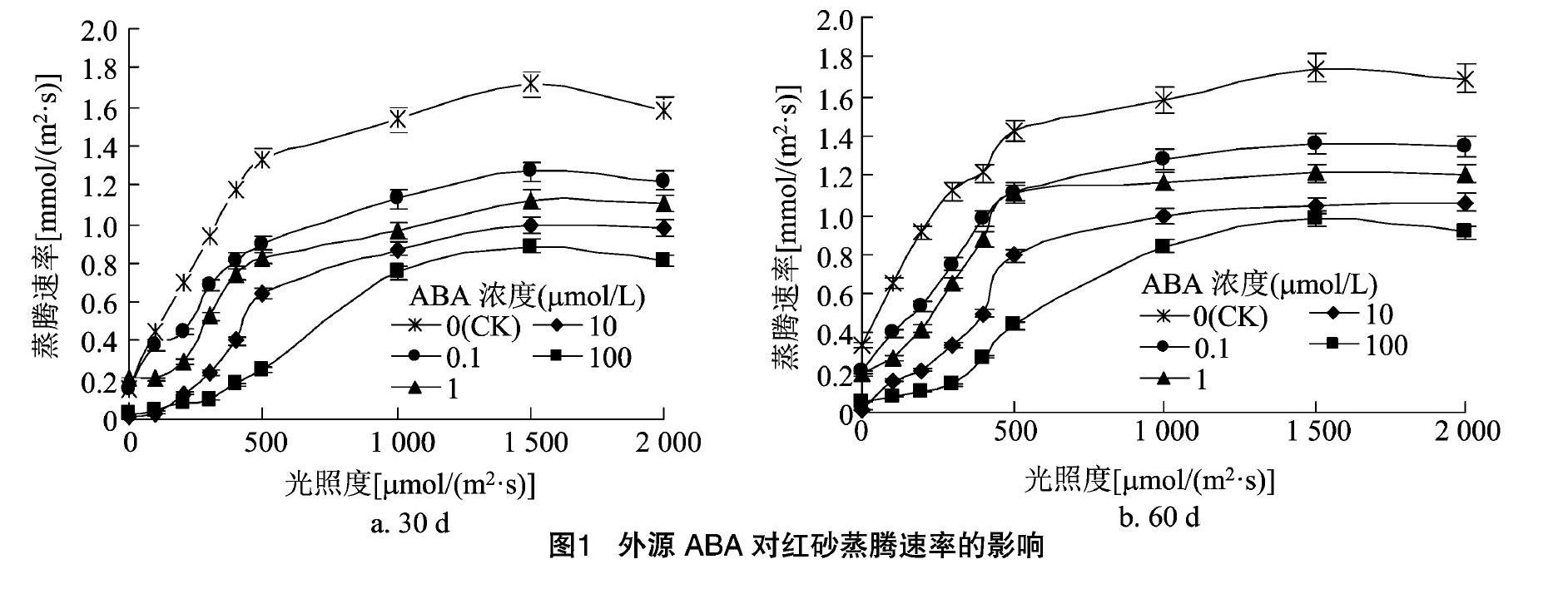
如图1所示,对照红砂叶片的蒸腾速率一直保持在高水平,喷施ABA后,各处理的Tr均不同程度地低于CK。随着光照度的增加,Tr先快速上升然后缓慢上升接着趋于稳定,随着ABA浓度的增大,Tr也随之下降,当ABA浓度为100 μmol/L时,Tr最小且变化趋势最明显,这可能说明外源ABA作用能够诱导红砂叶片气孔关闭,减少蒸腾,导致Tr降低,以便更好地适应外界环境;喷施ABA 60 d时,各处理的Tr均低于胁迫30 d时。由此推测,外施ABA浓度、作用时间均影响蒸腾速率的变化,Tr受限制主要与气孔因素有关。
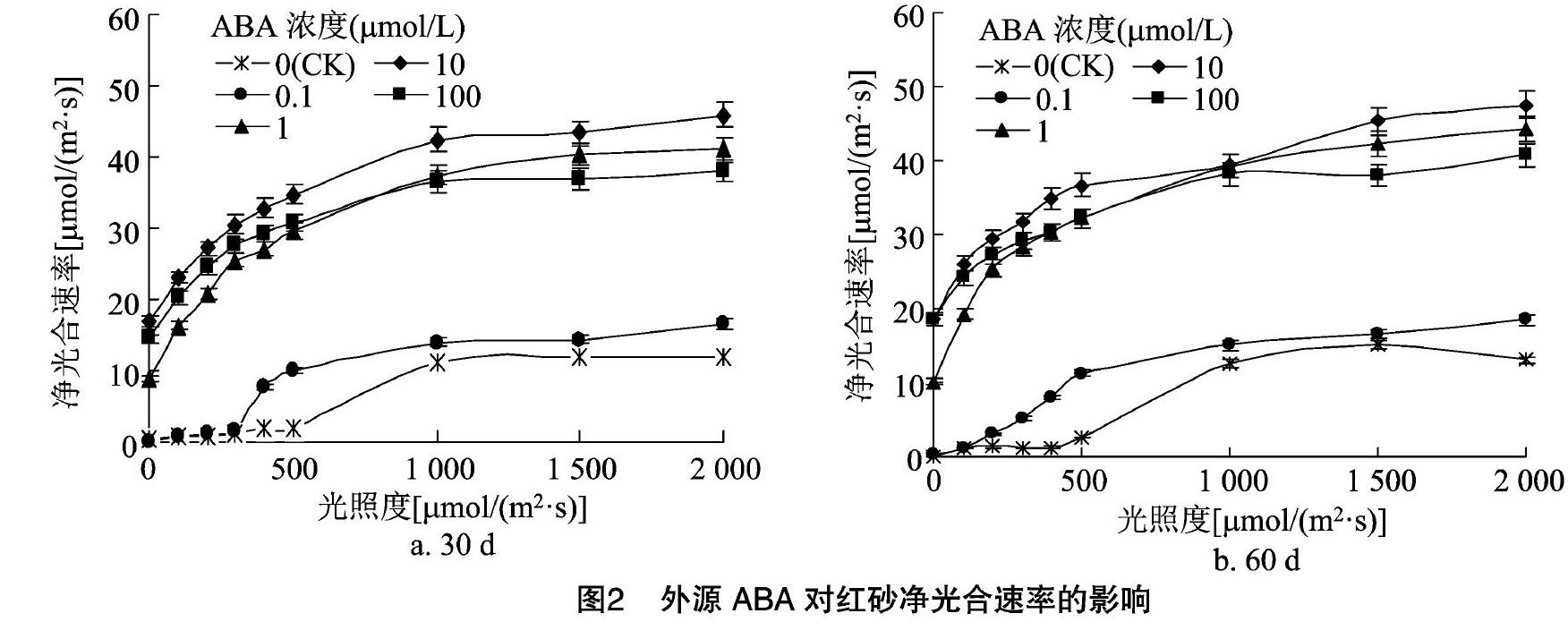
如图2所示,同一ABA浓度下,红砂叶片的净光合速率随着光照度的增大在达到光饱和点(LSP)前表现出先直线式快速升高而后曲线式缓慢升高的变化趋势。对照的Pn维持在最低水平,ABA胁迫30 d后,Pn呈不同程度的上升趋势,在ABA浓度为10 μmol/L时最大,而ABA浓度为100 μmol/L 时,Pn反而下降,出现光饱和点;ABA胁迫60 d后,各处理的Pn均低于胁迫初期且部分处理显著高于对照。说明外施一定浓度的ABA可以有效促進红砂的光合作用,但这种作用有一定的ABA浓度限制(10 μmol/L),达到或超过该浓度,促进作用会减缓。
随着光照度的增加,红砂叶片气孔导度呈先快速上升接着缓慢上升而后下降的趋势(图3)。对照的Gs维持在较高水平,ABA胁迫30 d后,各处理的Gs均随着ABA浓度的增大而显著降低,其中在CK、0.1 μmol/L ABA条件下,Gs的变化幅度较大,ABA浓度为100 μmol/L时,Gs的变化趋势不明显,这可能说明该浓度下红砂的气孔基本上失去了调节作用,导致对光照度变化的响应不敏感;当ABA胁迫60 d后,随着胁迫时间的延长,各处理的Gs均不同程度地高于胁迫初期,说明红砂植株生长在旱季时其Gs低于雨季,ABA胁迫时间的延长可能增强了植物气孔的调节能力,可见该植物在干旱时通过降低气孔开度来减少水分的散失,更有利于红砂适应逆境。
红砂光合作用的胞间CO2浓度的光响应变化规律不同于Gs,有2种不同的趋势:当光照度为0~500 μmol/(m2·s)时,随着光照度升高,在ABA浓度为10、100 μmol/L的条件下,Ci先上升后下降;在CK、0.1 μmol/L ABA和1 μmol/L ABA处理下,Ci呈先下降后上升的趋势(图4)。ABA胁迫30 d后,在0~500 μmol/(m2·s)光照度范围内,各处理的Ci变化幅度较大,但是随着光照度的增加,变化趋势基本一致,均缓慢上升且显著高于CK;ABA胁迫60 d后,随着胁迫时间的延长,各处理的Ci均低于胁迫初期,分析其原因,可能是随着Pn的上升,消耗的CO2增多,胞间CO2不能及时得到补足,所以导致Ci下降,此外,可能与土壤水分条件有关。
2.2 外源ABA对红砂幼苗叶绿素荧光参数的影响
叶绿素荧光是光合作用的探针,通过测定叶绿素荧光参数能够得出相关光能使用路径的信息,间接判断植物生长情况和抗逆性的大小,其中Fv/Fm、Fv/Fo(PSⅡ的潜在光化学活性)、Fm/Fo(最大荧光/初始荧光)、Fv(可变荧光)用来反映质体醌A(QA)的复原情况[22]。光化学效率的高低直接决定叶片光合作用的高低,因此,由于某种原因造成的光化学效率降低会成为光合作用的重要限制因子[23]。在正常供水(CK)条件下,Fv/Fm、Fv/Fo、Fm/Fo、Fv均较大,喷施ABA后,Fv/Fm、Fv/Fo、Fm/Fo、Fv均较CK有不同程度的降低,但是在ABA为100 μmol/L时又高于对照(图5),说明在该浓度下,红砂叶片用于光合电子传递的能量增大,从而维持较高的电子传递速率,促进植物生长。当ABA浓度为0~1 μmol/L 时,Fv/Fm、Fv/Fo、Fm/Fo、Fv均呈下降趋势,在1~10 μmol/L ABA浓度范围内,Fv/Fm、Fv/Fo、Fm/Fo呈缓慢上升趋势,而Fv呈下降趋势,在10~100 μmol/L ABA浓度范围内,各处理的参数均表现出快速直线上升的趋势,说明ABA浓度的增加可有效促进PSⅡ的电子传递活性。当ABA胁迫60 d后,各处理的参数较胁迫30 d时呈下降趋势。以上变化表明,ABA胁迫能够调节红砂对环境的适应性,并且与ABA浓度、胁迫时间有关。
3 讨论与结论
3.1 外源ABA对红砂幼苗光合特性的影响
外施ABA可以调节叶片气孔的开度,使气孔阻力增大,抑制气孔的蒸腾作用,大大降低叶片蒸腾失水的速度[24],因此可以通过提高CO2的同化能力和增强叶黄素循环来提高植物对光抑制的抗性。本试验结果表明,红砂通过保持较高水平的净光合速率和蒸腾速率来生存,属于植物避旱型,叶面喷
[FK(W20][TPZJJ5.tif][FK)]
施ABA后,气孔导度、蒸腾速率和净光合速率都在短时间内得到较好的抑制,抑制作用较为明显;随着光照度增大,Pn在达到光饱和点前表现出先直线式快速上升然后曲线式缓慢升高的规律,随着ABA浓度的增加,红砂叶片Tr和Gs随之降低且变化规律一致,在ABA浓度为100 μmol/L时,Pn下降,Gs随光照度增大的变化趋势不明显,说明其气孔基本上失去了调节作用,导致对光照度变化不敏感,主要与气孔因素有关。
ABA胁迫30 d后,随着光照度的增加,气孔打开,叶片Gs缓慢上升,但快速增加的Pn使得CO2的供应速度赶不上光合速率中CO2消耗的速度,胞间CO2不能及时得到补足,使得Ci在前期迅速下降[光照度≤500 μmol/(m2·s)];随着光照度进一步增加,Pn增加趋势变缓,而此时Gs也进一步开放,因此Ci下降速度变缓,这与周朝彬等对胡杨的研究结果[20]相似。随着ABA胁迫时间的延长,Ci、Pn和Tr均低于胁迫初期;而Gs均高于胁迫初期,这可能说明红砂植株在生长旱季的Gs低于雨季。这些变化表明,适度的ABA浓度(<100 μmol/L)有助于红砂更好地适应逆境,达到或超过此浓度则不利于红砂进行光合作用,10~100 μmol/L的浓度范围较大,有效ABA浓度也有可能存在于这一范围内。另外,以上结果可能还受红砂自身新合成的ABA、红砂植株大小以及叶片选择的影响,更可靠的结论还需通过进一步研究证明。
3.2 外源ABA对红砂幼苗叶绿素荧光参数的影响
当前,越来越多的研究结果表明,叶绿素的荧光信号通过植物体内发出,蕴含的光合作用信息非常丰富,其特征与植物受胁迫的水平有着密切关系,能够迅速、敏锐和无损伤地研究和探测完整植株在应力条件下光合作用的真实活动,常用来评估光合功效以及应激胁迫对叶绿素荧光信号的影响,所以通过植物光合过程中荧光特征的观测,能够了解植物受胁迫的生理状况[25]。许多研究结果表明,逆境胁迫的严重程度与Fm/Fo、Fv/Fo、qP、qN等参数被抑制的水平之间呈正相关关系,这些参數可以作为植物的抗性指数[26],已在多种植物中得到了应用,对玉米和冬小麦幼苗的研究结果表明,随着水分的降低,Fo增大,而Fv、Fv/Fm、Fv/Fo显著降低[27-28]。在通常情况下,暗适应后叶片的Fv/Fm是判断植物是否发生光抑制的指标,未受到环境胁迫时的Fv/Fm>0.8。本试验结果发现,在正常供水(CK)条件下,Fv/Fm、Fv/Fo、Fm/Fo、Fv均维持在较高水平,其中Fv/Fm在0.8以上,喷施ABA后,这些参数均不同程度地降低,但随着ABA浓度变化,各处理的变化趋势一致,即在ABA为0~1 μmol/L的范围内,各参数值均下降;在ABA为1~10 μmol/L的范围内,Fv/Fm、Fv/Fo、Fm/Fo呈缓慢上升趋势,而Fv呈下降趋势;在ABA为10~100 μmol/L的范围内,各处理的参数均表现出直线式快速上升的趋势;当ABA浓度为100 μmol/L时,各参数值均高于对照。由此说明,ABA能缓解干旱胁迫时PSⅡ的伤害,有利于叶片光能的捕捉和转换,从而增强光合作用,缓解胁迫对红砂的伤害程度,促进红砂幼苗生长,这与毛桂莲等对枸杞的研究结果[22]一致。随着ABA胁迫时间的延长,各处理的参数均低于胁迫初期。以上变化说明,红砂幼苗叶绿素荧光参数与ABA胁迫浓度、胁迫时间有关。
综上所述,外施一定浓度ABA(<100 μmol/L),能够缓解干旱胁迫对红砂的伤害程度,有效控制叶片的衰老进程,促进光合作用,超过该浓度时,作用会减小。因此,在荒漠植被恢复中,可通过喷施外源ABA来提高红砂的抗旱性,但必须掌握好喷施的浓度。
参考文献:
[1]胡小文,王彦荣,武艳培. 荒漠草原植物抗旱生理生态学研究进展[J]. 草业学报,2004,13(3):9-15.
[2]马剑英,陈发虎,夏敦胜,等. 荒漠植物红砂(Reaumuria soongorica)叶片元素和水分含量与土壤因子的关系[J]. 生态学报,2008,28(3):983-992.
[3]种培芳,曾继娟,单立山,等. 干旱胁迫下荒漠草地植物红砂幼苗对外源ABA的生理响应[J]. 草地学报,2016,24(5):1001-1008.
[4]刘玉冰,张腾国,安黎哲,等. 红砂正常和脱水组织中总RNA提取的改进CTAB法[J]. 中国沙漠,2006,26(4):600-603.
[5]曾彦军,王彦荣,庄光辉,等. 红砂和霸王种子萌发对干旱与播深条件的响应[J]. 生态学报,2004,24(8):1629-1634.
[6]李秋艳,方海燕. 沙埋对红砂幼苗出土和生长的影响[J]. 水土保持通报,2008,28(1):30-33.
[7]张颖娟,王玉山. 濒危灌木长叶红砂遗传多样性的RAPD分析[J]. 生态学杂志,2008,27(2):157-161.
[8]薛 焱,王迎春,王同智. 濒危植物长叶红砂适应盐胁迫的生理生化机制研究[J]. 西北植物学报,2012,32(1):136-142.
[9]种培芳,苏世平,李 毅. 4个地理种群红砂的抗旱性综合评价[J]. 草业学报,2011,20(5):26-33.
[10]马剑英,方向文,夏敦胜,等. 荒漠植物红砂叶片元素含量与气候因子的關系[J]. 植物生态学报,2008,32(4):848-857.
[11]张淑兰,王化庆. 脱落酸在植物对不良环境响应中的作用[J]. 昌潍师专学报,2000,19(5):23-25,41.
[12]汪月霞,索 标,赵鹏飞,等. 外源ABA对干旱胁迫下不同品种灌浆期小麦psbA基因表达的影响[J]. 作物学报,2011,37(8):1372-1377.
[13]吕祥勇. 干旱胁迫下玉米小苗根系和叶片ABA合成差异研究[D]. 扬州:扬州大学,2007.
[14]王臣立,韩士杰,黄明茹. 干旱胁迫下沙地樟子松脱落酸变化及生理响应[J]. 东北林业大学学报,2001,29(1):40-43.
[15]木合塔尔·扎热. 尖果沙枣幼苗抗旱特性及其对外源脱落酸和外源硅的响应[D]. 乌鲁木齐:新疆农业大学,2009.
[16]陈 娟. 干旱胁迫和外源ABA对生姜生长和根际效应的影响研究[D]. 成都:四川大学,2007.
[17]张静媛,慕自新,张岁岐. 外源ABA对拟南芥根系皮层细胞水力学特征的影响[J]. 植物生理学报,2013,49(3):264-272.
[18]郭新红,姜孝成,潘晓玲. 渗透胁迫和外源脱落酸对梭梭幼苗生理特性的影响[J]. 生命科学研究,2000,4(4):337-342.
[19]阮英慧,董守坤,刘丽君,等. 干旱胁迫下外源脱落酸对大豆花期生理特性的影响[J]. 大豆科学,2012,31(3):385-388,394.
[20]周朝彬,宋于洋,王炳举,等. 干旱胁迫对胡杨光合和叶绿素荧光参数的影响[J]. 西北林学院学报,2009,24(4):5-9.
[21]Rohacek K. Chlorophyll fluorescence parameters:the definitions,photosynthetic meaning,and mutual relationships[J]. Photosynthetica,2002,40(1):13-29.
[22]毛桂莲,许 兴,谢亚军. 外源ABA对NaCl胁迫下枸杞幼苗生长和叶绿素荧光特性的影响[J]. 江苏农业科学,2005(6):111-114.
[23]张秋英,李发东,刘孟雨,等. 不同水分条件下小麦旗叶叶绿素a荧光参数与子粒灌浆速率[J]. 华北农学报,2003,18(1):26-28.
[24]于 洋,王晶英,肖云鹏. 干旱与外源ABA交互作用对水曲柳苗木光合参数的影响[J]. 东北林业大学学报,2009,37(3):41-43,66.
[25]李 晓,冯 伟,曾晓春. 叶绿素荧光分析技术及应用进展[J]. 西北植物学报,2006,26(10):2186-2196.
[26]冯建灿,胡秀丽,毛训甲. 叶绿素荧光动力学在研究植物逆境生理中的应用[J]. 经济林研究,2002,20(4):14-18,30.
[27]王建程,严昌荣,卜玉山. 不同水分与养分水平对玉米叶绿素荧光特性的影响[J]. 中国农业气象,2005,26(2):95-98.
[28]杨晓青,张岁岐,梁宗锁,等. 水分胁迫对不同抗旱类型冬小麦幼苗叶绿素荧光参数的影响[J]. 西北植物学报,2004,24(5):812-816.
